- Энергообмен и его регуляция

Содержание
- 3. Гликолиз + цикл Кребса + окислительное фосфорилирование → до 38 моль АТФ
- 4. Преобладающими субстратами для образования АТФ в митохондриях служат – НАДН и отчасти - НАДФН Разность в
- 5. НАДH НАД H+ H+ H+ 2H++1/2O2 H2O H+ АДФ АТФ H+ межмембранное пространство внутренняя мембрана митохондрии
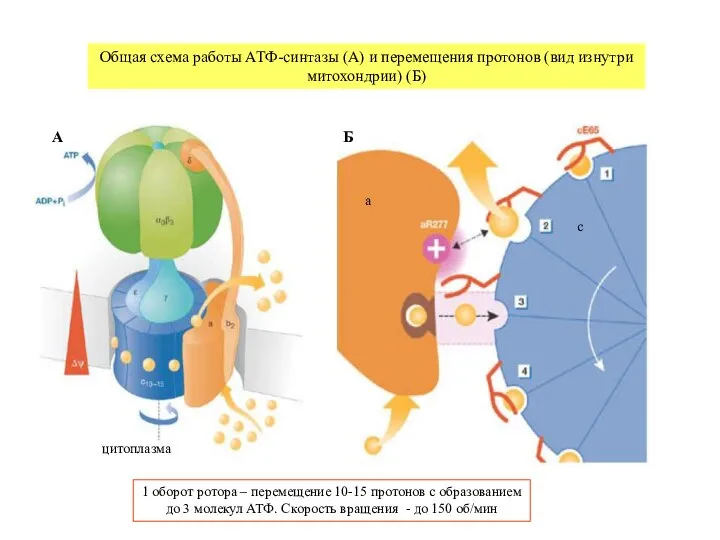
- 6. цитоплазма Общая схема работы АТФ-синтазы (А) и перемещения протонов (вид изнутри митохондрии) (Б) 1 оборот ротора
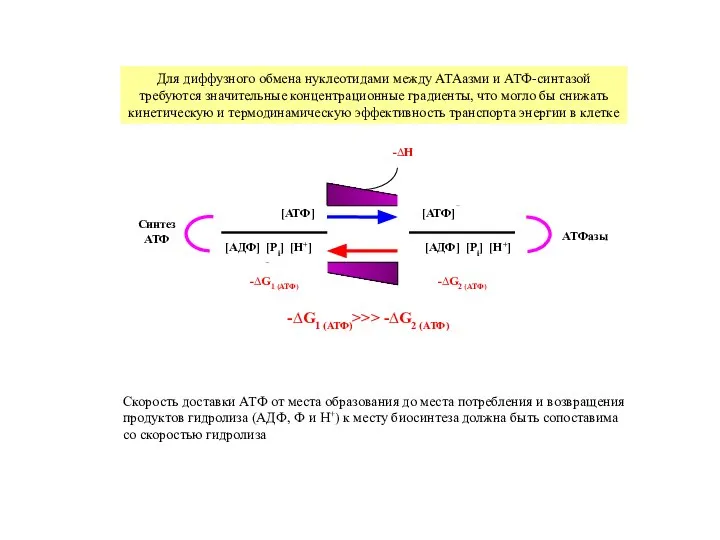
- 7. Скорость доставки АТФ от места образования до места потребления и возвращения продуктов гидролиза (АДФ, Ф и
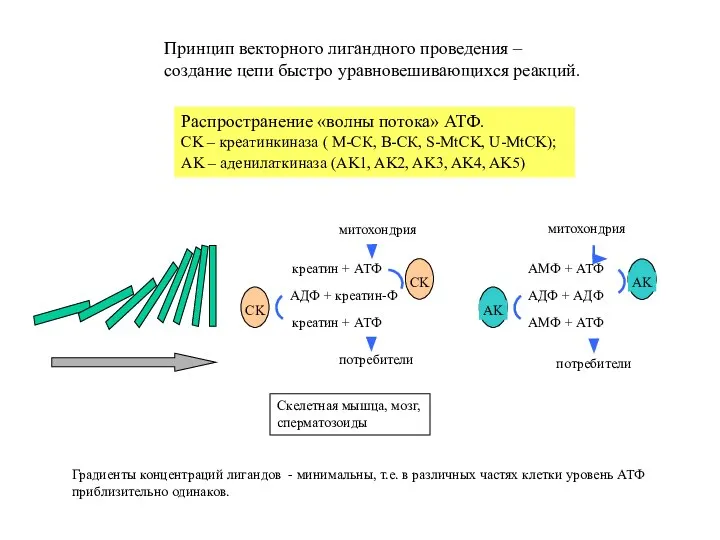
- 8. креатин + АТФ АДФ + креатин-Ф креатин + АТФ потребители митохондрия митохондрия АМФ + АТФ АДФ
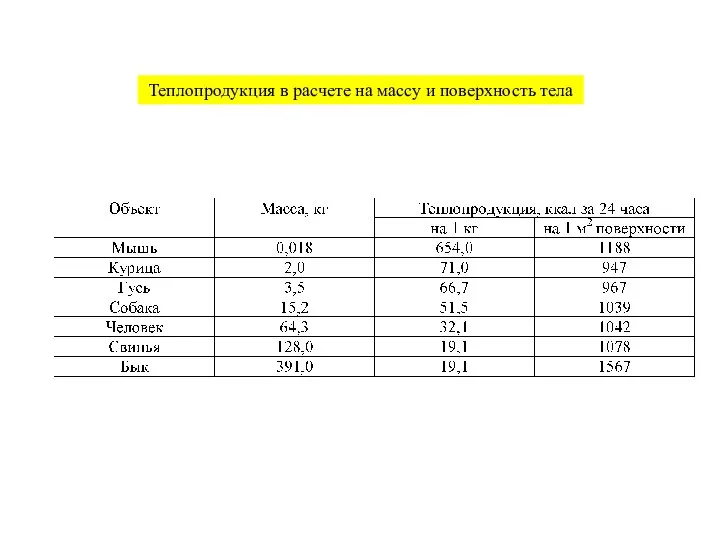
- 10. Интегральные показатели энергообмена
- 12. Регистрация газообмена и выделения H2O Регистрация теплообмена Методами прямой и непрямой (по газообмену) калориметрии, а также
- 14. Факториальный индикатор обмена равен соотношению энергозатрат при максимальной активности и в состоянии покоя
- 15. Жир – 0-3% Углеводы – 5-10% Белки – 20-30% Алкоголь -10-30% Доля энергетической ценности продуктов, затрачиваемая
- 16. Интенсивность энергообмена определяется : температурными условиями окружающей среды, доступностью пищи
- 17. Терморегуляция
- 18. Относительность изотермии: температура разных частей тела – от 24,5̊ до 38,5̊ суточные колебания – от 36,4̊
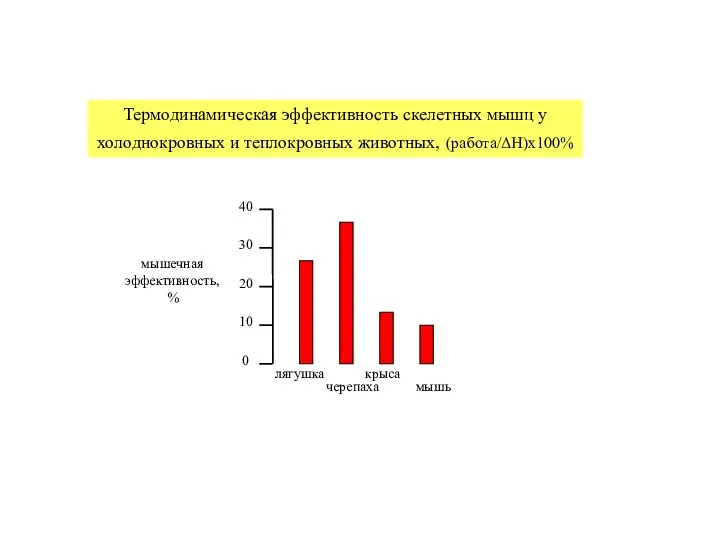
- 19. Термодинамическая эффективность скелетных мышц у холоднокровных и теплокровных животных, (работа/ΔH)x100%
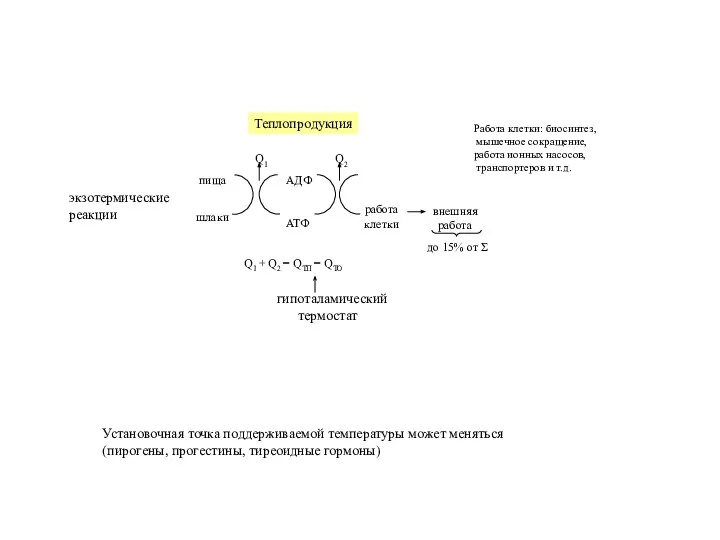
- 21. экзотермические реакции Работа клетки: биосинтез, мышечное сокращение, работа ионных насосов, транспортеров и т.д. гипоталамический термостат Установочная
- 22. Термонейтральная область Теплопродукция недрожательный, дрожательный термогенез
- 23. НАДH НАД H+ H+ H+ 2H++1/2O2 H2O H+ H+ H+ UCP АДФ АТФ H+ H+ H+
- 24. Функции рассопрягающих белков UCP 1 - 5 UCP2 вызывает ↓АТФ/АДФ → ↓свободных радикалов (защитная функция от
- 25. Гормональная регуляция термогенеза
- 26. Регуляция экспрессии рассопрягающего белка UCP1 в буром жире Синергизм катехоламинов и тиреоидных гормонов: только норадреналин –
- 27. Синергизм тиреоидных гормонов и симпатической стимуляции в индукции термогенеза в буром жире Клинические данные: Гипотиреоз –
- 28. nNOS nNOS’ nNOS’ NO Транспортеры электронов T3 T3 митохондрия Тиреоидные гормоны снимают ингибирующее действие нитроксидсинтазы на
- 29. TRE T3 NRF1 TRE PGC-1α NRF-RE цитохром c mtTFA 1-я волна 2-я волна тканеспецифичная экспрессия Действие
- 30. Температура тела Время, час 37ºС 30ºС 0 24 22ºС 4ºС Температура окружающей среды TRα/β(-/-) Интактные Холодовая
- 31. Потребление O2 мл/мин/кг Температура окружающей среды, ºС 0 37 Нижние критические температуры Поддерживаемая температура тела 0
- 32. 1 100 10000 Норадреналин, нМ Прирост потребления O2 Интактные TRα/β(-/-) Тиреоидные гормоны через свои рецепторы сенсибилизируют
- 33. 1 100 0 10000 +T3 Норадреналин, нМ Продукция цАМФ +GC-1 Без гормона Избирательный агонист рецептора β
- 34. Дейодиназа D2 в клетках бурого жира повышает уровень T3, необходимого для активации TRα, сенсибилизирующего клетки к
- 35. Каталитическая реакция дейодирования T4 стимулирует убиквитинилирование и инактивацию дейодиназы D2. Норадреналин повышает активность дейодиназы D2, индуцируя
- 36. D2 – в мышцах человека, но не грызунов.
- 37. 0 Усилие (мН/мм2) 3000 Тепло (мВт/г) 40 гипотиреоз эутиреоз медленная мышца быстрая мышца Влияние тиреоидных гормонов
- 38. Мишени тиреоидных гормонов в буром жире: UCP1 белки дыхательной цепи переноса электронов широкий спектр белков митохондрий
- 39. Регуляция пищевого поведения
- 40. Nutrition. 2009 Nov-Dec;25(11-12):1186-92. 30-Дневные крысы массой 100 г получали диету с нормальным содержанием белка (17%) и
- 41. Взаимосвязи центров терморегуляции и пищевого поведения
- 42. Грелин Обестатин Области мозга, связанные с потреблением пищи: Аркуатное ядро (не защищено гематоэнцефалическим барьером; возможна регуляция
- 43. Нейропептид Y – NPY организатор пищевого поведения Рецепторы - Y1 и Y5 (антагонисты рецепторов снимают ночной
- 44. «Голодная» секреция грелина желудком
- 45. Голодание, грелин Стимуляция нейронов аркуатного ядра NPY/ AgRP Высвобождение NPY и AgRP Прием пищи, лептин Стимуляция
- 46. Frontiers in Systems Neuroscience, 2015, v 9, Article 150
- 47. Figure . Leptin and insulin signaling pathways in the hypothalamus Leptin binding to its receptor (OB-Rb)
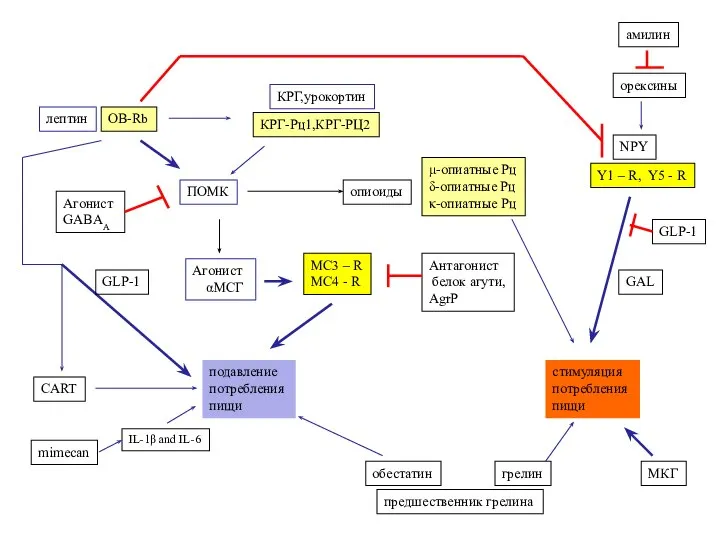
- 48. MC3 – R MC4 - R Агонист αМСГ Антагонист белок агути, AgrP ПОМК лептин КРГ,урокортин опиоиды
- 49. Ожирение
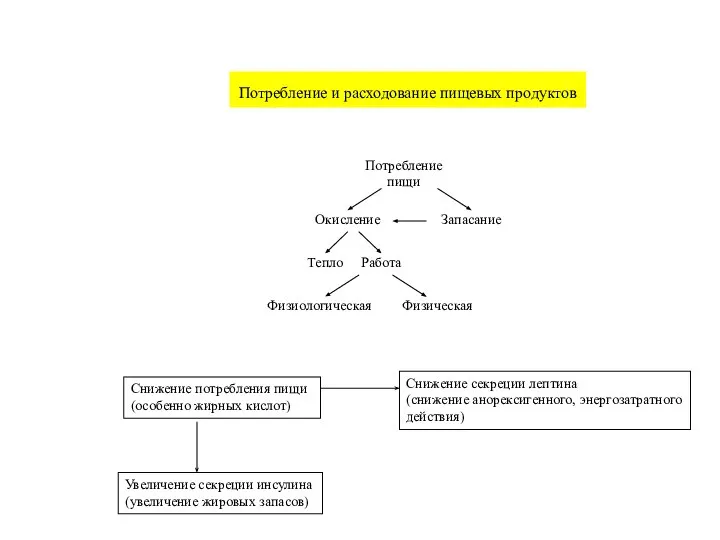
- 50. Снижение потребления пищи (особенно жирных кислот) Снижение секреции лептина (снижение анорексигенного, энергозатратного действия) Увеличение секреции инсулина
- 51. Детское ожирение Увеличение количества адипоцитов в результате усиленной дифференцировки преадипоцитов в адипоциты под действием PPARγ, активированных
- 53. Скачать презентацию
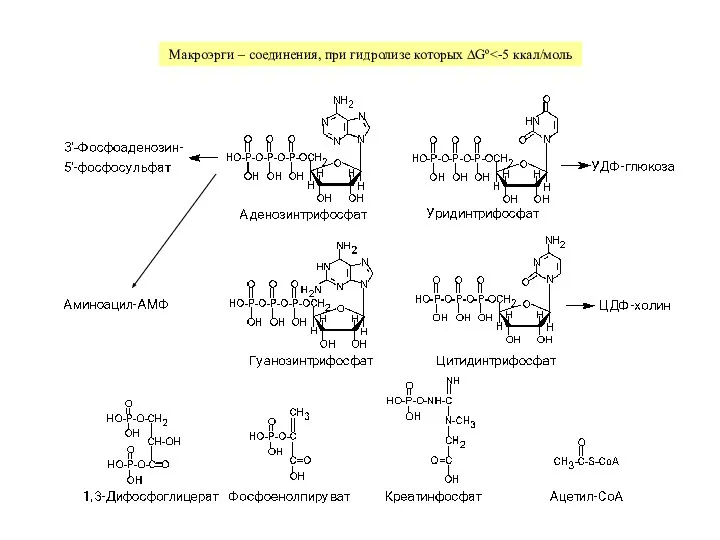
Слайд 3Гликолиз + цикл Кребса + окислительное фосфорилирование → до 38 моль АТФ
Гликолиз + цикл Кребса + окислительное фосфорилирование → до 38 моль АТФ
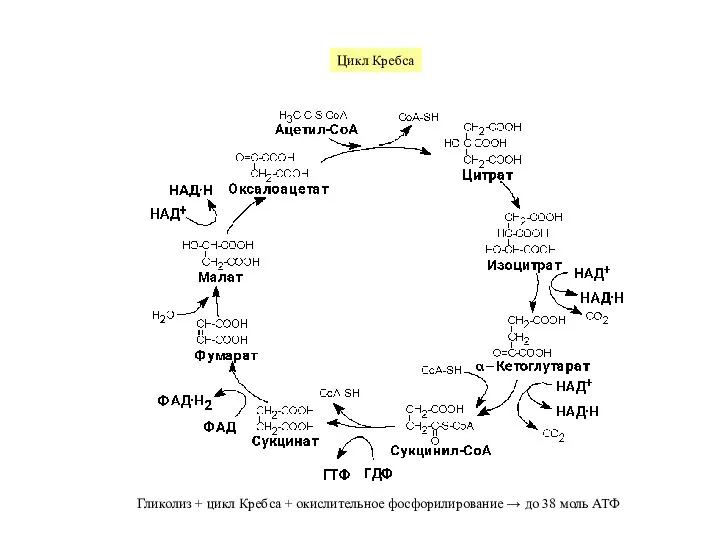
Слайд 4Преобладающими субстратами для образования АТФ в митохондриях служат – НАДН и отчасти
Преобладающими субстратами для образования АТФ в митохондриях служат – НАДН и отчасти
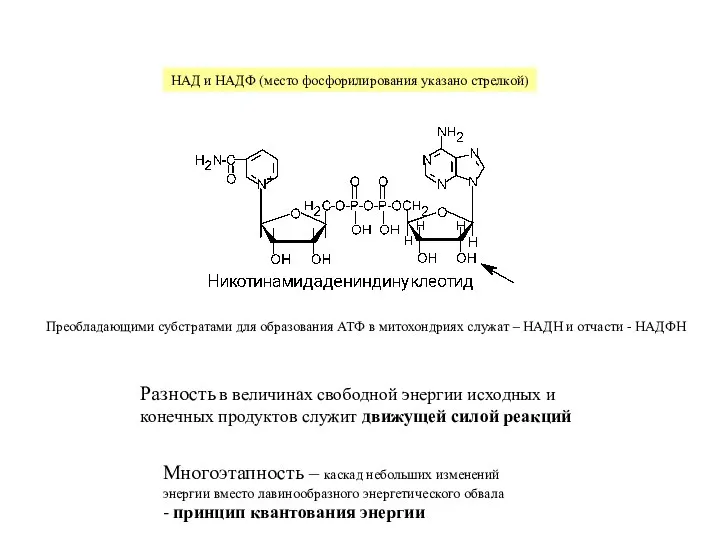
Слайд 5НАДH
НАД
H+
H+
H+
2H++1/2O2
H2O
H+
АДФ
АТФ
H+
межмембранное
пространство
внутренняя мембрана
митохондрии
матрикс
цепь переноса электронов
АТФ-синтаза
Цепь переноса электронов, фосфорилирование
Внутренняя часть
митохондрии
НАДH
НАД
H+
H+
H+
2H++1/2O2
H2O
H+
АДФ
АТФ
H+
межмембранное
пространство
внутренняя мембрана
митохондрии
матрикс
цепь переноса электронов
АТФ-синтаза
Цепь переноса электронов, фосфорилирование
Внутренняя часть
митохондрии
Слайд 6цитоплазма
Общая схема работы АТФ-синтазы (А) и перемещения протонов (вид изнутри митохондрии) (Б)
цитоплазма
Общая схема работы АТФ-синтазы (А) и перемещения протонов (вид изнутри митохондрии) (Б)
Слайд 7Скорость доставки АТФ от места образования до места потребления и возвращения
продуктов гидролиза
Скорость доставки АТФ от места образования до места потребления и возвращения
продуктов гидролиза
Слайд 8креатин + АТФ
АДФ + креатин-Ф
креатин + АТФ
потребители
митохондрия
митохондрия
АМФ + АТФ
АДФ + АДФ
АМФ +
креатин + АТФ
АДФ + креатин-Ф
креатин + АТФ
потребители
митохондрия
митохондрия
АМФ + АТФ
АДФ + АДФ
АМФ +
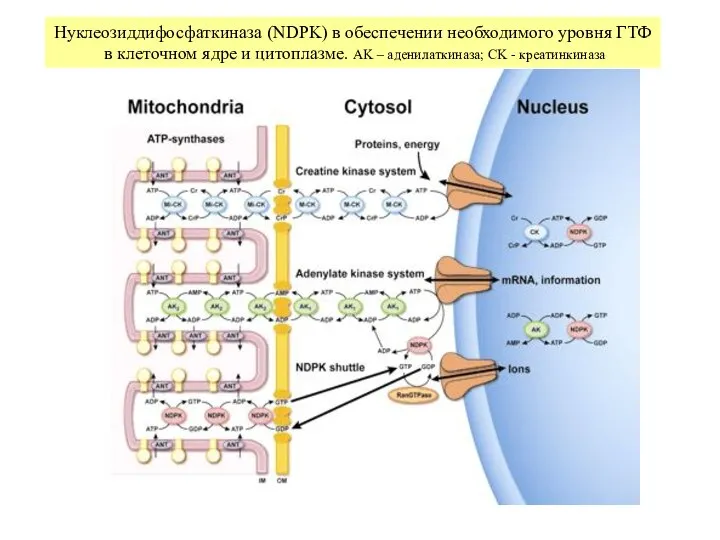
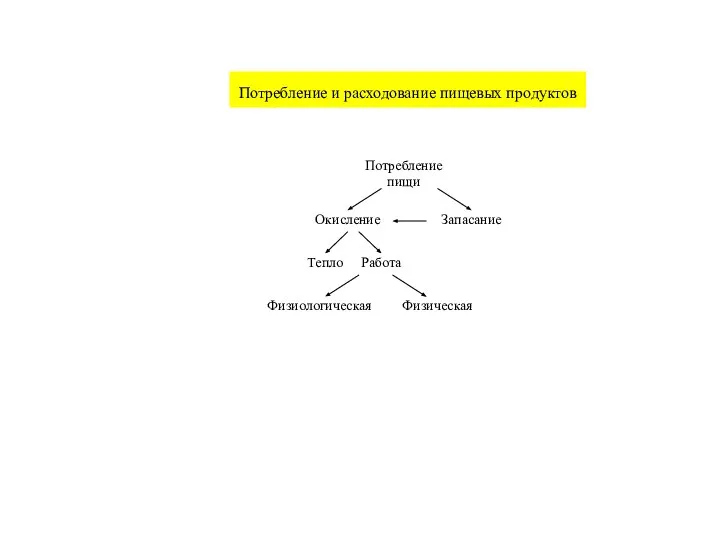
Слайд 10Интегральные показатели энергообмена
Интегральные показатели энергообмена

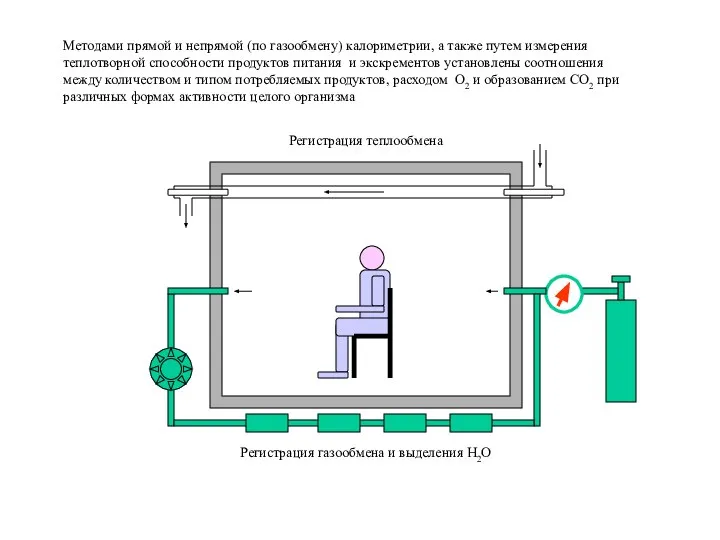
Слайд 12Регистрация газообмена и выделения H2O
Регистрация теплообмена
Методами прямой и непрямой (по газообмену) калориметрии,
Регистрация газообмена и выделения H2O
Регистрация теплообмена
Методами прямой и непрямой (по газообмену) калориметрии,
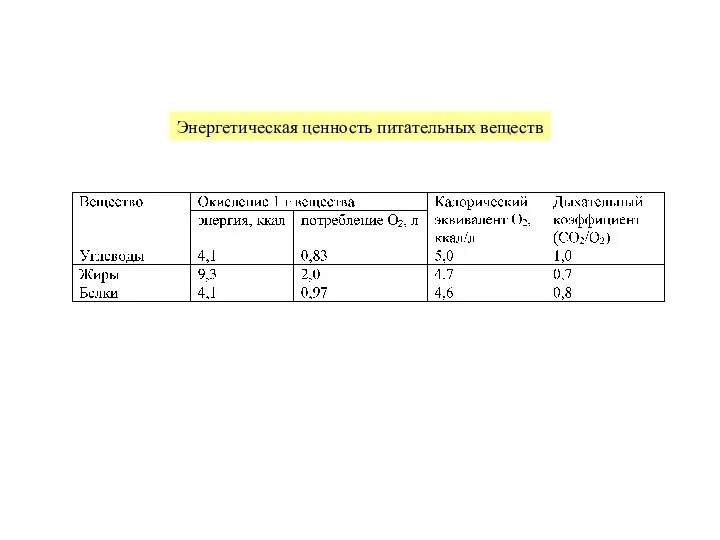
Слайд 14Факториальный индикатор обмена равен соотношению энергозатрат при максимальной
активности и в состоянии
Факториальный индикатор обмена равен соотношению энергозатрат при максимальной
активности и в состоянии

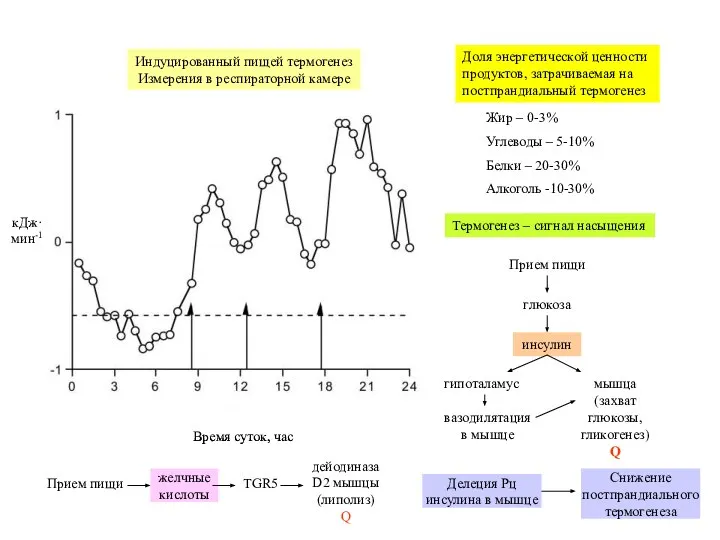
Слайд 15Жир – 0-3%
Углеводы – 5-10%
Белки – 20-30%
Алкоголь -10-30%
Доля энергетической ценности продуктов, затрачиваемая
Жир – 0-3%
Углеводы – 5-10%
Белки – 20-30%
Алкоголь -10-30%
Доля энергетической ценности продуктов, затрачиваемая
Слайд 16Интенсивность энергообмена определяется :
температурными условиями окружающей среды,
доступностью пищи
Интенсивность энергообмена определяется :
температурными условиями окружающей среды,
доступностью пищи

Слайд 17Терморегуляция
Терморегуляция

Слайд 18Относительность изотермии:
температура разных частей тела – от 24,5̊ до 38,5̊
суточные колебания
Относительность изотермии:
температура разных частей тела – от 24,5̊ до 38,5̊
суточные колебания

Слайд 19Термодинамическая эффективность скелетных мышц у холоднокровных и теплокровных животных, (работа/ΔH)x100%
Термодинамическая эффективность скелетных мышц у холоднокровных и теплокровных животных, (работа/ΔH)x100%
Слайд 21экзотермические
реакции
Работа клетки: биосинтез,
мышечное сокращение,
работа ионных насосов,
транспортеров и т.д.
гипоталамический
термостат
Установочная
экзотермические
реакции
Работа клетки: биосинтез,
мышечное сокращение,
работа ионных насосов,
транспортеров и т.д.
гипоталамический
термостат
Установочная
Слайд 22Термонейтральная
область
Теплопродукция
недрожательный,
дрожательный
термогенез
Термонейтральная
область
Теплопродукция
недрожательный,
дрожательный
термогенез
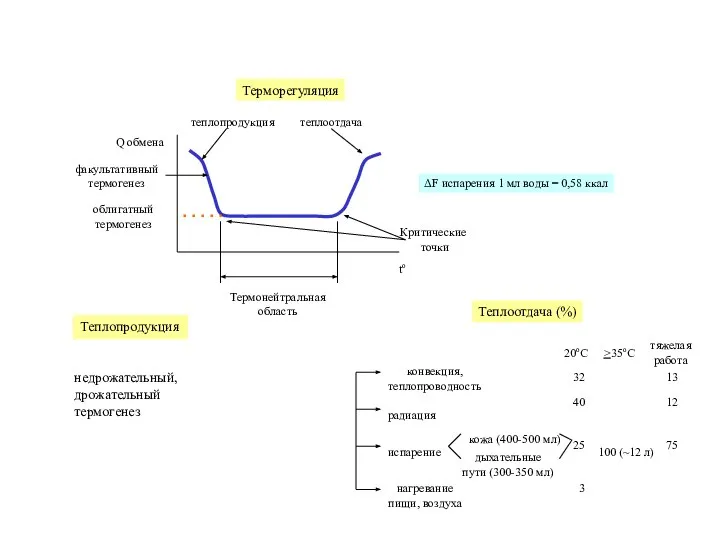
Слайд 23НАДH
НАД
H+
H+
H+
2H++1/2O2
H2O
H+
H+
H+
UCP
АДФ
АТФ
H+
H+
H+
“утечки” 20-25% термогенеза в состоянии покоя
межмембранное
пространство
внутренняя мембрана
матрикс
цепь переноса электронов
АТФ-синтаза
Цепь переноса электронов, фосфорилирование
НАДH
НАД
H+
H+
H+
2H++1/2O2
H2O
H+
H+
H+
UCP
АДФ
АТФ
H+
H+
H+
“утечки” 20-25% термогенеза в состоянии покоя
межмембранное
пространство
внутренняя мембрана
матрикс
цепь переноса электронов
АТФ-синтаза
Цепь переноса электронов, фосфорилирование
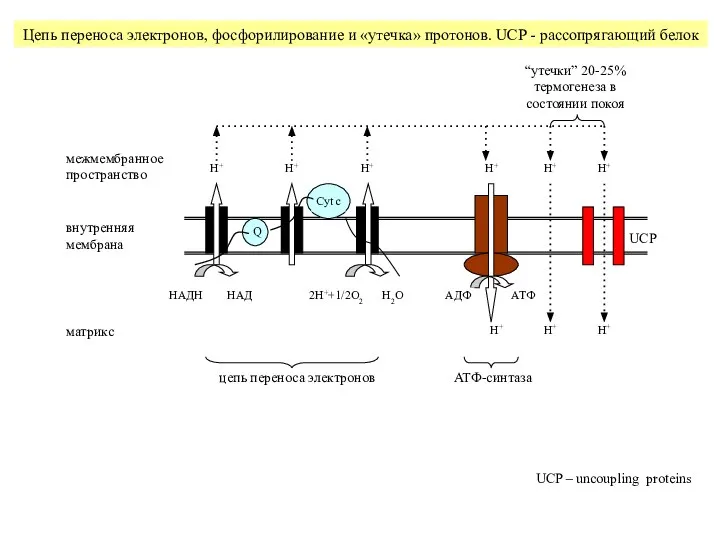
Слайд 24Функции рассопрягающих белков UCP 1 - 5
UCP2 вызывает ↓АТФ/АДФ → ↓свободных
Функции рассопрягающих белков UCP 1 - 5
UCP2 вызывает ↓АТФ/АДФ → ↓свободных

Слайд 25Гормональная регуляция термогенеза
Гормональная регуляция термогенеза

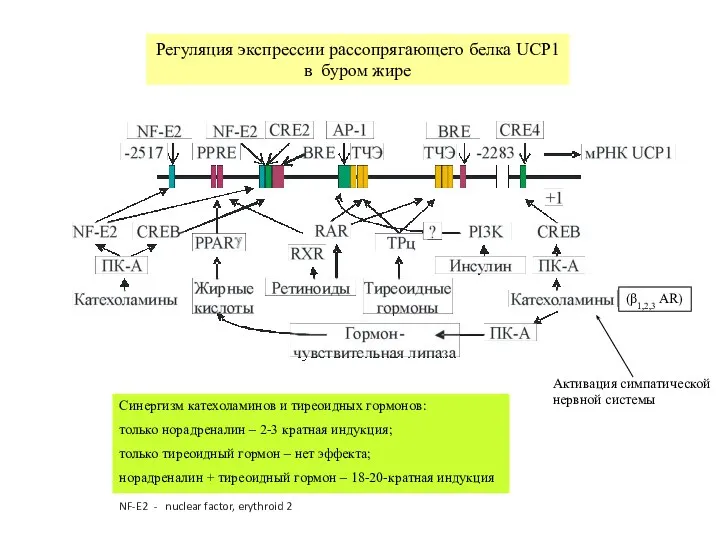
Слайд 26Регуляция экспрессии рассопрягающего белка UCP1
в буром жире
Синергизм катехоламинов и тиреоидных гормонов:
только
Регуляция экспрессии рассопрягающего белка UCP1
в буром жире
Синергизм катехоламинов и тиреоидных гормонов:
только
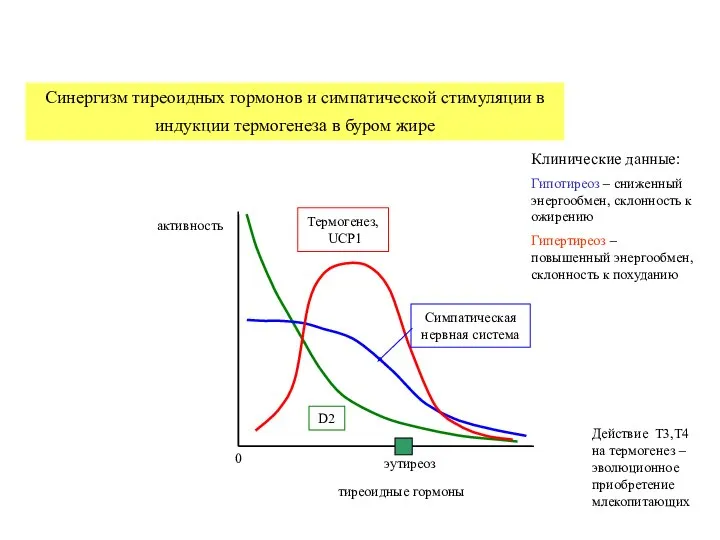
Слайд 27Синергизм тиреоидных гормонов и симпатической стимуляции в индукции термогенеза в буром жире
Синергизм тиреоидных гормонов и симпатической стимуляции в индукции термогенеза в буром жире
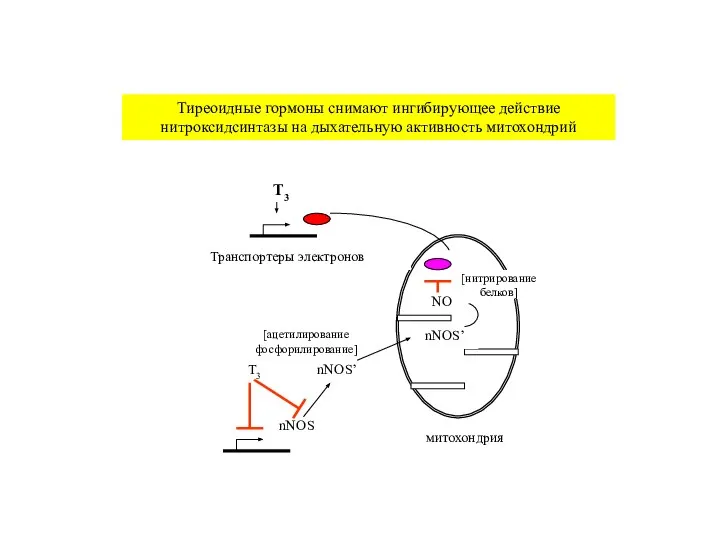
Слайд 28nNOS
nNOS’
nNOS’
NO
Транспортеры электронов
T3
T3
митохондрия
Тиреоидные гормоны снимают ингибирующее действие нитроксидсинтазы на дыхательную активность митохондрий
[нитрирование
nNOS
nNOS’
nNOS’
NO
Транспортеры электронов
T3
T3
митохондрия
Тиреоидные гормоны снимают ингибирующее действие нитроксидсинтазы на дыхательную активность митохондрий
[нитрирование
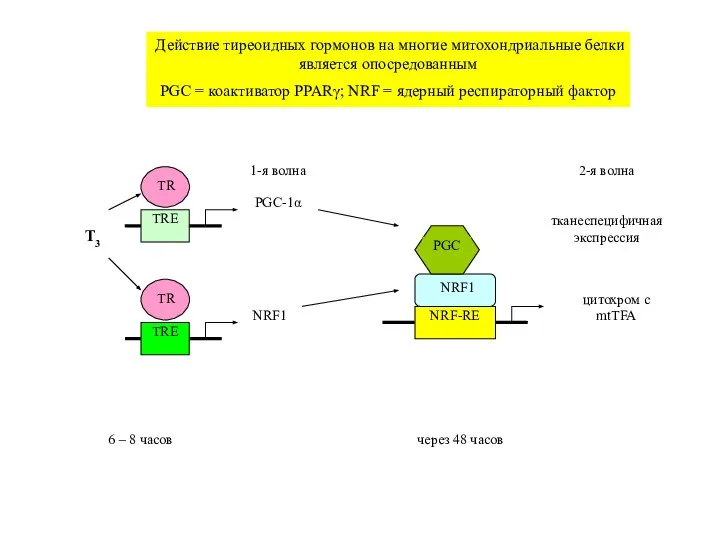
Слайд 29TRE
T3
NRF1
TRE
PGC-1α
NRF-RE
цитохром c
mtTFA
1-я волна
2-я волна
тканеспецифичная
экспрессия
Действие тиреоидных гормонов на многие митохондриальные белки является опосредованным
TRE
T3
NRF1
TRE
PGC-1α
NRF-RE
цитохром c
mtTFA
1-я волна
2-я волна
тканеспецифичная
экспрессия
Действие тиреоидных гормонов на многие митохондриальные белки является опосредованным
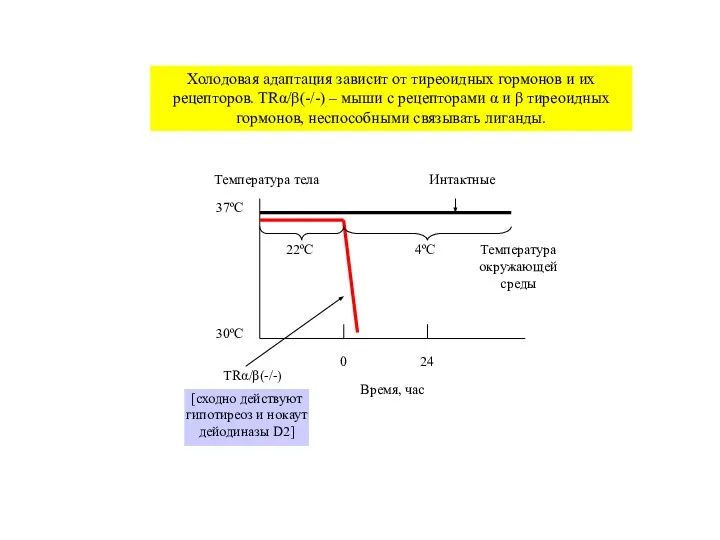
Слайд 30Температура тела
Время, час
37ºС
30ºС
0
24
22ºС
4ºС
Температура
окружающей
среды
TRα/β(-/-)
Интактные
Холодовая адаптация зависит от тиреоидных гормонов и их
Температура тела
Время, час
37ºС
30ºС
0
24
22ºС
4ºС
Температура
окружающей
среды
TRα/β(-/-)
Интактные
Холодовая адаптация зависит от тиреоидных гормонов и их
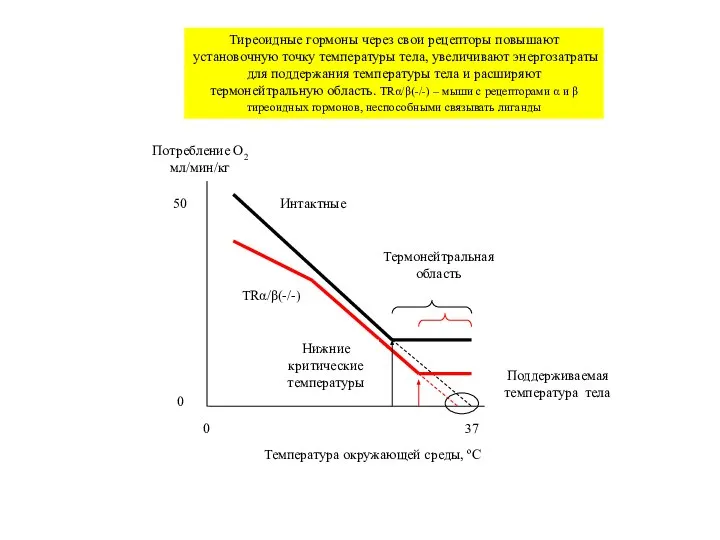
Слайд 31Потребление O2
мл/мин/кг
Температура окружающей среды, ºС
0
37
Нижние критические температуры
Поддерживаемая
температура тела
0
50
Интактные
TRα/β(-/-)
Термонейтральная область
Тиреоидные гормоны через свои
Потребление O2
мл/мин/кг
Температура окружающей среды, ºС
0
37
Нижние критические температуры
Поддерживаемая
температура тела
0
50
Интактные
TRα/β(-/-)
Термонейтральная область
Тиреоидные гормоны через свои
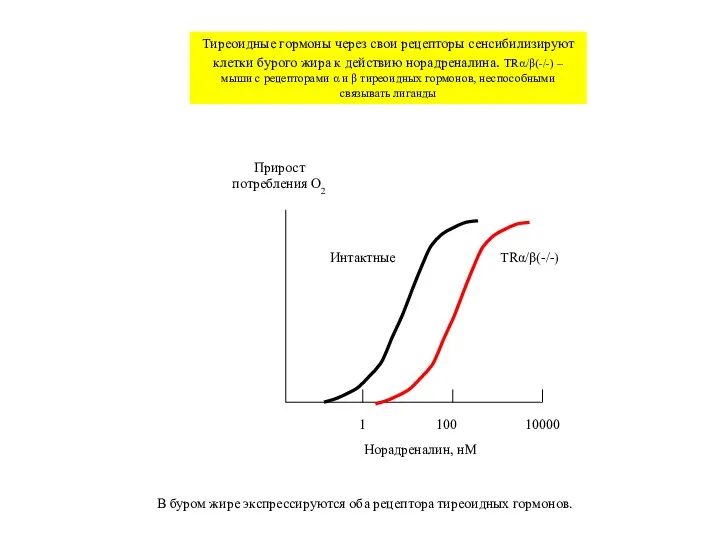
Слайд 321
100
10000
Норадреналин, нМ
Прирост потребления O2
Интактные
TRα/β(-/-)
Тиреоидные гормоны через свои рецепторы сенсибилизируют клетки бурого жира
1
100
10000
Норадреналин, нМ
Прирост потребления O2
Интактные
TRα/β(-/-)
Тиреоидные гормоны через свои рецепторы сенсибилизируют клетки бурого жира
Слайд 331
100
0
10000
+T3
Норадреналин, нМ
Продукция цАМФ
+GC-1
Без гормона
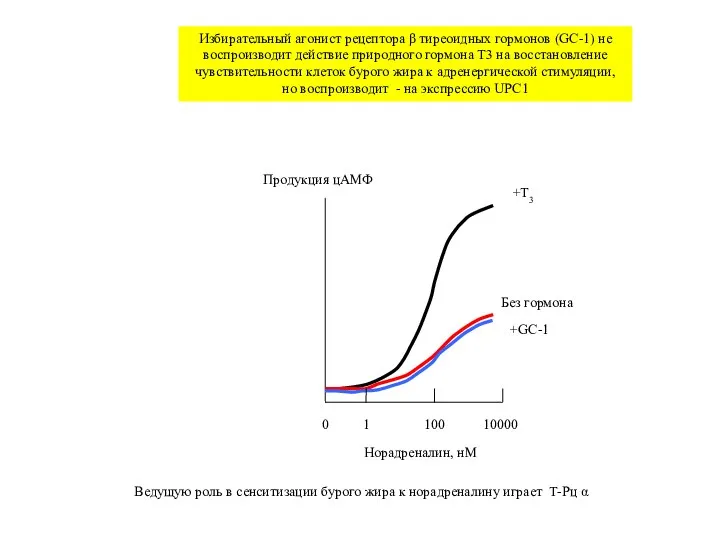
Избирательный агонист рецептора β тиреоидных гормонов (GC-1) не воспроизводит
1
100
0
10000
+T3
Норадреналин, нМ
Продукция цАМФ
+GC-1
Без гормона
Избирательный агонист рецептора β тиреоидных гормонов (GC-1) не воспроизводит
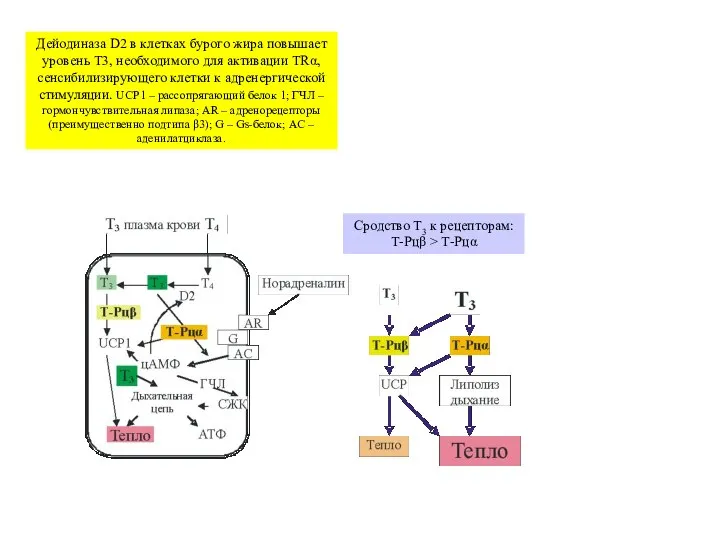
Слайд 34Дейодиназа D2 в клетках бурого жира повышает уровень T3, необходимого для активации
Дейодиназа D2 в клетках бурого жира повышает уровень T3, необходимого для активации
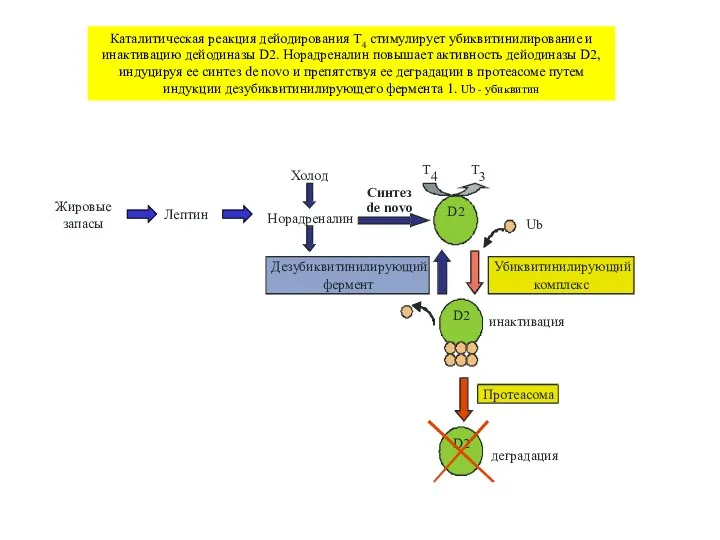
Слайд 35Каталитическая реакция дейодирования T4 стимулирует убиквитинилирование и инактивацию дейодиназы D2. Норадреналин повышает
Каталитическая реакция дейодирования T4 стимулирует убиквитинилирование и инактивацию дейодиназы D2. Норадреналин повышает
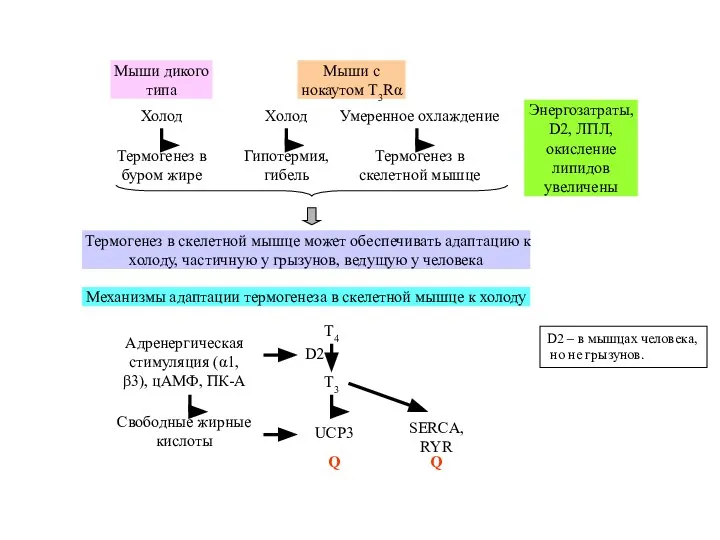
Слайд 36D2 – в мышцах человека,
но не грызунов.
D2 – в мышцах человека,
но не грызунов.
Слайд 370
Усилие (мН/мм2)
3000
Тепло (мВт/г)
40
гипотиреоз
эутиреоз
медленная мышца
быстрая мышца
Влияние тиреоидных гормонов на термогенез в покоящейся и
0
Усилие (мН/мм2)
3000
Тепло (мВт/г)
40
гипотиреоз
эутиреоз
медленная мышца
быстрая мышца
Влияние тиреоидных гормонов на термогенез в покоящейся и
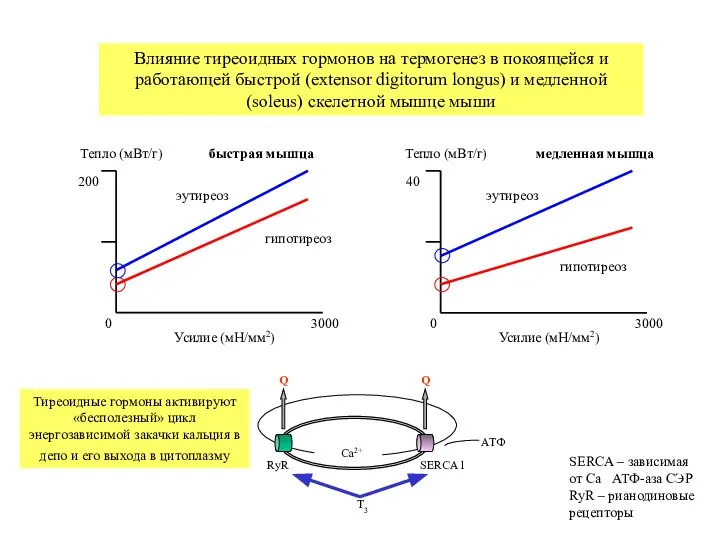
Слайд 38
Мишени тиреоидных гормонов в буром жире:
UCP1
белки дыхательной цепи переноса электронов
широкий спектр
Мишени тиреоидных гормонов в буром жире:
UCP1
белки дыхательной цепи переноса электронов
широкий спектр

Слайд 39Регуляция пищевого поведения
Регуляция пищевого поведения

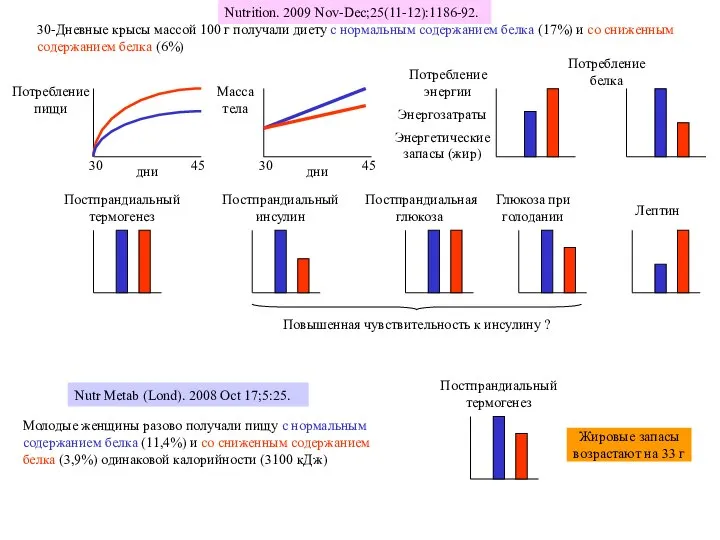
Слайд 40Nutrition. 2009 Nov-Dec;25(11-12):1186-92.
30-Дневные крысы массой 100 г получали диету с нормальным
Nutrition. 2009 Nov-Dec;25(11-12):1186-92.
30-Дневные крысы массой 100 г получали диету с нормальным
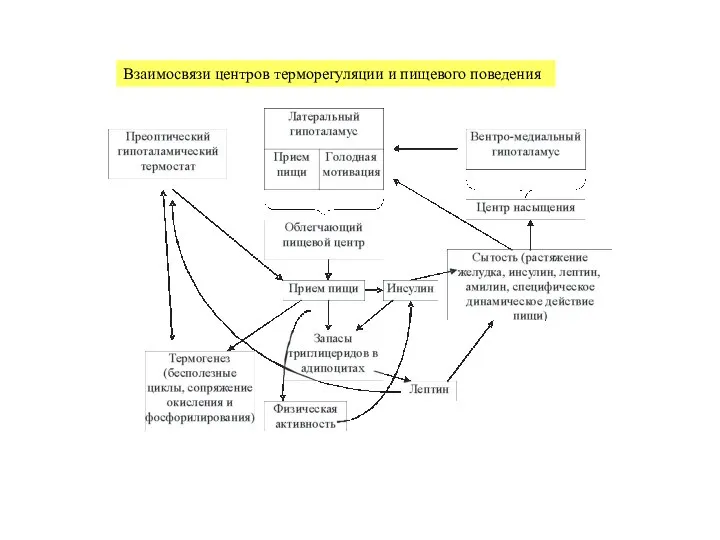
Слайд 41Взаимосвязи центров терморегуляции и пищевого поведения
Взаимосвязи центров терморегуляции и пищевого поведения
Слайд 42Грелин
Обестатин
Области мозга, связанные с потреблением пищи:
Аркуатное ядро (не защищено гематоэнцефалическим барьером; возможна
Грелин
Обестатин
Области мозга, связанные с потреблением пищи:
Аркуатное ядро (не защищено гематоэнцефалическим барьером; возможна

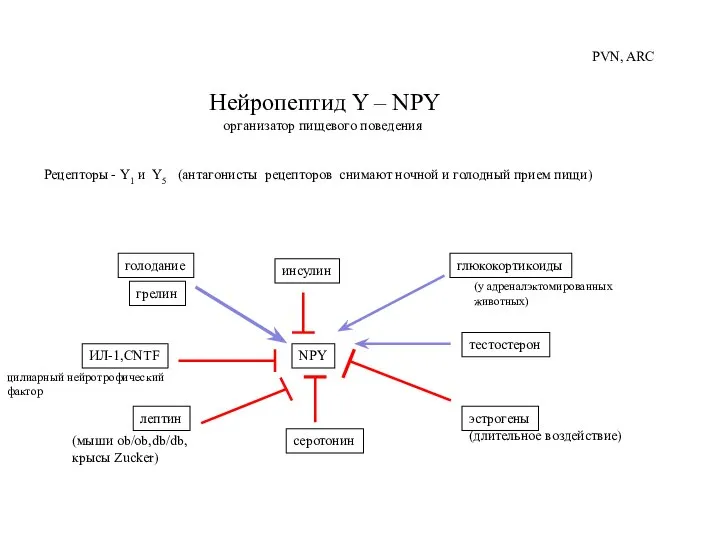
Слайд 43Нейропептид Y – NPY
организатор пищевого поведения
Рецепторы - Y1 и Y5 (антагонисты
Нейропептид Y – NPY
организатор пищевого поведения
Рецепторы - Y1 и Y5 (антагонисты
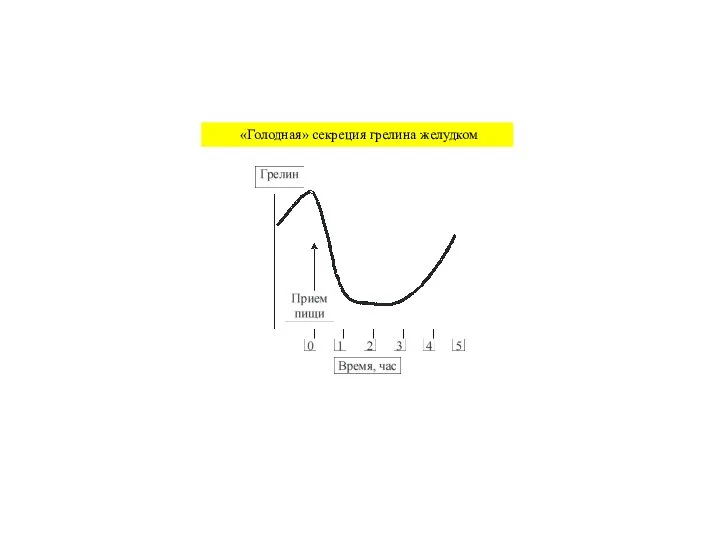
Слайд 44 «Голодная» секреция грелина желудком
«Голодная» секреция грелина желудком
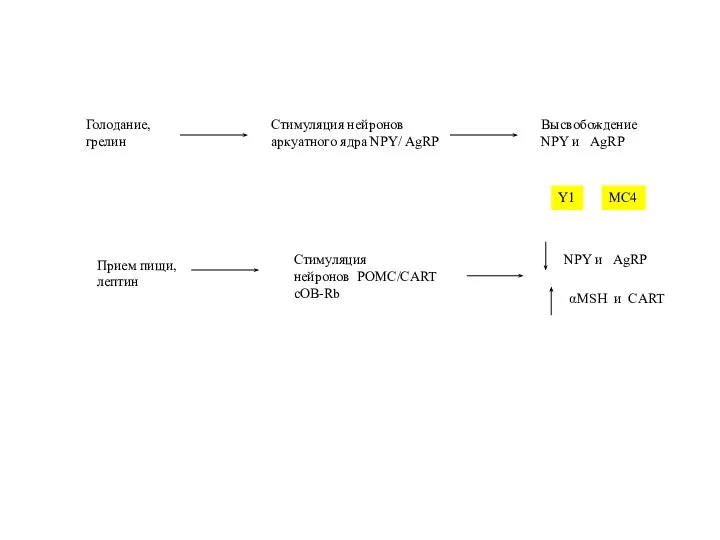
Слайд 45Голодание,
грелин
Стимуляция нейронов
аркуатного ядра NPY/ AgRP
Высвобождение
NPY и AgRP
Прием пищи,
лептин
Стимуляция
нейронов POMC/CART
сOB-Rb
NPY и AgRP
αМSH и
Голодание,
грелин
Стимуляция нейронов
аркуатного ядра NPY/ AgRP
Высвобождение
NPY и AgRP
Прием пищи,
лептин
Стимуляция
нейронов POMC/CART
сOB-Rb
NPY и AgRP
αМSH и
Слайд 46Frontiers in Systems Neuroscience, 2015, v 9, Article 150
Frontiers in Systems Neuroscience, 2015, v 9, Article 150
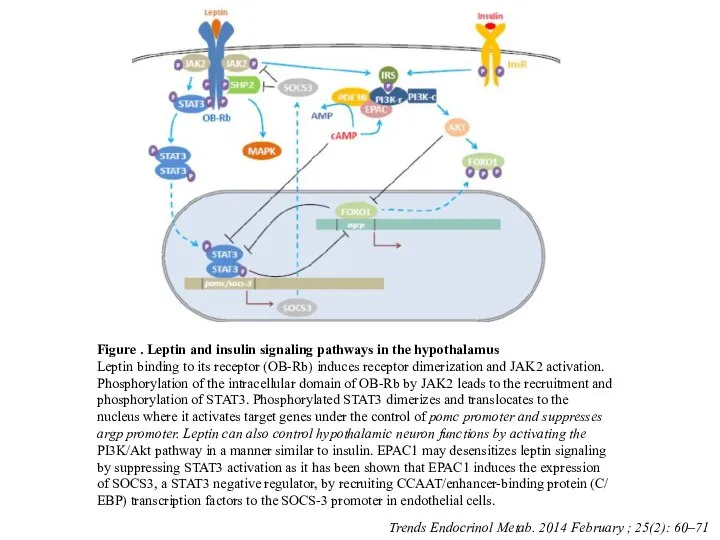
Слайд 47Figure . Leptin and insulin signaling pathways in the hypothalamus
Leptin binding to
Figure . Leptin and insulin signaling pathways in the hypothalamus
Leptin binding to
Слайд 48MC3 – R
MC4 - R
Агонист
αМСГ
Антагонист
белок агути,
AgrP
ПОМК
лептин
КРГ,урокортин
опиоиды
подавление
потребления
пищи
стимуляция
потребления
пищи
предшественник грелина
обестатин
грелин
NPY
GAL
μ-опиатные
MC3 – R
MC4 - R
Агонист
αМСГ
Антагонист
белок агути,
AgrP
ПОМК
лептин
КРГ,урокортин
опиоиды
подавление
потребления
пищи
стимуляция
потребления
пищи
предшественник грелина
обестатин
грелин
NPY
GAL
μ-опиатные
Слайд 49Ожирение
Ожирение

Слайд 50Снижение потребления пищи
(особенно жирных кислот)
Снижение секреции лептина
(снижение анорексигенного, энергозатратного
действия)
Увеличение секреции инсулина
(увеличение
Снижение потребления пищи
(особенно жирных кислот)
Снижение секреции лептина
(снижение анорексигенного, энергозатратного
действия)
Увеличение секреции инсулина
(увеличение
Слайд 51Детское ожирение
Увеличение количества адипоцитов в результате усиленной
дифференцировки преадипоцитов в адипоциты под действием
PPARγ,
Детское ожирение
Увеличение количества адипоцитов в результате усиленной
дифференцировки преадипоцитов в адипоциты под действием
PPARγ,

 Вегетативное размножение (стеблевое)
Вегетативное размножение (стеблевое) Гигиена сердечно-сосудистой системы
Гигиена сердечно-сосудистой системы Фотосинтез
Фотосинтез Биология - наука о живой природе
Биология - наука о живой природе Антропогенез. Стадии антропогенеза
Антропогенез. Стадии антропогенеза Подготовка к охоте и охота древних людей
Подготовка к охоте и охота древних людей Как тебе живется друг. Ахатинские улитки
Как тебе живется друг. Ахатинские улитки Панда
Панда Путешествие в страну Витаминию
Путешествие в страну Витаминию Животный мир тайги
Животный мир тайги Наймиліші собаки світу
Наймиліші собаки світу Свойства ферментов
Свойства ферментов Лиственные и хвойные породы деревьев
Лиственные и хвойные породы деревьев Отдел Покрытосеменные растения
Отдел Покрытосеменные растения Динофитовые водоросли
Динофитовые водоросли Влияние факторов среды на строение листа. Видоизменения листьев
Влияние факторов среды на строение листа. Видоизменения листьев Презентация на тему Прибрежно водные растения
Презентация на тему Прибрежно водные растения  Особливості селекції гладіолусів
Особливості селекції гладіолусів Обмен веществ и превращение энергии в клетке. Пластический обмен
Обмен веществ и превращение энергии в клетке. Пластический обмен Деление клетки. Митоз
Деление клетки. Митоз Основные закономерности наследования. Множественный аллелизм
Основные закономерности наследования. Множественный аллелизм Роль витаминов в жизни человека
Роль витаминов в жизни человека Презентация на тему Что у Земли внутри
Презентация на тему Что у Земли внутри  Обмен веществ и превращение энергии в клетке. 9 класс
Обмен веществ и превращение энергии в клетке. 9 класс Skeleton
Skeleton Западносибирский речной бобр
Западносибирский речной бобр Выделение. Выделительная система животных
Выделение. Выделительная система животных Разнообразие животных
Разнообразие животных