- Обмен липидов. Анаболизм липидов

Содержание
- 2. СИНТЕЗ ЖИРНЫХ КИСЛОТ
- 3. СИНТЕЗ ЖИРНЫХ КИСЛОТ ЛОКАЛИЗАЦИЯ В КЛЕТКЕ: -ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16) -МИТОХОНДРИИ (элонгаза синтезирует
- 4. СИНТЕЗ ЖИРНЫХ КИСЛОТ Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при окислении глюкозы в
- 5. СИНТЕЗ ЖИРНЫХ КИСЛОТ Синтез жирных кислот требует: источник углеродного скелета: ацетил-КоА дополнительный источник углерода: СО2 (для
- 6. Выделяют три этапа образования пальмитиновой кислоты: транспортировка ацетил-КоА из митохондрий в цитоплазму; образование малонил-КоА из ацетил-КоА;
- 7. СИНТЕЗ ЖИРНЫХ КИСЛОТ Транспорт ацетил-КоА в цитоплазму
- 8. СИНТЕЗ ЖИРНЫХ КИСЛОТ Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных кислот Карбоксилирование ацетил-КоА
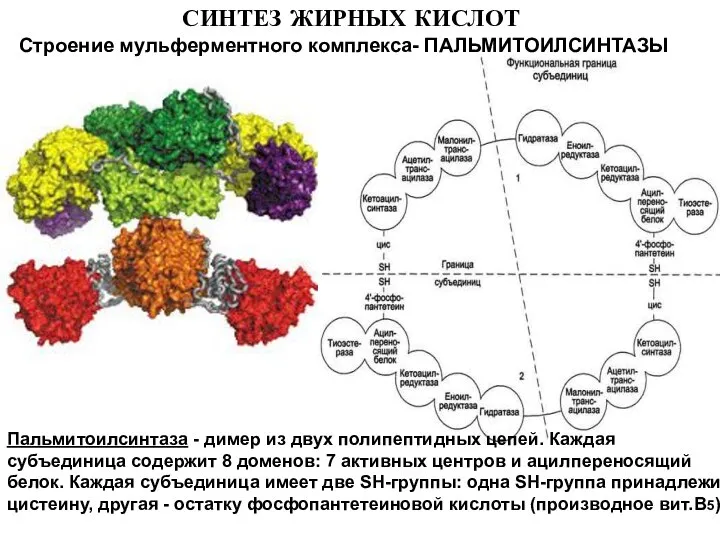
- 9. СИНТЕЗ ЖИРНЫХ КИСЛОТ Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ Пальмитоилсинтаза - димер из двух полипептидных цепей. Каждая субъединица
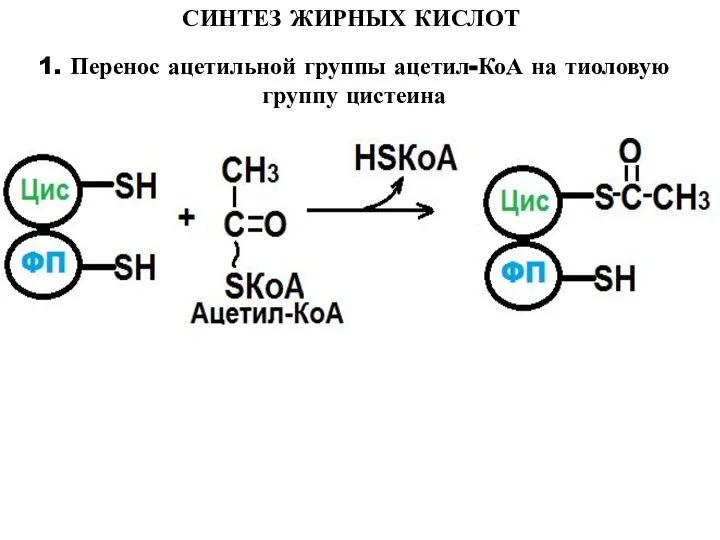
- 10. СИНТЕЗ ЖИРНЫХ КИСЛОТ 1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
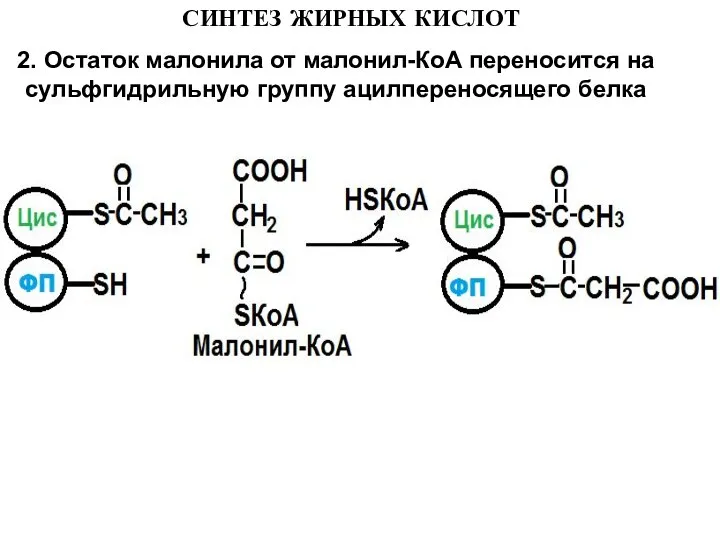
- 11. СИНТЕЗ ЖИРНЫХ КИСЛОТ 2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего белка
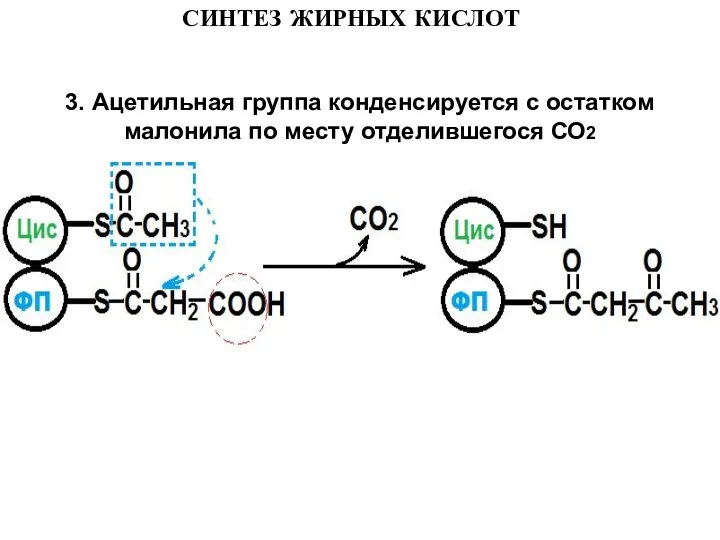
- 12. 3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2 СИНТЕЗ ЖИРНЫХ КИСЛОТ
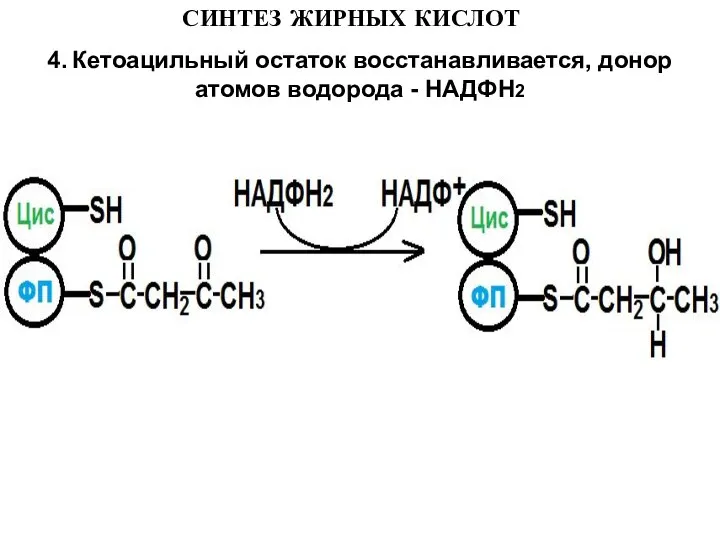
- 13. СИНТЕЗ ЖИРНЫХ КИСЛОТ 4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
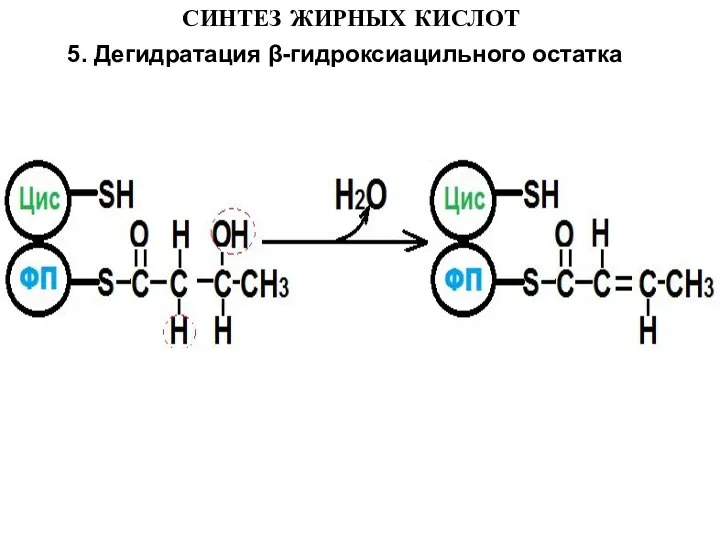
- 14. СИНТЕЗ ЖИРНЫХ КИСЛОТ 5. Дегидратация β-гидроксиацильного остатка
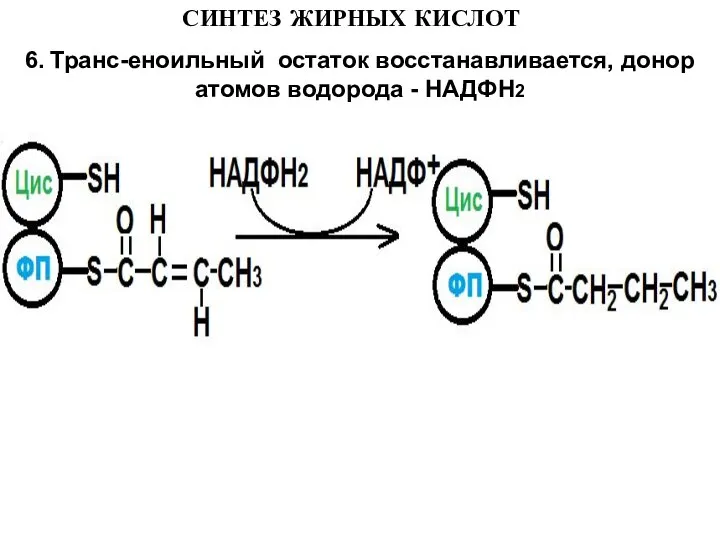
- 15. СИНТЕЗ ЖИРНЫХ КИСЛОТ 6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
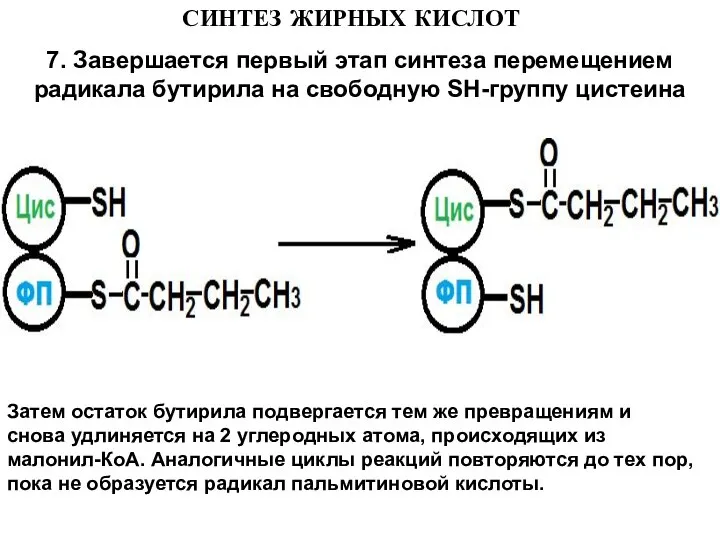
- 16. СИНТЕЗ ЖИРНЫХ КИСЛОТ 7. Завершается первый этап синтеза перемещением радикала бутирила на свободную SH-группу цистеина Затем
- 17. СИНТЕЗ ЖИРНЫХ КИСЛОТ Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту
- 18. ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2- в качестве восстановителя.
- 19. ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ Основные жирные кислоты, образующиеся в организме человека в результате десатурации – пальмитоолеиновая (С16:1Δ9)
- 20. РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ Регулируется пищевым режимом: - тормозит голодание - активирует богатая углеводами пища 2.
- 21. СИНТЕЗ ХОЛЕСТЕРИНА Субстрат для синтеза холестерола- ацетил-КоА Активация синтеза холестерина происходит при поступлении в организм пищи,
- 22. СИНТЕЗ ХОЛЕСТЕРИНА ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА: • синтез мевалоната (С6) • синтез сквалена из мевалоната (С30)
- 23. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА Регуляторная реакция синтеза холестерина
- 24. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
- 25. СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
- 26. РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА 1. Регулируется пищевым режимом: • тормозит голодание • активирует богатая углеводами пища 2.
- 27. СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
- 28. СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ 1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани) 2. Ресинтез триацилглицеролов в
- 29. СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
- 31. Скачать презентацию

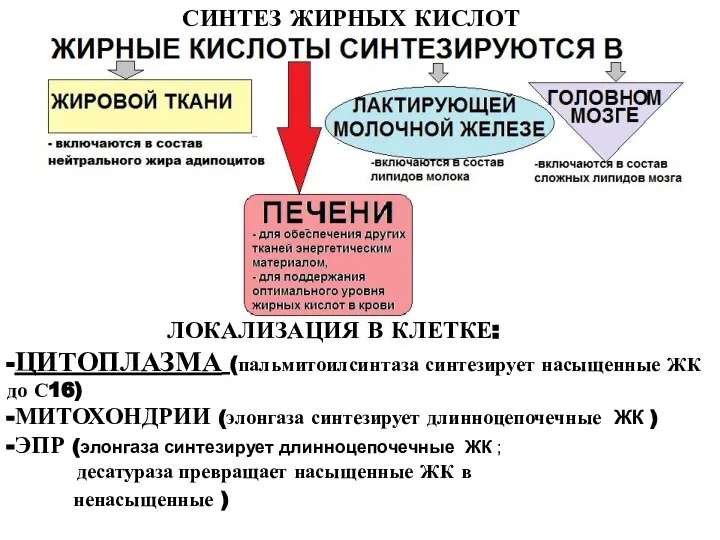
Слайд 3СИНТЕЗ ЖИРНЫХ КИСЛОТ
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16)
-МИТОХОНДРИИ
СИНТЕЗ ЖИРНЫХ КИСЛОТ
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16)
-МИТОХОНДРИИ
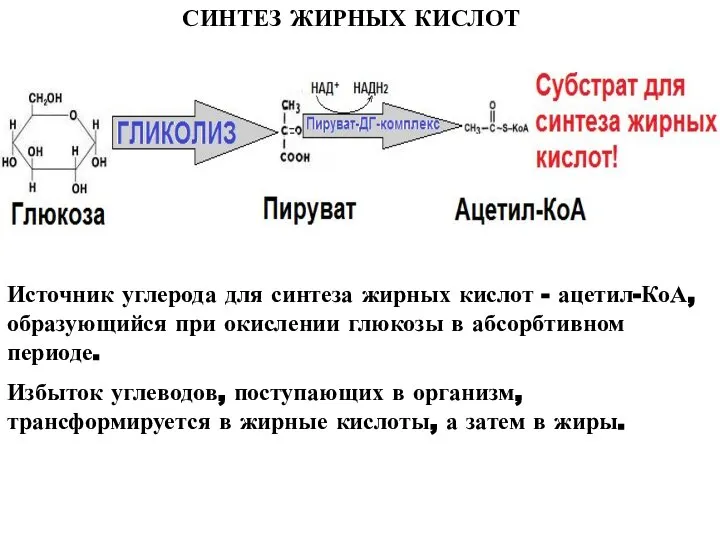
Слайд 4СИНТЕЗ ЖИРНЫХ КИСЛОТ
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
Слайд 5СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
дополнительный источник углерода:
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
дополнительный источник углерода:

Слайд 6 Выделяют три этапа образования пальмитиновой кислоты:
транспортировка ацетил-КоА из митохондрий в цитоплазму;
образование
Выделяют три этапа образования пальмитиновой кислоты:
транспортировка ацетил-КоА из митохондрий в цитоплазму;
образование

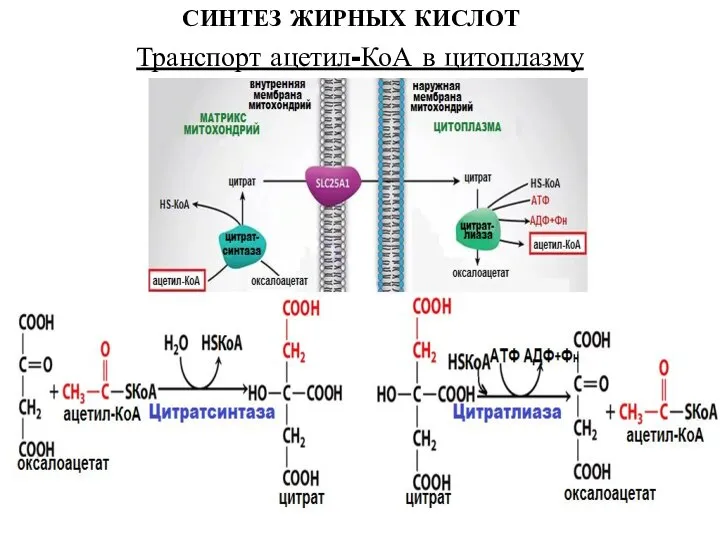
Слайд 7СИНТЕЗ ЖИРНЫХ КИСЛОТ
Транспорт ацетил-КоА в цитоплазму
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Транспорт ацетил-КоА в цитоплазму
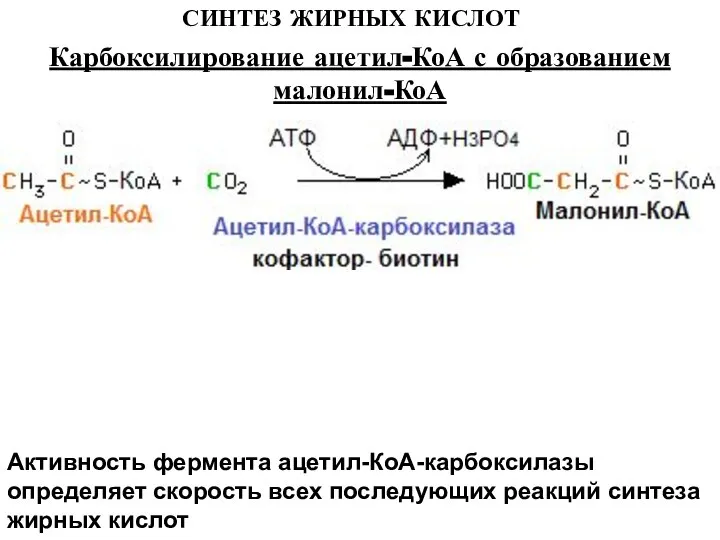
Слайд 8СИНТЕЗ ЖИРНЫХ КИСЛОТ
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
Слайд 9СИНТЕЗ ЖИРНЫХ КИСЛОТ
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
Слайд 10СИНТЕЗ ЖИРНЫХ КИСЛОТ
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
СИНТЕЗ ЖИРНЫХ КИСЛОТ
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
Слайд 11СИНТЕЗ ЖИРНЫХ КИСЛОТ
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
СИНТЕЗ ЖИРНЫХ КИСЛОТ
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
Слайд 123. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
Слайд 13СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
Слайд 14СИНТЕЗ ЖИРНЫХ КИСЛОТ
5. Дегидратация β-гидроксиацильного остатка
СИНТЕЗ ЖИРНЫХ КИСЛОТ
5. Дегидратация β-гидроксиацильного остатка
Слайд 15СИНТЕЗ ЖИРНЫХ КИСЛОТ
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
Слайд 16СИНТЕЗ ЖИРНЫХ КИСЛОТ
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
СИНТЕЗ ЖИРНЫХ КИСЛОТ
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
Слайд 17СИНТЕЗ ЖИРНЫХ КИСЛОТ
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
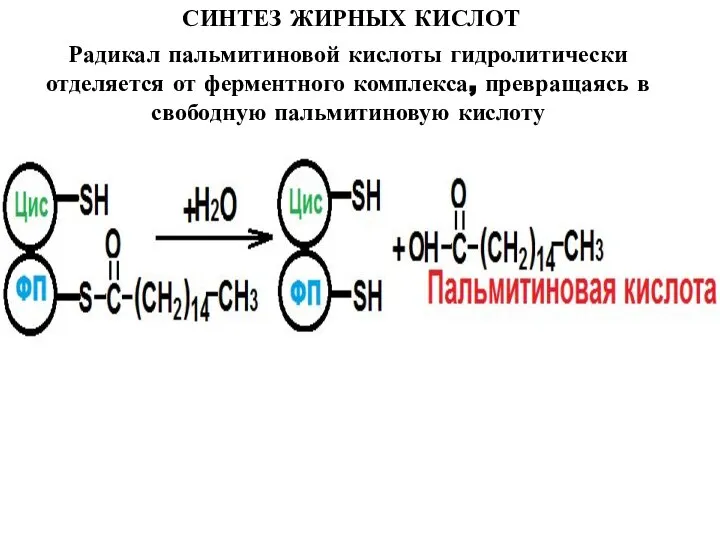
Слайд 18ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-
ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-

Слайд 19ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Основные жирные кислоты, образующиеся в организме человека в результате десатурации
ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Основные жирные кислоты, образующиеся в организме человека в результате десатурации
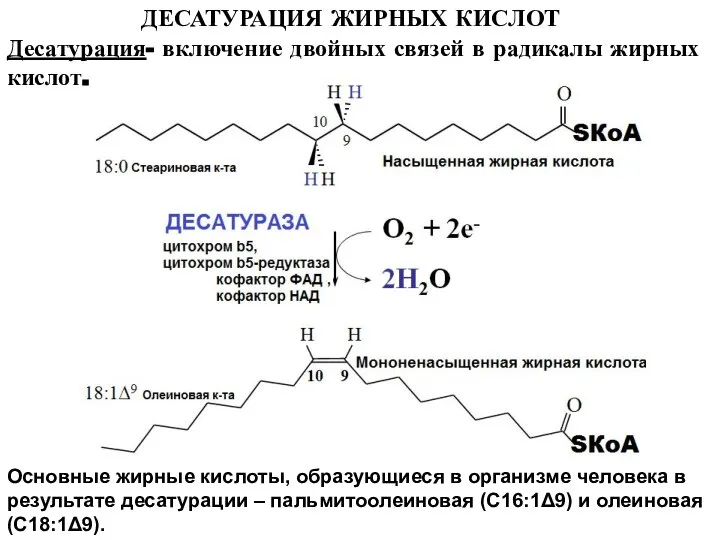
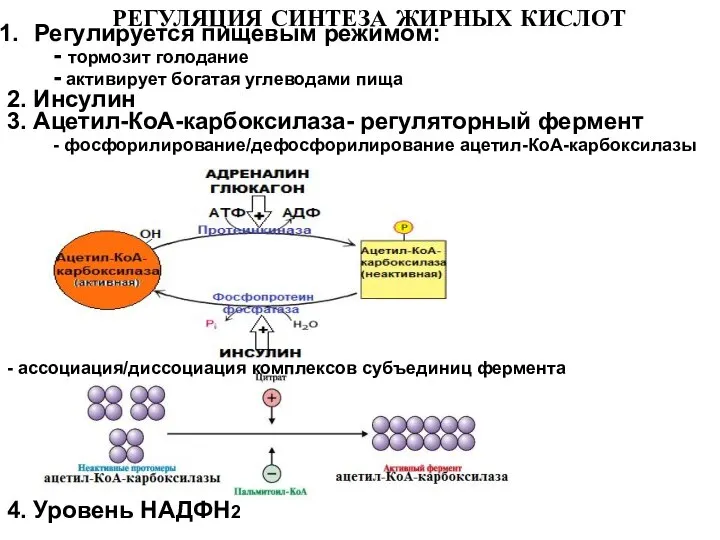
Слайд 20РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
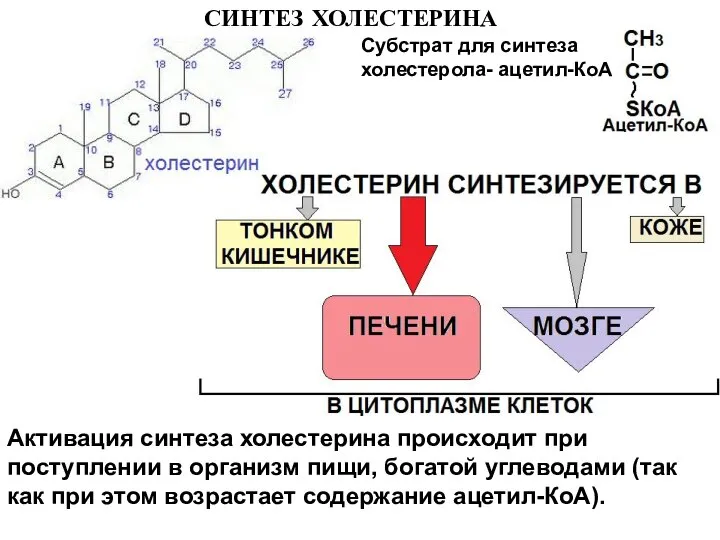
Слайд 21СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
Слайд 22СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната
СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната

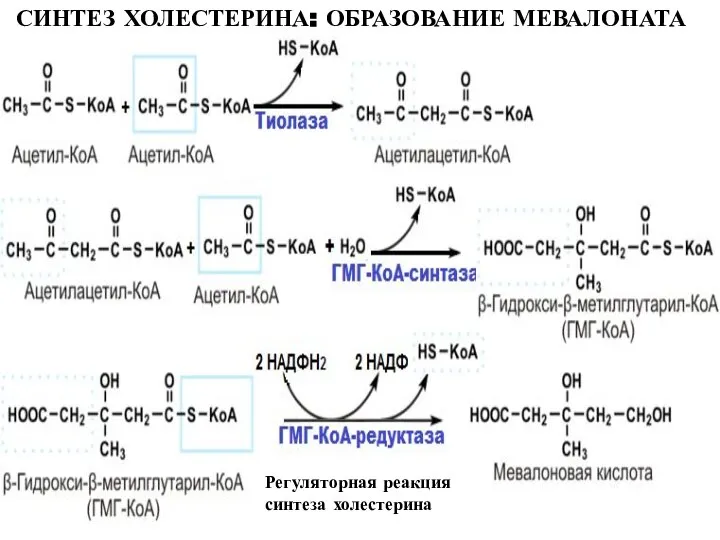
Слайд 23СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
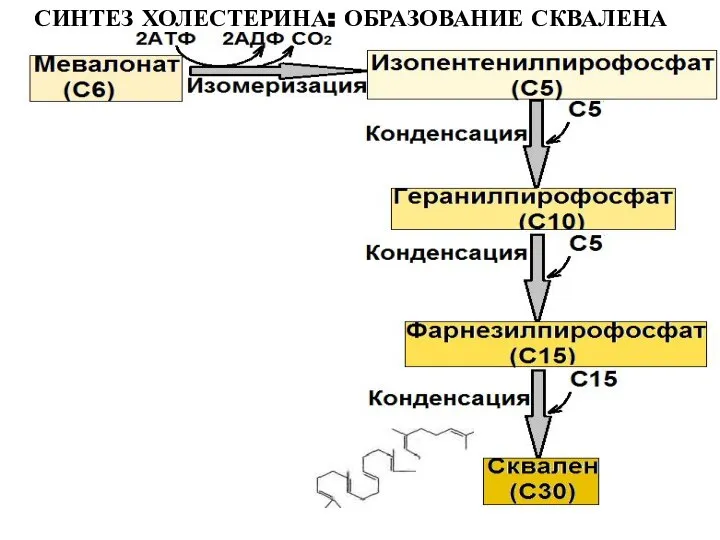
Слайд 24СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
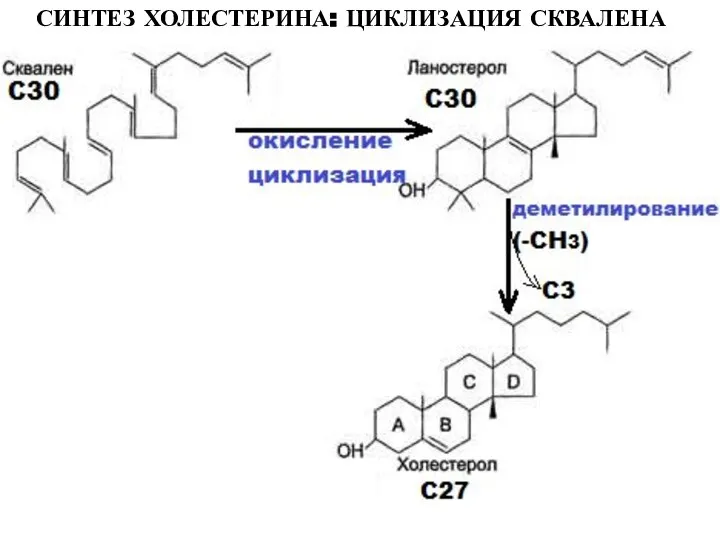
Слайд 25СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
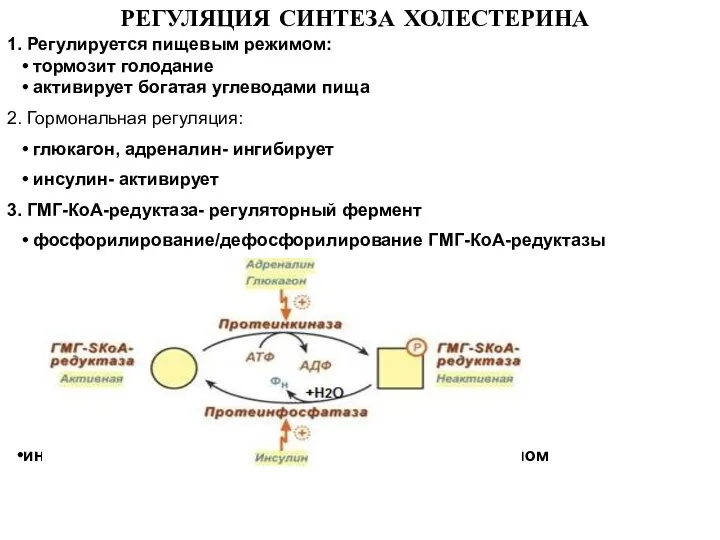
Слайд 26 РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
Слайд 27СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
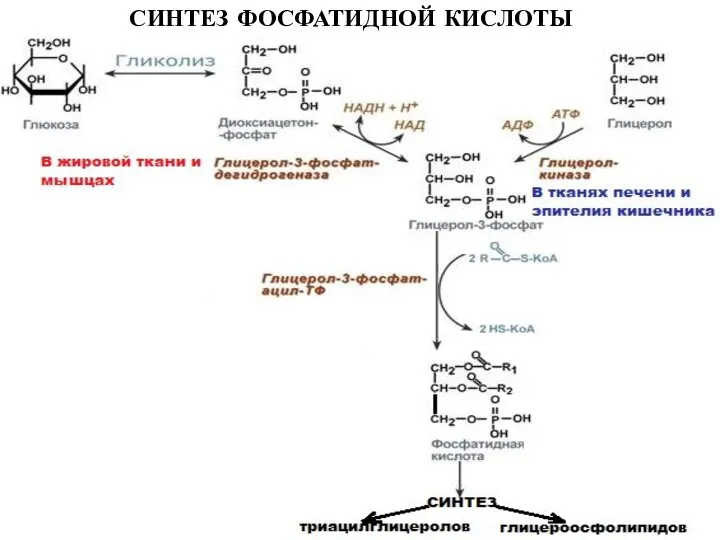
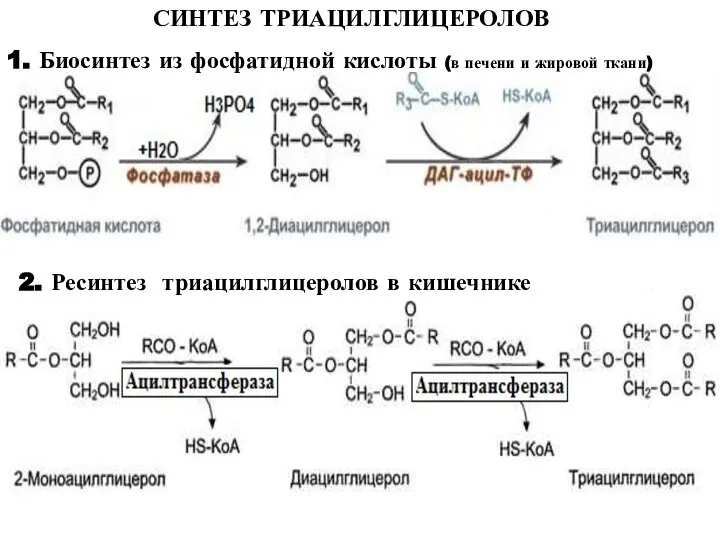
Слайд 28СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
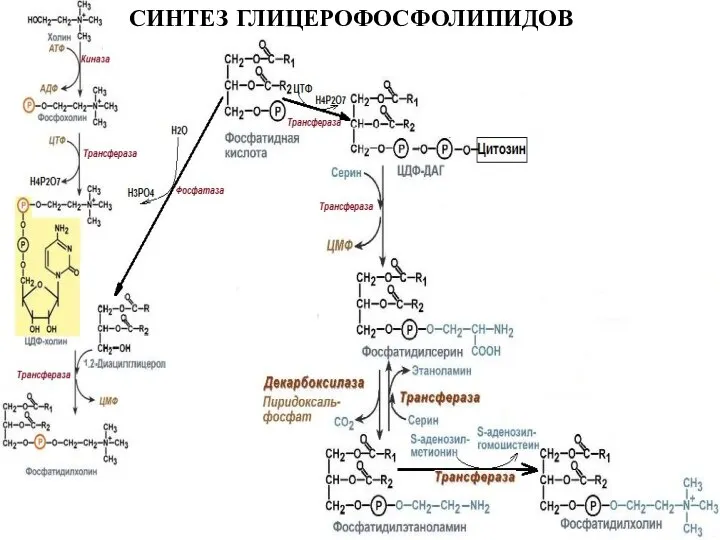
Слайд 29СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
 Карнотавр - род хищных динозавров из семейства абелизаврид
Карнотавр - род хищных динозавров из семейства абелизаврид Отряды птиц. Воробьинообразные и голенастые
Отряды птиц. Воробьинообразные и голенастые Зоология. Введение
Зоология. Введение Решение задач по биохимии. 10 класс
Решение задач по биохимии. 10 класс Воздушное питание. Фотосинтез
Воздушное питание. Фотосинтез Обыкновенная корневая гниль
Обыкновенная корневая гниль Вермикомпостирование жиросодержащих отходов и избыточного ила с целью получения биоудобрения
Вермикомпостирование жиросодержащих отходов и избыточного ила с целью получения биоудобрения Аэробное и анаэробное дыхание
Аэробное и анаэробное дыхание Ирис
Ирис Класс Пресмыкающиеся
Класс Пресмыкающиеся Клетка№1
Клетка№1 Презентация на тему Звери запасаются жиром
Презентация на тему Звери запасаются жиром  Презентация на тему Кроссворд на тему Организм человека
Презентация на тему Кроссворд на тему Организм человека  Шляпочные грибы
Шляпочные грибы Лактоза и её компания
Лактоза и её компания Многообразие водорослей
Многообразие водорослей Генетика популяций
Генетика популяций Транспорт веществ на примере окрашивания розы
Транспорт веществ на примере окрашивания розы Психофизиология. Психофизиологическая проблема
Психофизиология. Психофизиологическая проблема Вирусы. Клеточные и неклеточные формы. Часть 3
Вирусы. Клеточные и неклеточные формы. Часть 3 Почва и её подземные обитатели
Почва и её подземные обитатели Шырайгүл, петуния
Шырайгүл, петуния Осень в жизни травоядных и всеядных
Осень в жизни травоядных и всеядных Пингвины
Пингвины Понятие о межклеточной кооперации в иммуногенезе
Понятие о межклеточной кооперации в иммуногенезе Общая характеристика типа Кольчатые черви на примере дождевого червя
Общая характеристика типа Кольчатые черви на примере дождевого червя Макростехиометрические характеристики биотехнологического процесса
Макростехиометрические характеристики биотехнологического процесса Явления и закономерности на клеточном уровне. Многообразие клеток
Явления и закономерности на клеточном уровне. Многообразие клеток