- Биосинтез и катаболизм коллагена, гликозаминогликанов и протеогликанов

Содержание
- 2. Cодержание Синтез и катаболизм коллагена. Заболевания, связанные с нарушением синтеза и созревания коллагена Синтез и катаболизм
- 3. ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА Коллаген синтезируют и поставляют в межклеточный матрикс почти все клетки (фибробласты,
- 4. ВНУТРИКЛЕТОЧНЫЕ СТАДИИ СИНТЕЗА КОЛЛАГЕНА (ЭТАП I) Протеолиз N- пептидов а). b). c). d). e). f).
- 5. Внутриклеточный этап синтеза коллагена Синтез α-цепей коллагена на полирибосомах – пре-про- коллагена. 2. Сигнальный пептид ориентирует
- 6. Рис. 1.2. Структурная формула аскорбиновой кислоты. Гидроксилирование остатков пролина и лизина, которое начинается в период трансляции
- 7. Рис. 1.3. Гидроксилирование остатков пролина в проколлагеновой α-цепи с образованием 4-гидроксипролина. Реакция гидроксилирования: один атом кислорода
- 9. Следующая посттрансляционная модификация радикалов аминокислот про-α-цепи коллагена — гликозилирование остатков гидроксилизина. К ОН — группам радикала
- 10. По окончании гидроксилирования и гликозилирования три про-α-цепи коллагена соединяются между собой водородными связями, в С- концевых
- 11. Внеклеточные стадии синтеза коллагена (этап II) g). h). i). Микрофибриллы Образование макрофибрилл и волокон коллагена j).
- 12. Первой модификацией внеклеточного этапа является частичный протеолиз. Под действием аминопептидазы и карбоксипептидазы отщепляются N- и С-концевые
- 13. Внеклеточный этап - модификация молекул проколлагена Рис. 1.4. Окисление лизина в структуре коллагена: 1 - формирование
- 14. На следующем этапе аллизин и 5-гидроксиаллизин конденсируются вместе с лизиловыми и гидроксилизиловыми остатками; формируются внутри- и
- 15. Ассоциация молекул коллагена по принципу «бок-в-бок». Ассоциация молекул («сшивка») нерастворимого коллагена по принципу «бок-в-бок». Сборка фибрилл
- 16. Ферменты: 1 - проколлагенпролин-4-диоксигеназа; 2 - проколлагенлизин-5-диоксигеназа; 3 - протеин-лизин-6-оксидаза. Рис. 1.1. Синтез коллагена. Схема синтеза
- 17. Прочность соединения молекул тропоколлагена в микрофибриллах обеспечивается поперечными ковалентными сшивками, которые образуются между радикалами аминокислот —
- 18. Все типы коллагенов в зависимости от структуры делят на несколько групп: фибриллообразующие, ассоциированные с фибриллами коллагена,
- 19. Типы коллагеновых белков в тканях полости рта Таблица 1.1 Локализация основных типов коллагеновых белков в тканях
- 20. Регуляция обмена коллагена Синтез коллагена регулируется разными способами. Сам коллаген и N-npo-пептиды после своего отщепления тормозят
- 24. Нарушение синтеза коллагеновых белков у человека Любые нарушения в синтезе коллагеновых белков клинически проявляются, в первую
- 25. Структура коллагена при сахарном диабете При сахарном диабете вследствие неспособности клеток захватывать глюкозу из плазмы крови
- 26. Несовершенный остеогенез— это генетическое нарушение, которое вызывает в организме недостаточное или неправильное образование фиброзной ткани, основным
- 27. МЕТАБОЛИЗМ ЭЛАСТИНА Синтез эластина: Синтез и созревание белка эластина состоит из двух этапов: внутриклеточный и внеклеточный.
- 28. I. Внутриклеточный этап. Мономерная форма эластина синтезируется на рибосомах, связанных с ЭПР, и называется тропоэластином. В
- 29. Рис. Поперечные сшивки в структуре эластина: А - десмозин, образованый четырьмя остатками лизина; Б - лизиннорлейцин,
- 30. Аминокислотный состав эластина: - 27% глицина; - 19% аланина; - 10% валина; - 4,7% лейцина. Благодаря
- 31. СТРУКТУРА ЭЛАСТИНА Рис. 1.14. Структурная модель эластина: А - состояние расслабления; Б - состояние растяжения. Катаболизм
- 32. НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА При снижении образования десмозинов (или их отсутствии) поперечные сшивки образуются в недостаточном количестве
- 33. Состав и строение муцинов (от анг. mucus – слизь) Гликопротеины, в которых много остатков пролина (до
- 34. Синтез муцинов Высокоспециализированные клетки поднижнечелюстных и подъязычных слюнных желез синтезируют белок муцин. В этих клетках хорошо
- 35. Присоединения GalNAc к остаткам серина и треонина обеспечивается GalNAс-трансферазой , происходит в цис-цистернах аппарата Гольджи. После
- 36. В слюне присутствует муцин-1 (Mr 250 кДа) и муцин-2 (Mr 100 кДа). Внутренние поверхности ротовой полости,
- 37. Муцины выполняют специфические функции: Основные белки, обеспечивающие вязкость слюны. Муцины участвуют в образовании мицелл слюны, (помогают
- 38. Протеогликаны и гликозаминогликаны Протеогликаны - состоят из различных стержневых (коровых) белков, к которым через N- и
- 39. Особенности строения важнейших гликозаминогликанов
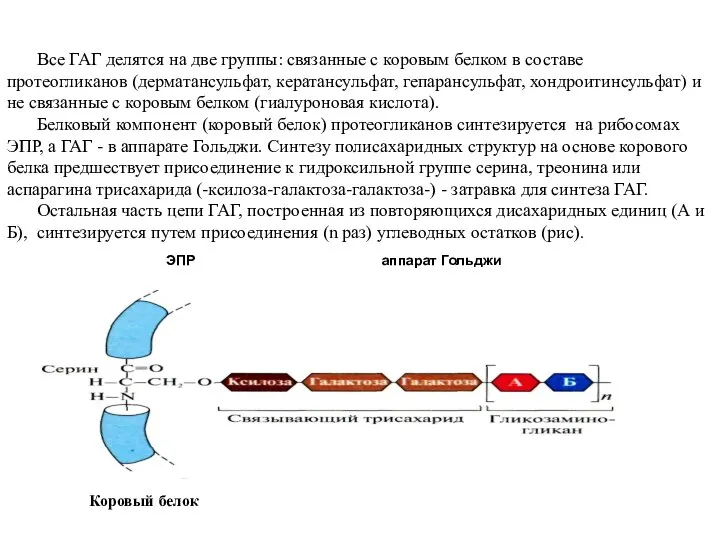
- 40. Все ГАГ делятся на две группы: связанные с коровым белком в составе протеогликанов (дерматансульфат, кератансульфат, гепарансульфат,
- 41. Структура надмолекулярного протеогликанового комплекса
- 44. Аминосахара и гексуроновые кислоты синтезируются из глюкозы. Непосредственным же предшественником N-ацетилглюкозамина и N-ацетилгалактозамина является фруктозо-6-фосфат. Источником
- 45. Рис. 1.21. Синтез гликозаминогликанов. Ферменты: 1 - гексокиназа; 2 - фосфоглюкоизомераза; 3 - аминотрансфераза; 4 -
- 46. Разные протеогликаны отличаются размерами молекул, относительным содержанием белка и набором гликозаминогликанов. Протеогликаны в больших количествах представлены:
- 47. Распад протеогликанов Распад протеогликанов - физиологический процесс, заключающийся в регулярном обновлении внеклеточных и внутриклеточных макромолекул. В
- 48. 2. Распад гликозаминогликанов Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2 ) многих из них
- 49. Протеогликаны и гликозаминогликаны в тканях полости рта
- 50. ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ они являются структурными компонентами межклеточного матрикса; протеогликаны и гликозаминогликаны специфически взаимодействуют с
- 51. Мукополисахаридозы - тяжёлые наследственные заболевания, обусловленые дефектами гидролаз, участвующих в катаболизме гликозаминогликанов. В лизосомах тканей, для
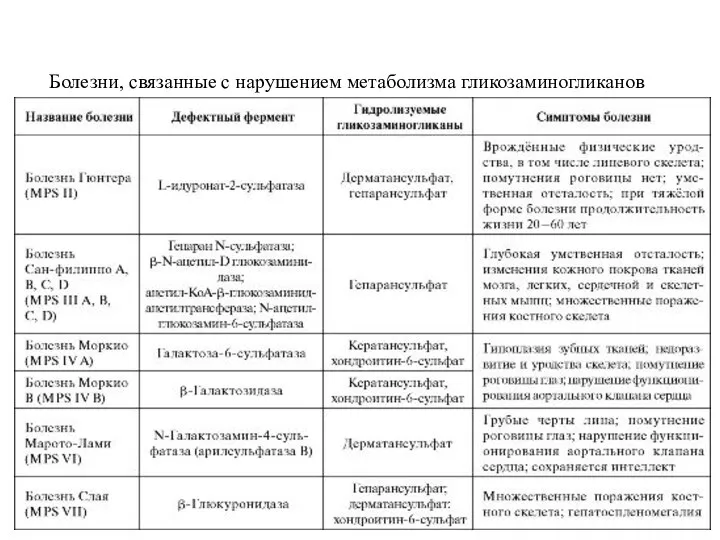
- 52. Болезни, связанные с нарушением метаболизма гликозаминогликанов
- 54. Скачать презентацию
Слайд 2Cодержание
Синтез и катаболизм коллагена.
Заболевания, связанные с нарушением синтеза и созревания коллагена
Синтез
Cодержание
Синтез и катаболизм коллагена.
Заболевания, связанные с нарушением синтеза и созревания коллагена
Синтез

Слайд 3ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Коллаген синтезируют и поставляют в межклеточный матрикс почти
ЭТАПЫ СИНТЕЗА И СОЗРЕВАНИЯ КОЛЛАГЕНА
Коллаген синтезируют и поставляют в межклеточный матрикс почти

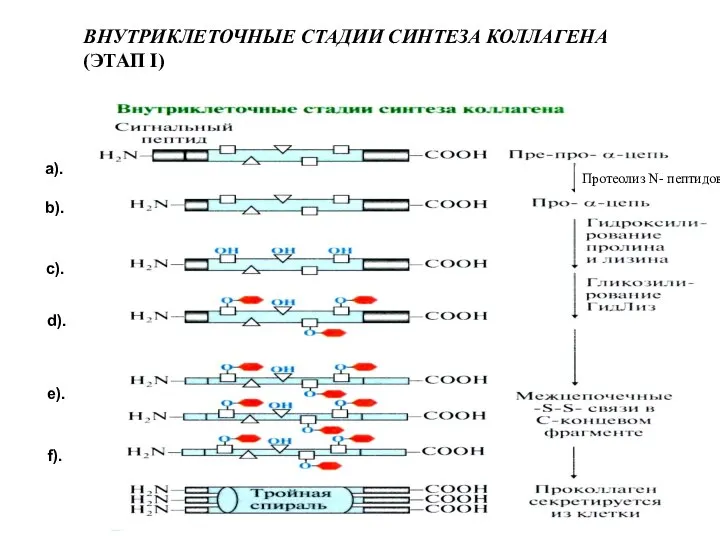
Слайд 4ВНУТРИКЛЕТОЧНЫЕ СТАДИИ СИНТЕЗА КОЛЛАГЕНА (ЭТАП I)
Протеолиз N- пептидов
а).
b).
c).
d).
e).
f).
ВНУТРИКЛЕТОЧНЫЕ СТАДИИ СИНТЕЗА КОЛЛАГЕНА (ЭТАП I)
Протеолиз N- пептидов
а).
b).
c).
d).
e).
f).
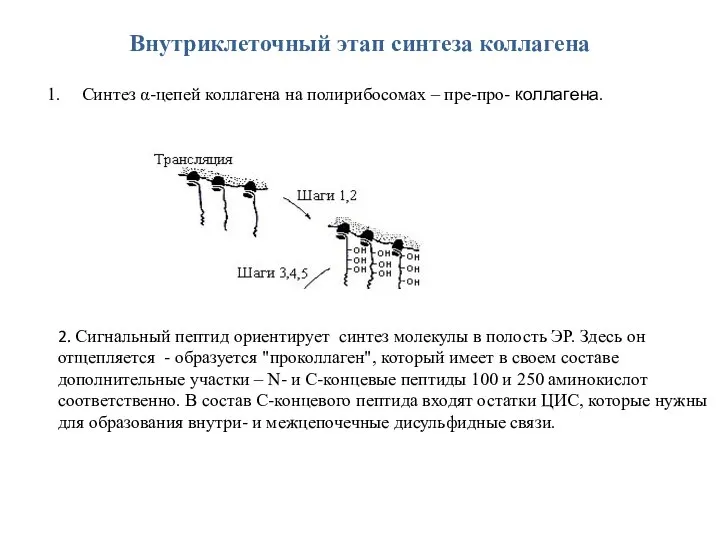
Слайд 5Внутриклеточный этап синтеза коллагена
Синтез α-цепей коллагена на полирибосомах – пре-про- коллагена.
2. Сигнальный
Внутриклеточный этап синтеза коллагена
Синтез α-цепей коллагена на полирибосомах – пре-про- коллагена.
2. Сигнальный
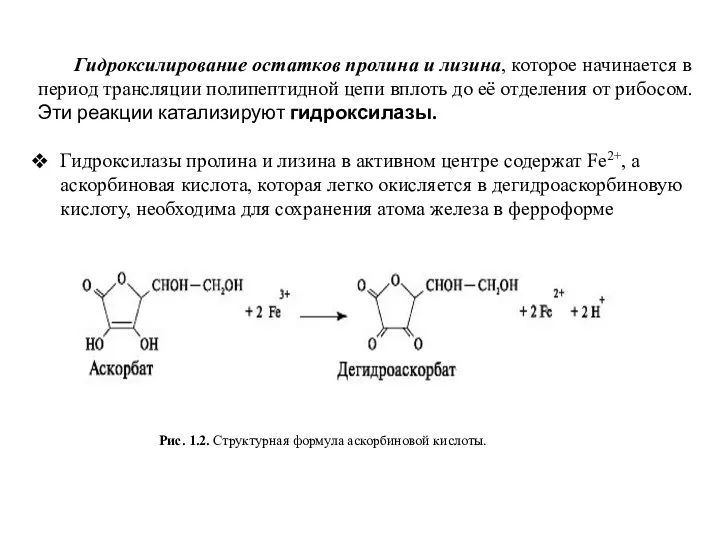
Слайд 6Рис. 1.2. Структурная формула аскорбиновой кислоты.
Гидроксилирование остатков пролина и лизина, которое начинается в
Рис. 1.2. Структурная формула аскорбиновой кислоты.
Гидроксилирование остатков пролина и лизина, которое начинается в
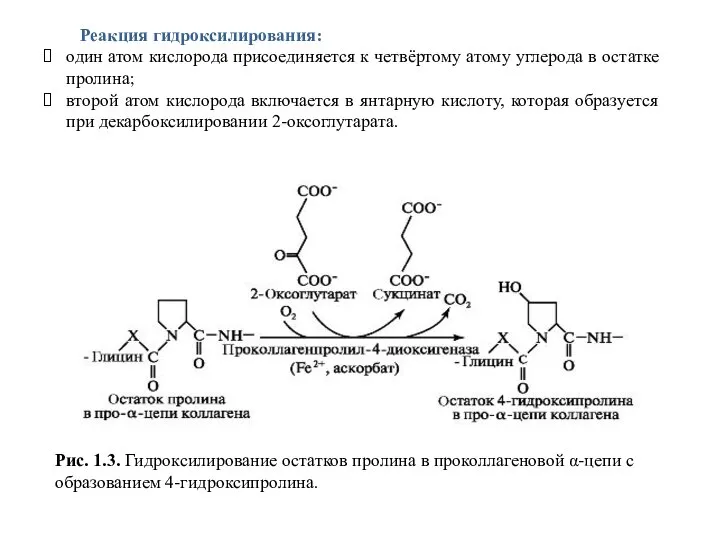
Слайд 7Рис. 1.3. Гидроксилирование остатков пролина в проколлагеновой α-цепи с образованием 4-гидроксипролина.
Реакция гидроксилирования:
один
Рис. 1.3. Гидроксилирование остатков пролина в проколлагеновой α-цепи с образованием 4-гидроксипролина.
Реакция гидроксилирования:
один

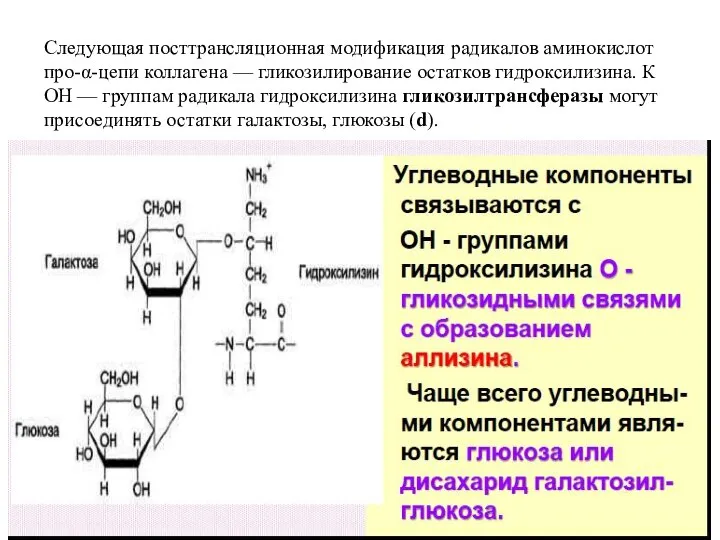
Слайд 9Следующая посттрансляционная модификация радикалов аминокислот про-α-цепи коллагена — гликозилирование остатков гидроксилизина. К
Следующая посттрансляционная модификация радикалов аминокислот про-α-цепи коллагена — гликозилирование остатков гидроксилизина. К
Слайд 10По окончании гидроксилирования и гликозилирования три про-α-цепи коллагена соединяются между собой водородными
По окончании гидроксилирования и гликозилирования три про-α-цепи коллагена соединяются между собой водородными

Слайд 11Внеклеточные стадии синтеза коллагена (этап II)
g).
h).
i).
Микрофибриллы
Образование макрофибрилл и волокон коллагена
j).
Этапы формирования коллагенового
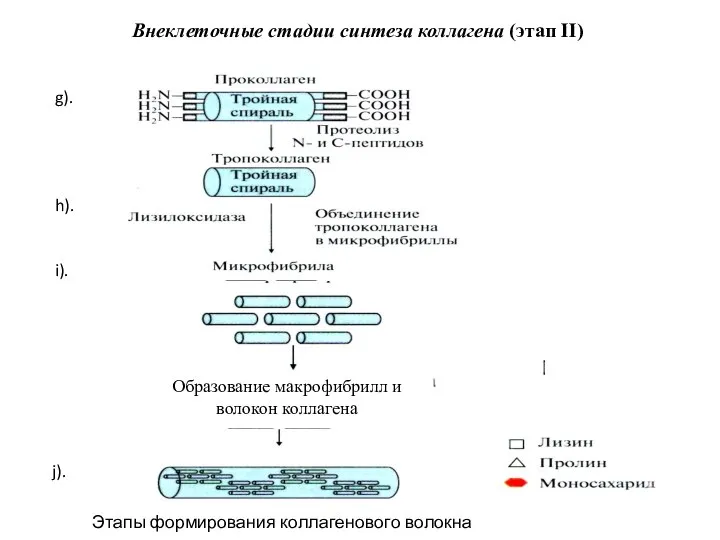
Внеклеточные стадии синтеза коллагена (этап II)
g).
h).
i).
Микрофибриллы
Образование макрофибрилл и волокон коллагена
j).
Этапы формирования коллагенового
Слайд 12Первой модификацией внеклеточного этапа является частичный протеолиз. Под действием аминопептидазы и карбоксипептидазы
Первой модификацией внеклеточного этапа является частичный протеолиз. Под действием аминопептидазы и карбоксипептидазы

Слайд 13Внеклеточный этап - модификация молекул проколлагена
Рис. 1.4. Окисление лизина в структуре коллагена:
1 - формирование
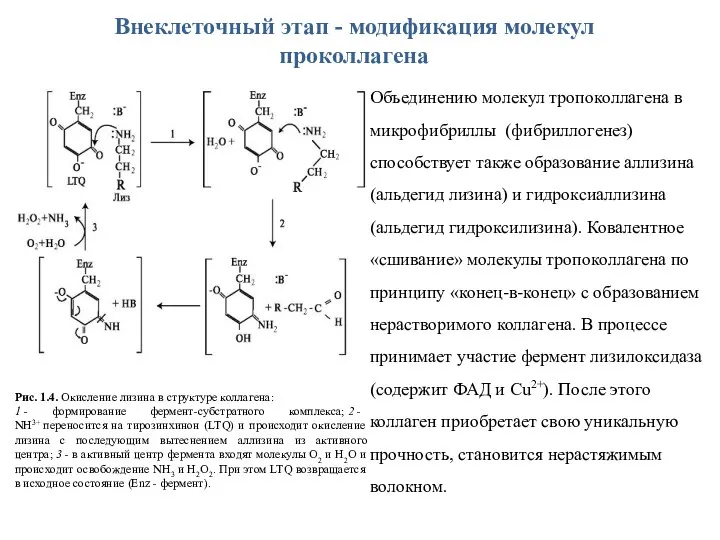
Внеклеточный этап - модификация молекул проколлагена
Рис. 1.4. Окисление лизина в структуре коллагена:
1 - формирование
Слайд 14 На следующем этапе аллизин и 5-гидроксиаллизин конденсируются вместе с лизиловыми и гидроксилизиловыми
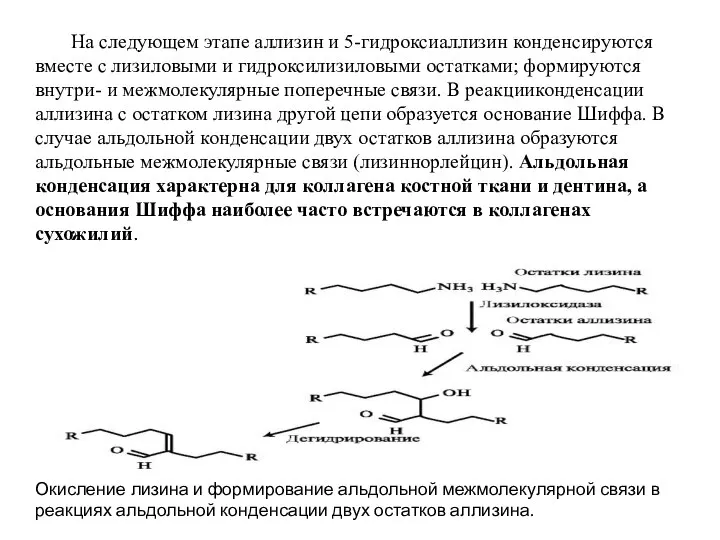
На следующем этапе аллизин и 5-гидроксиаллизин конденсируются вместе с лизиловыми и гидроксилизиловыми
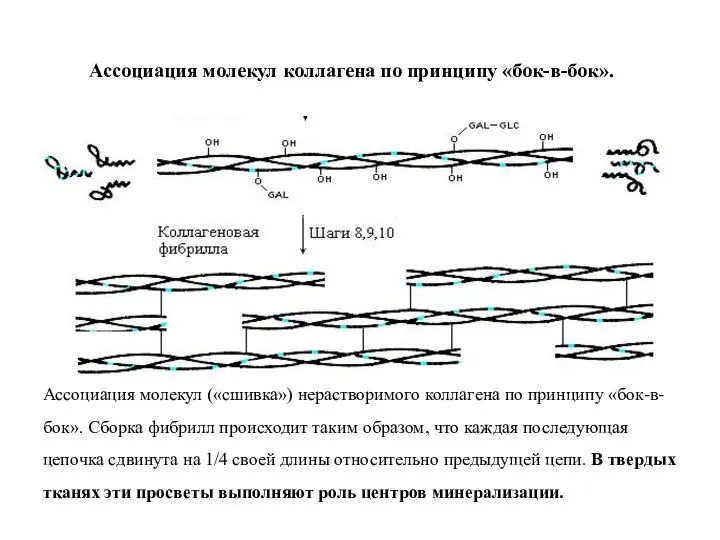
Слайд 15Ассоциация молекул коллагена по принципу «бок-в-бок».
Ассоциация молекул («сшивка») нерастворимого коллагена по
Ассоциация молекул коллагена по принципу «бок-в-бок».
Ассоциация молекул («сшивка») нерастворимого коллагена по
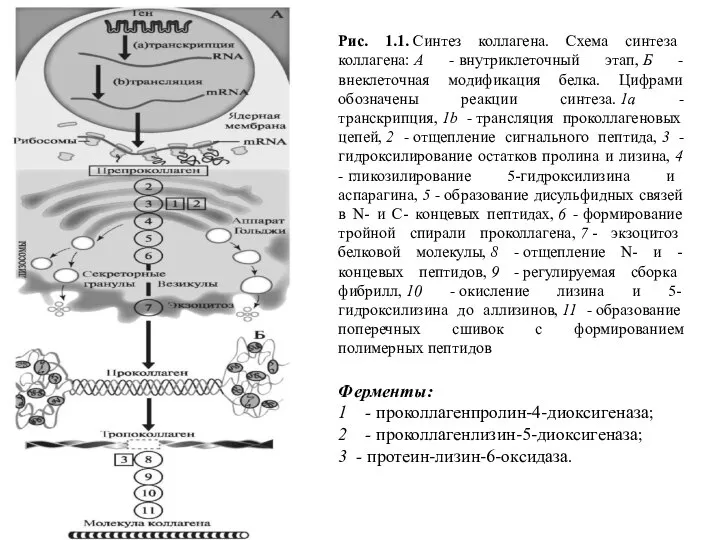
Слайд 16Ферменты:
1 - проколлагенпролин-4-диоксигеназа;
2 - проколлагенлизин-5-диоксигеназа;
3 - протеин-лизин-6-оксидаза.
Рис. 1.1. Синтез коллагена. Схема синтеза коллагена: А - внутриклеточный этап, Б -внеклеточная
Ферменты:
1 - проколлагенпролин-4-диоксигеназа;
2 - проколлагенлизин-5-диоксигеназа;
3 - протеин-лизин-6-оксидаза.
Рис. 1.1. Синтез коллагена. Схема синтеза коллагена: А - внутриклеточный этап, Б -внеклеточная
Слайд 17Прочность соединения молекул тропоколлагена в микрофибриллах обеспечивается поперечными ковалентными сшивками, которые образуются
Прочность соединения молекул тропоколлагена в микрофибриллах обеспечивается поперечными ковалентными сшивками, которые образуются

Слайд 18
Все типы коллагенов в зависимости от структуры делят на несколько групп:
фибриллообразующие,
ассоциированные
Все типы коллагенов в зависимости от структуры делят на несколько групп:
фибриллообразующие,
ассоциированные

Слайд 19Типы коллагеновых белков в тканях полости рта
Таблица 1.1
Локализация основных типов коллагеновых белков
Типы коллагеновых белков в тканях полости рта
Таблица 1.1
Локализация основных типов коллагеновых белков

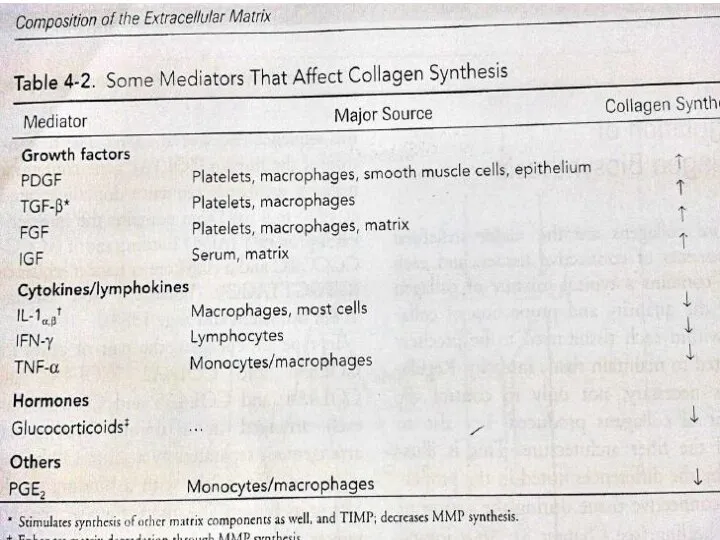
Слайд 20Регуляция обмена коллагена
Синтез коллагена регулируется разными способами.
Сам коллаген и N-npo-пептиды после
Регуляция обмена коллагена
Синтез коллагена регулируется разными способами.
Сам коллаген и N-npo-пептиды после



Слайд 24
Нарушение синтеза коллагеновых белков у человека
Любые нарушения в синтезе коллагеновых белков клинически
Нарушение синтеза коллагеновых белков у человека
Любые нарушения в синтезе коллагеновых белков клинически

Слайд 25Структура коллагена при сахарном диабете
При сахарном диабете вследствие неспособности клеток захватывать глюкозу
Структура коллагена при сахарном диабете
При сахарном диабете вследствие неспособности клеток захватывать глюкозу

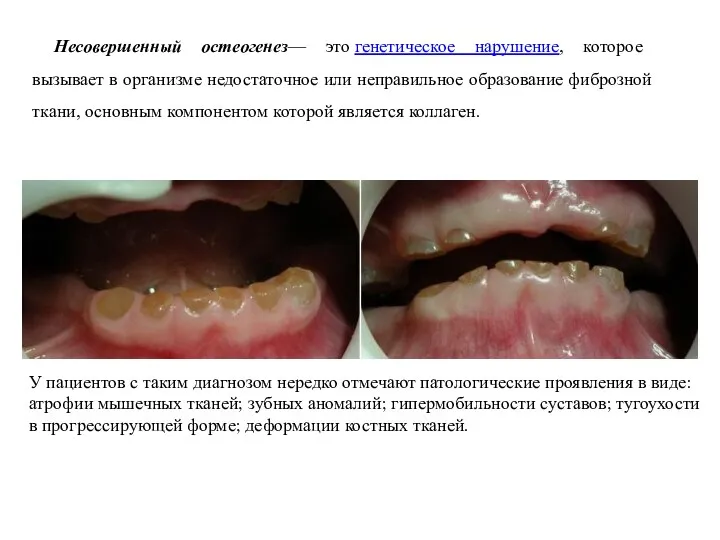
Слайд 26Несовершенный остеогенез— это генетическое нарушение, которое вызывает в организме недостаточное или неправильное образование
Несовершенный остеогенез— это генетическое нарушение, которое вызывает в организме недостаточное или неправильное образование
Слайд 27МЕТАБОЛИЗМ ЭЛАСТИНА
Синтез эластина:
Синтез и созревание белка эластина состоит из двух этапов:
МЕТАБОЛИЗМ ЭЛАСТИНА
Синтез эластина:
Синтез и созревание белка эластина состоит из двух этапов:

Слайд 28I. Внутриклеточный этап. Мономерная форма эластина синтезируется на рибосомах, связанных с ЭПР,
I. Внутриклеточный этап. Мономерная форма эластина синтезируется на рибосомах, связанных с ЭПР,

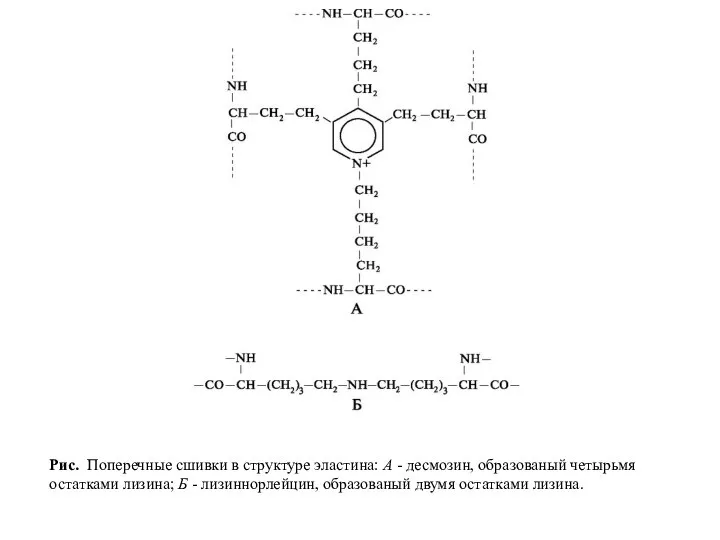
Слайд 29Рис. Поперечные сшивки в структуре эластина: А - десмозин, образованый четырьмя остатками лизина; Б - лизиннорлейцин,
Рис. Поперечные сшивки в структуре эластина: А - десмозин, образованый четырьмя остатками лизина; Б - лизиннорлейцин,
Слайд 30Аминокислотный состав эластина:
- 27% глицина;
- 19% аланина;
- 10% валина;
- 4,7% лейцина.
Благодаря наличию
Аминокислотный состав эластина:
- 27% глицина;
- 19% аланина;
- 10% валина;
- 4,7% лейцина.
Благодаря наличию

Слайд 31СТРУКТУРА ЭЛАСТИНА
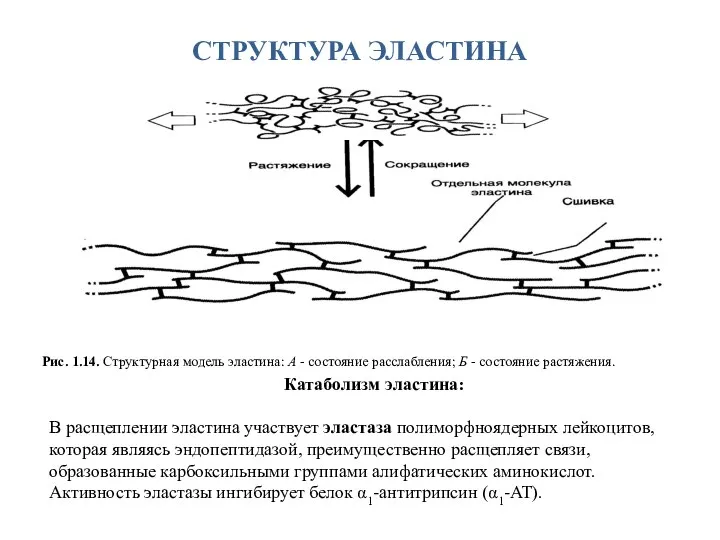
Рис. 1.14. Структурная модель эластина: А - состояние расслабления; Б - состояние растяжения.
Катаболизм эластина:
В расщеплении
СТРУКТУРА ЭЛАСТИНА
Рис. 1.14. Структурная модель эластина: А - состояние расслабления; Б - состояние растяжения.
Катаболизм эластина:
В расщеплении
Слайд 32НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА
При снижении образования десмозинов (или их отсутствии) поперечные сшивки образуются
НАРУШЕНИЯ СТРУКТУРЫ ЭЛАСТИНА
При снижении образования десмозинов (или их отсутствии) поперечные сшивки образуются

Слайд 33Состав и строение муцинов (от анг. mucus – слизь)
Гликопротеины, в которых
Состав и строение муцинов (от анг. mucus – слизь)
Гликопротеины, в которых

Слайд 34Синтез муцинов
Высокоспециализированные клетки поднижнечелюстных и подъязычных слюнных желез синтезируют белок муцин.
В
Синтез муцинов
Высокоспециализированные клетки поднижнечелюстных и подъязычных слюнных желез синтезируют белок муцин.
В

Слайд 35Присоединения GalNAc к остаткам серина и треонина обеспечивается GalNAс-трансферазой , происходит в
Присоединения GalNAc к остаткам серина и треонина обеспечивается GalNAс-трансферазой , происходит в

Слайд 36В слюне присутствует муцин-1 (Mr 250 кДа) и муцин-2 (Mr 100 кДа).
Внутренние
В слюне присутствует муцин-1 (Mr 250 кДа) и муцин-2 (Mr 100 кДа).
Внутренние

Слайд 37Муцины выполняют специфические функции:
Основные белки, обеспечивающие вязкость слюны. Муцины участвуют в образовании
Муцины выполняют специфические функции:
Основные белки, обеспечивающие вязкость слюны. Муцины участвуют в образовании

Слайд 38Протеогликаны и гликозаминогликаны
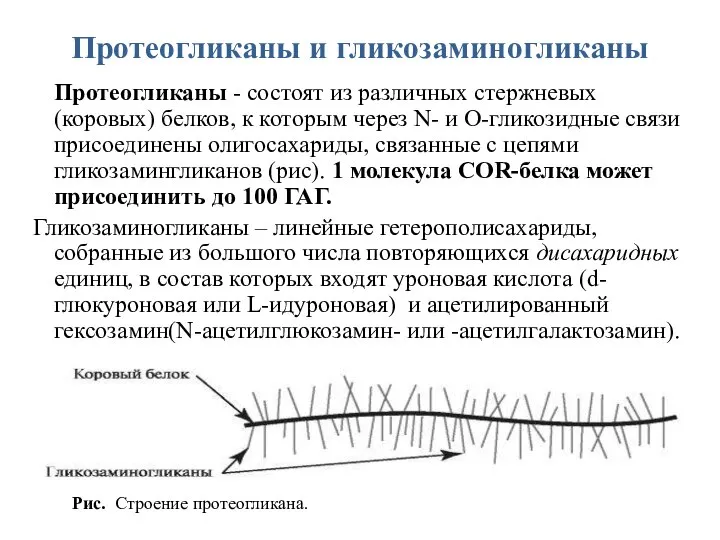
Протеогликаны - состоят из различных стержневых (коровых) белков, к которым через
Протеогликаны и гликозаминогликаны
Протеогликаны - состоят из различных стержневых (коровых) белков, к которым через
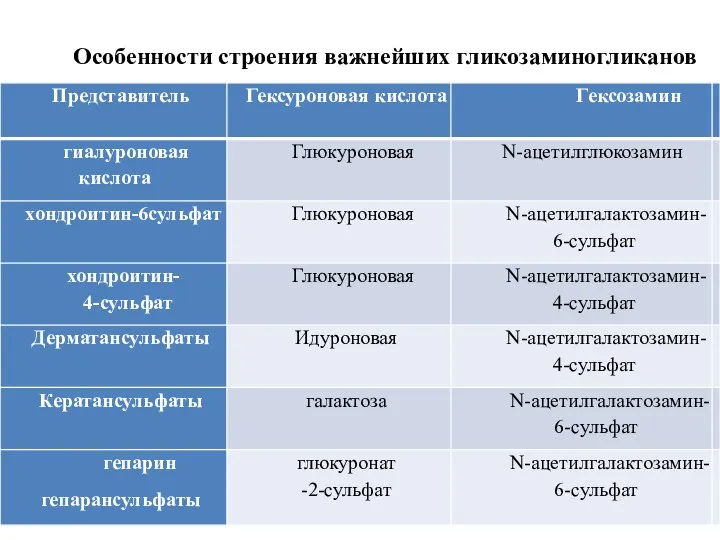
Слайд 39Особенности строения важнейших гликозаминогликанов
Особенности строения важнейших гликозаминогликанов
Слайд 40Все ГАГ делятся на две группы: связанные с коровым белком в составе
Все ГАГ делятся на две группы: связанные с коровым белком в составе
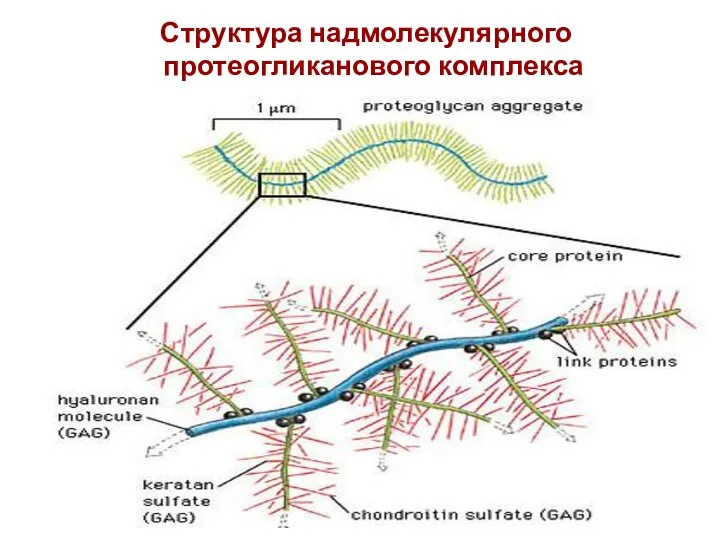
Слайд 41 Структура надмолекулярного протеогликанового комплекса
Структура надмолекулярного протеогликанового комплекса


Слайд 44 Аминосахара и гексуроновые кислоты синтезируются из глюкозы. Непосредственным же предшественником N-ацетилглюкозамина и
Аминосахара и гексуроновые кислоты синтезируются из глюкозы. Непосредственным же предшественником N-ацетилглюкозамина и

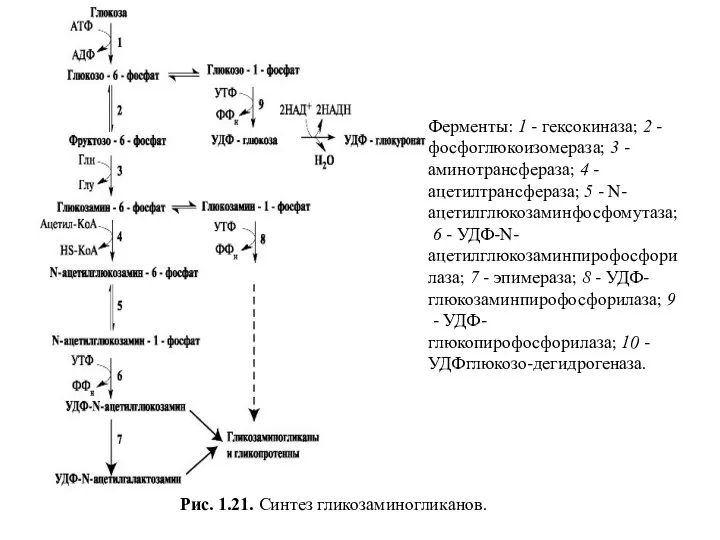
Слайд 45Рис. 1.21. Синтез гликозаминогликанов.
Ферменты: 1 - гексокиназа; 2 - фосфоглюкоизомераза; 3 - аминотрансфераза; 4 - ацетилтрансфераза; 5 - N-ацетилглюкозаминфосфомутаза; 6 - УДФ-N-ацетилглюкозаминпирофосфорилаза; 7 - эпимераза; 8 - УДФ- глюкозаминпирофосфорилаза; 9 -
Рис. 1.21. Синтез гликозаминогликанов.
Ферменты: 1 - гексокиназа; 2 - фосфоглюкоизомераза; 3 - аминотрансфераза; 4 - ацетилтрансфераза; 5 - N-ацетилглюкозаминфосфомутаза; 6 - УДФ-N-ацетилглюкозаминпирофосфорилаза; 7 - эпимераза; 8 - УДФ- глюкозаминпирофосфорилаза; 9 -
Слайд 46Разные протеогликаны отличаются размерами молекул, относительным содержанием белка и набором гликозаминогликанов.
Протеогликаны
Разные протеогликаны отличаются размерами молекул, относительным содержанием белка и набором гликозаминогликанов.
Протеогликаны

Слайд 47Распад протеогликанов
Распад протеогликанов - физиологический процесс, заключающийся в регулярном обновлении внеклеточных и
Распад протеогликанов
Распад протеогликанов - физиологический процесс, заключающийся в регулярном обновлении внеклеточных и

Слайд 482. Распад гликозаминогликанов
Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2 )
2. Распад гликозаминогликанов
Гликозаминогликаны отличаются высокой скоростью обмена: полупериод жизни (Т 1/2 )

Слайд 49
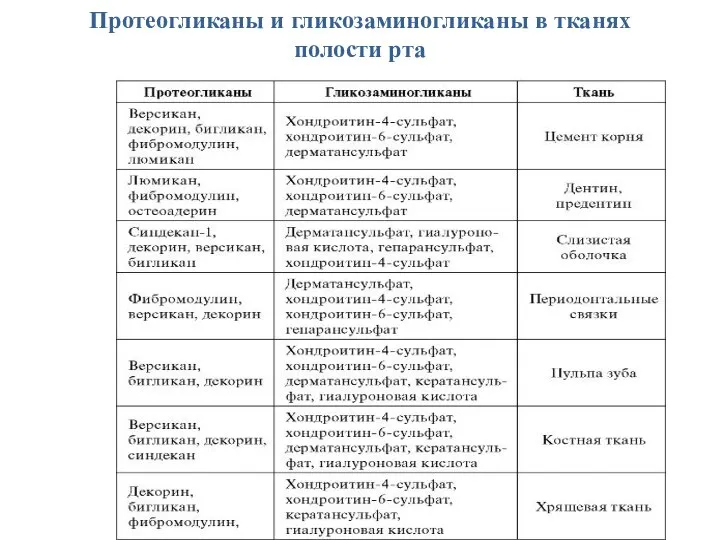
Протеогликаны и гликозаминогликаны в тканях полости рта
Протеогликаны и гликозаминогликаны в тканях полости рта
Слайд 50ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ
они являются структурными компонентами межклеточного матрикса;
протеогликаны и гликозаминогликаны специфически
ФУНКЦИИ ПРОТЕОГЛИКАНОВ И ГЛИКОЗАМИНОГЛИКАНОВ
они являются структурными компонентами межклеточного матрикса;
протеогликаны и гликозаминогликаны специфически

Слайд 51Мукополисахаридозы - тяжёлые наследственные заболевания, обусловленые дефектами гидролаз, участвующих в катаболизме гликозаминогликанов. В
Мукополисахаридозы - тяжёлые наследственные заболевания, обусловленые дефектами гидролаз, участвующих в катаболизме гликозаминогликанов. В

Слайд 52Болезни, связанные с нарушением метаболизма гликозаминогликанов
Болезни, связанные с нарушением метаболизма гликозаминогликанов
 Высшие споровые растения
Высшие споровые растения Митоз и мейоз
Митоз и мейоз Бычий цепень. Микробиология
Бычий цепень. Микробиология Сердце, сосуды. Фазы работы сердца
Сердце, сосуды. Фазы работы сердца Гуморальная регуляции организма
Гуморальная регуляции организма Роль животных в жизни человека
Роль животных в жизни человека Презентация на тему Какая у нас память
Презентация на тему Какая у нас память  Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений
Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений Обмен веществ
Обмен веществ Растения. Царство растения
Растения. Царство растения Головной мозг человека. (Лекция 6)
Головной мозг человека. (Лекция 6) Птицы - наши друзья
Птицы - наши друзья Органы пищеварения человека. Строение и значение зубов
Органы пищеварения человека. Строение и значение зубов Презентация на тему ОСЕНЬ И ЗИМА В ЖИЗНИ РАСТЕНИЙ
Презентация на тему ОСЕНЬ И ЗИМА В ЖИЗНИ РАСТЕНИЙ 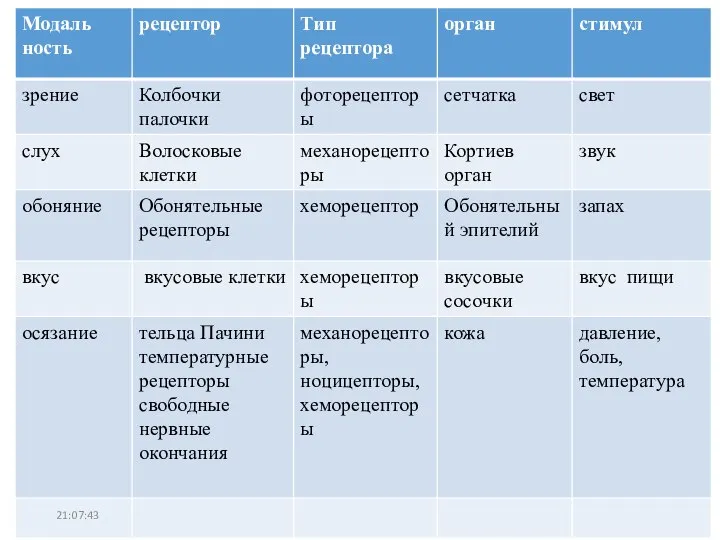 Механорецепторы. Тип рецептора
Механорецепторы. Тип рецептора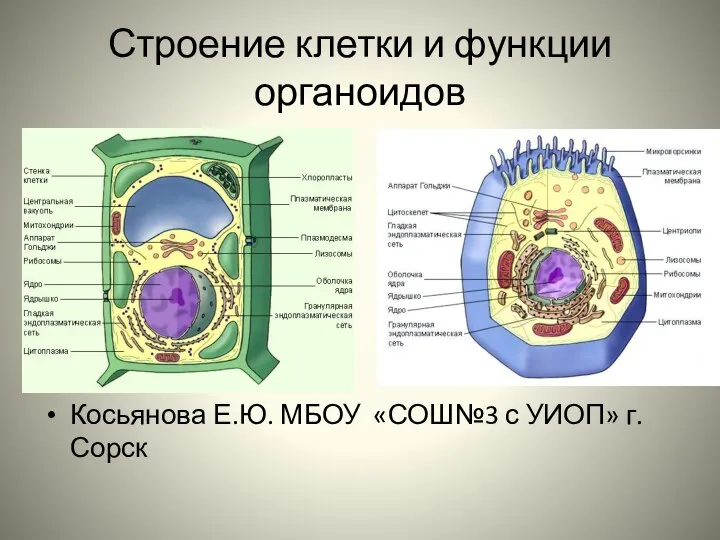 Строение клетки и функции органоидов. Органеллы клетки
Строение клетки и функции органоидов. Органеллы клетки Удивительные и загадочные Ахатины (Achatina fulica)
Удивительные и загадочные Ахатины (Achatina fulica) Создание гербария Травки-муравки
Создание гербария Травки-муравки Коллекция как проект
Коллекция как проект Пропорции и строение фигуры человека
Пропорции и строение фигуры человека Внутриутробное развитие у млекопитающих. Виды плаценты млекопитающих
Внутриутробное развитие у млекопитающих. Виды плаценты млекопитающих Сеченовка экзамен
Сеченовка экзамен Ткани растений. 5 класс
Ткани растений. 5 класс Фотосинтез
Фотосинтез История генетики. Основные понятия генетики (урок 19)
История генетики. Основные понятия генетики (урок 19) Реципрокті (жинақталған) тежелу
Реципрокті (жинақталған) тежелу Органоиды клетки и их функции (§ 8)
Органоиды клетки и их функции (§ 8) Осьминог. Анотомия и физиология
Осьминог. Анотомия и физиология