- Обмен липидов Катаболизм липидов

Содержание
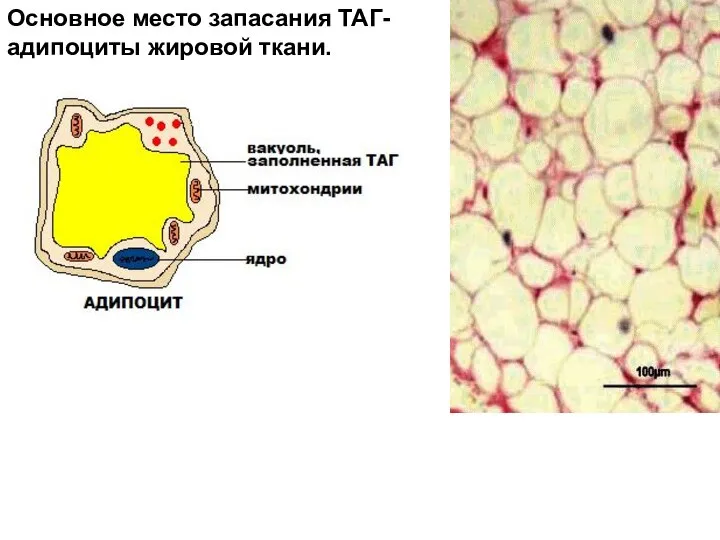
- 2. Основное место запасания ТАГ- адипоциты жировой ткани.
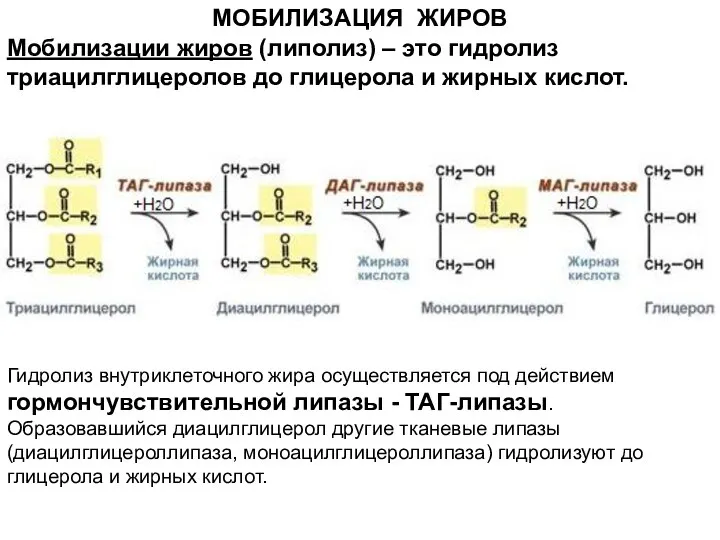
- 3. МОБИЛИЗАЦИЯ ЖИРОВ Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы. Образовавшийся диацилглицерол другие тканевые
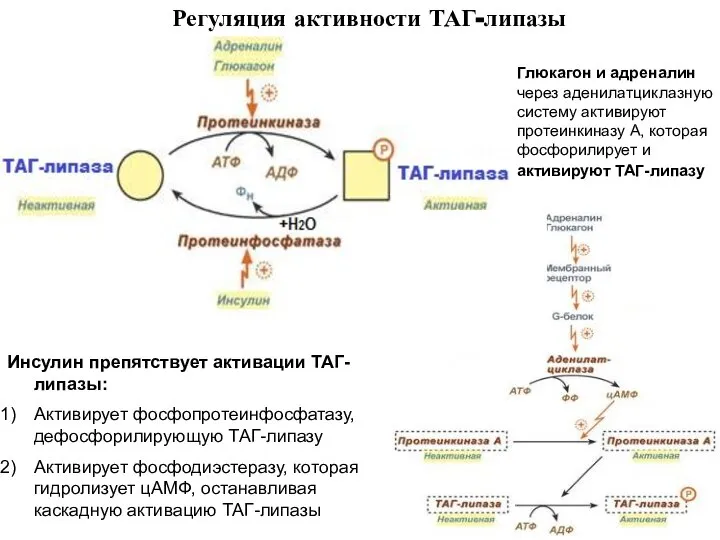
- 4. Регуляция активности ТАГ-липазы Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А, которая фосфорилирует и активируют
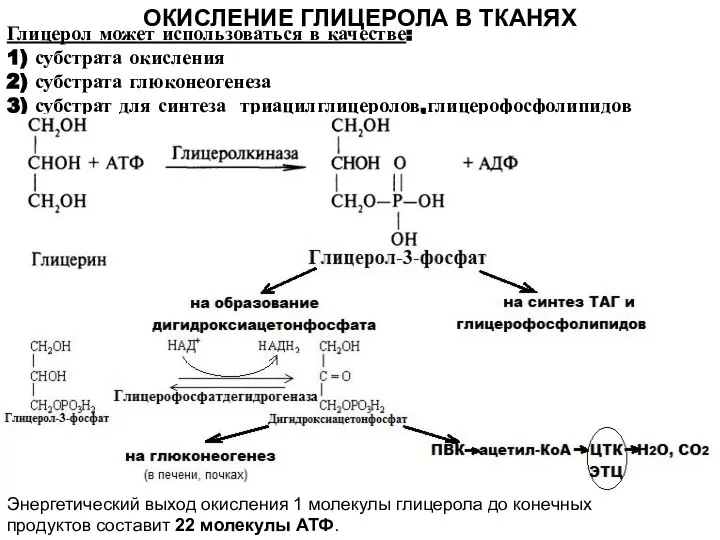
- 5. ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ Глицерол может использоваться в качестве: 1) субстрата окисления 2) субстрата глюконеогенеза 3)
- 6. Обозначение атомов углерода в жирной кислоте β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ β-Окисление - специфический путь катаболизма жирных
- 7. АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
- 8. ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ Карнитинацилтрансфераза I- регуляторный фермент β-окисления. + АДФ, АМФ, ацил-КоА -
- 9. РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
- 10. РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ: - молекула ацетил-КоА ФАДН2 НАДН2 укороченный на 2
- 11. ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ 1 виток β-окисления жирных кислот дает: 1 НАДН2→3 АТФ 1
- 12. ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
- 13. РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ - АТФ/АДФ - НАД+/НАДН2 наличие субстрата- жирных кислот, поступающих в
- 14. КЕТОНОВЫЕ ТЕЛА Содержание кетоновых тел в сыворотке крови человека в норме 0,03- 0,6 мМ/л Концентрация кетоновых
- 15. БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)
- 16. синтез ГМГ-КоА-синтазы увеличивается при повышении концентрации жирных кислот в крови (голодание, физическая работа) - ГМГ-КоА-синтаза ингибируется
- 17. ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА -сахарный диабет, -длительное голодание, -несбалансированное питание (нарушение соотношения углеводов и липидов пищи), -продолжительная
- 18. КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)
- 20. Скачать презентацию
Слайд 3МОБИЛИЗАЦИЯ ЖИРОВ
Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы. Образовавшийся
МОБИЛИЗАЦИЯ ЖИРОВ
Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы. Образовавшийся
Слайд 4Регуляция активности ТАГ-липазы
Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А, которая
Регуляция активности ТАГ-липазы
Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А, которая
Слайд 5ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ
Глицерол может использоваться в качестве:
1) субстрата окисления
2) субстрата
ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ
Глицерол может использоваться в качестве:
1) субстрата окисления
2) субстрата
Слайд 6Обозначение атомов углерода в жирной кислоте
β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
β-Окисление - специфический путь
Обозначение атомов углерода в жирной кислоте
β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
β-Окисление - специфический путь

Слайд 7АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
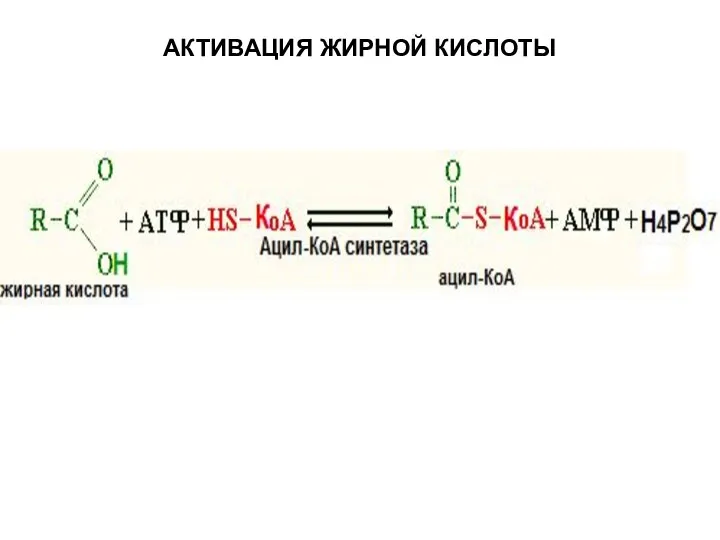
Слайд 8ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ
Карнитинацилтрансфераза I- регуляторный фермент β-окисления.
+ АДФ,
ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ
Карнитинацилтрансфераза I- регуляторный фермент β-окисления.
+ АДФ,
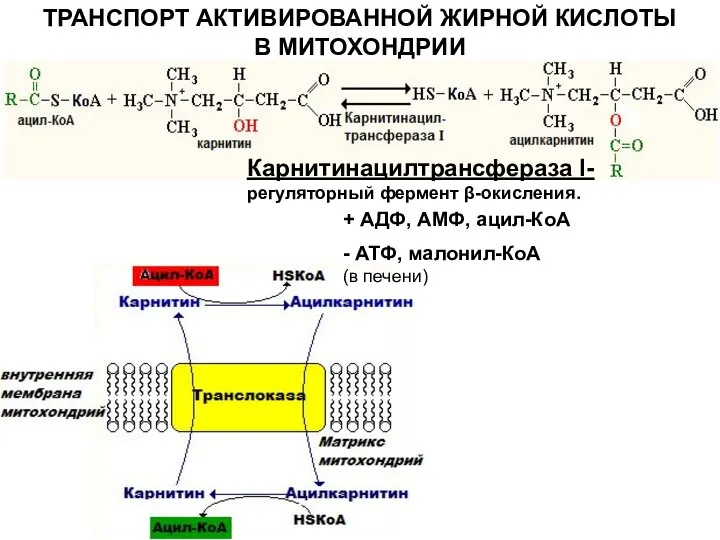
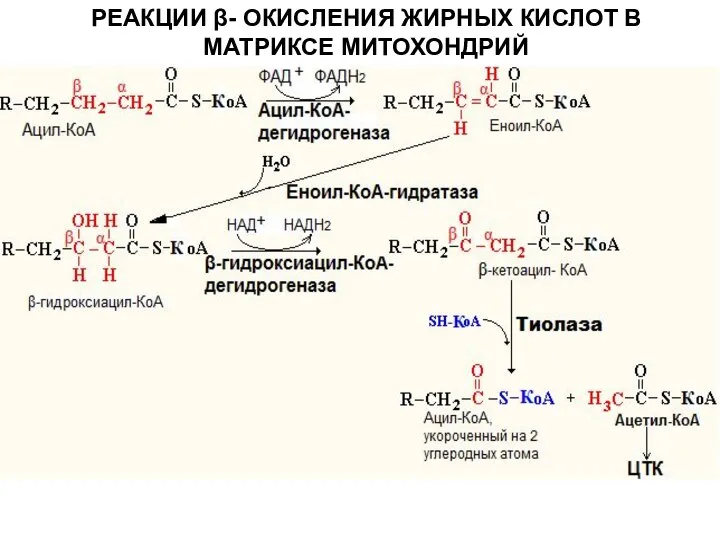
Слайд 9РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
Слайд 10РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ:
- молекула ацетил-КоА
ФАДН2
НАДН2
укороченный на 2
РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ:
- молекула ацетил-КоА
ФАДН2
НАДН2
укороченный на 2
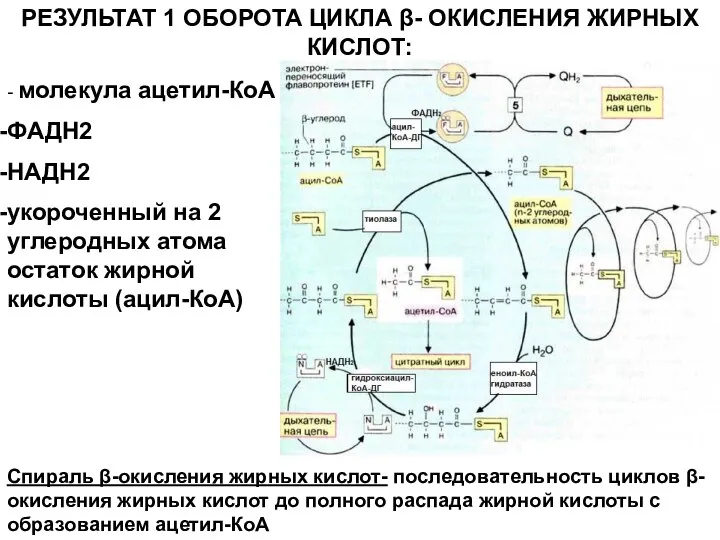
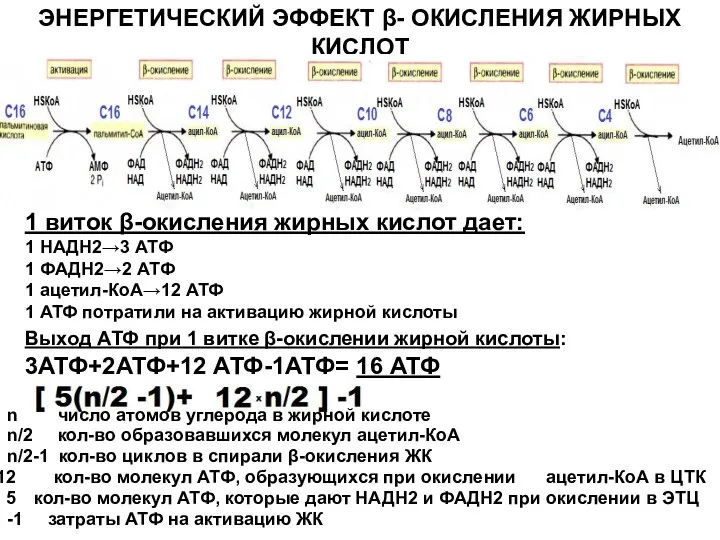
Слайд 11ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
1 виток β-окисления жирных кислот дает:
1 НАДН2→3
ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
1 виток β-окисления жирных кислот дает:
1 НАДН2→3
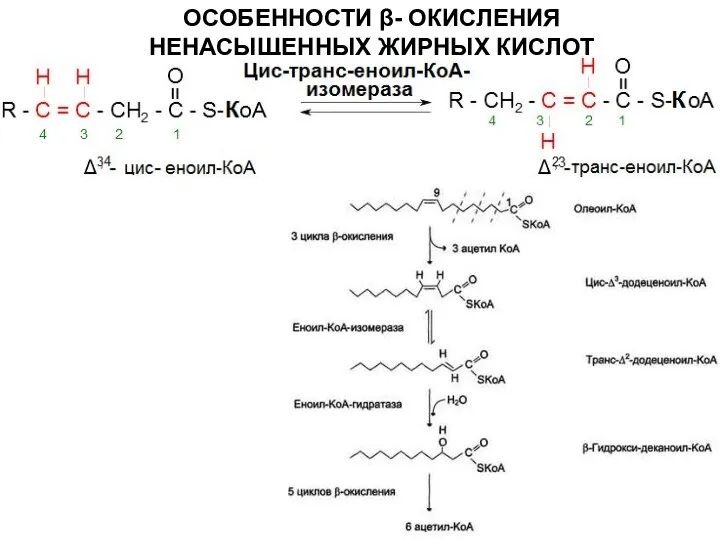
Слайд 12ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
Слайд 13РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
- АТФ/АДФ
- НАД+/НАДН2
наличие субстрата- жирных кислот, поступающих
РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
- АТФ/АДФ
- НАД+/НАДН2
наличие субстрата- жирных кислот, поступающих

Слайд 14КЕТОНОВЫЕ ТЕЛА
Содержание кетоновых тел в сыворотке крови человека в норме 0,03- 0,6
КЕТОНОВЫЕ ТЕЛА
Содержание кетоновых тел в сыворотке крови человека в норме 0,03- 0,6

Слайд 15БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)
БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)
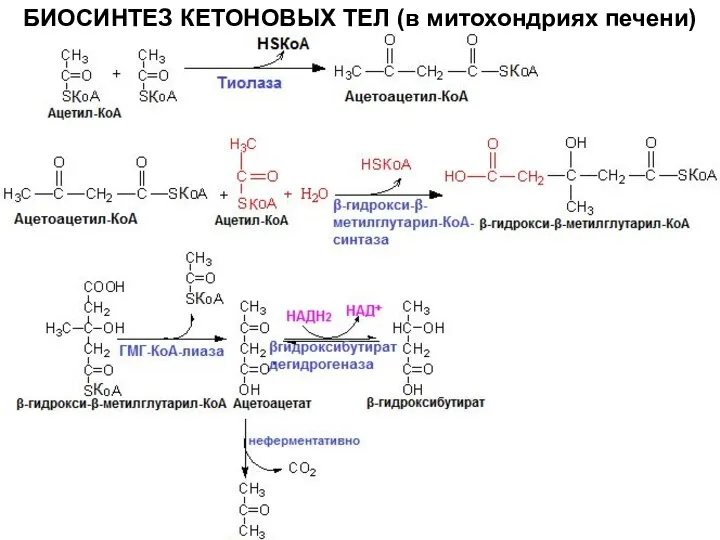
Слайд 16синтез ГМГ-КоА-синтазы увеличивается при повышении концентрации жирных кислот в крови (голодание, физическая

Слайд 17ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА
-сахарный диабет,
-длительное голодание,
-несбалансированное питание (нарушение соотношения углеводов и липидов пищи),
-продолжительная
ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА
-сахарный диабет,
-длительное голодание,
-несбалансированное питание (нарушение соотношения углеводов и липидов пищи),
-продолжительная

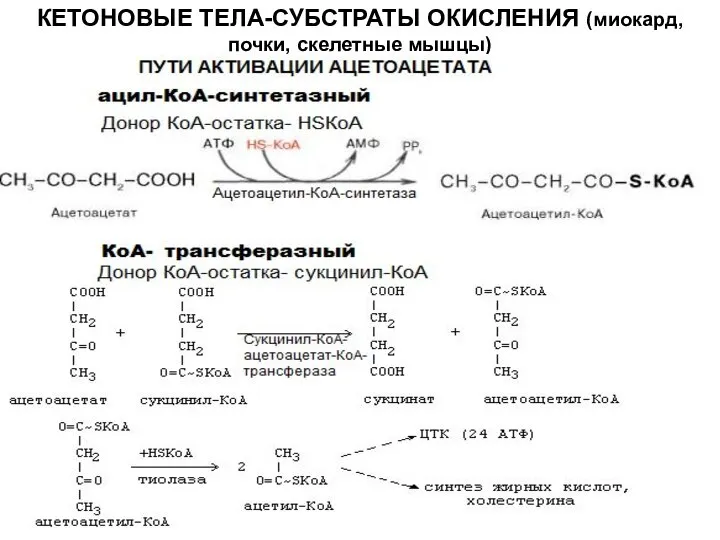
Слайд 18КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)
КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)
 304890b5af2e4dd89e9358e66fda3faa
304890b5af2e4dd89e9358e66fda3faa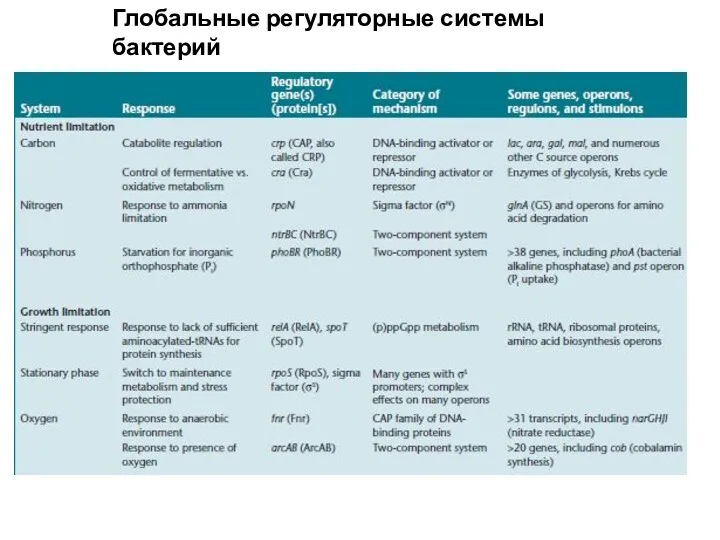 Глобальные регуляторные системы бактерий
Глобальные регуляторные системы бактерий Царство растений
Царство растений Өзектілігі
Өзектілігі Криптиды. Криптозоология
Криптиды. Криптозоология Отряды млекопитающих
Отряды млекопитающих Основы Физиологии труда. Труд умственный и труд физический
Основы Физиологии труда. Труд умственный и труд физический Экологическая капель. Интеллектуальная игра для учащихся 6-9 классов
Экологическая капель. Интеллектуальная игра для учащихся 6-9 классов будова клітини 27
будова клітини 27 Презентация на тему "Генетический тест" - презентации по Биологии
Презентация на тему "Генетический тест" - презентации по Биологии Презентация на тему Уход за кожей. Болезни кожи
Презентация на тему Уход за кожей. Болезни кожи  Регуляция дыхания. ФУС поддержания постоянства газов в крови
Регуляция дыхания. ФУС поддержания постоянства газов в крови Презентация на тему Популяция – единица эволюции
Презентация на тему Популяция – единица эволюции  Витамины и здоровье человека
Витамины и здоровье человека Вирусы
Вирусы Естественные методы защиты растений
Естественные методы защиты растений Презентация на тему Разнообразие животного и растительного мира
Презентация на тему Разнообразие животного и растительного мира  Онтогенез
Онтогенез Биологический мониторинг состояния растительного и животного мира Бурзянского района
Биологический мониторинг состояния растительного и животного мира Бурзянского района Переваривание и всасывание в тонком и толстом кишечнике. Роль поджелудочной железы и печени
Переваривание и всасывание в тонком и толстом кишечнике. Роль поджелудочной железы и печени Биологические приборы и инструменты
Биологические приборы и инструменты Культурные растения. 6 класс
Культурные растения. 6 класс Рекрутування нейромоторних одиниць та аеробно-анаеробний перехід
Рекрутування нейромоторних одиниць та аеробно-анаеробний перехід Сельдеобразные, отряд лучепёрых рыб
Сельдеобразные, отряд лучепёрых рыб Улитка - чудо природы
Улитка - чудо природы Происхождение жизни на Земле 5 класс - Презентация_
Происхождение жизни на Земле 5 класс - Презентация_ Мутации. Системы репарации. Мутационная и комбинативная изменчивость прокариот и эукариот
Мутации. Системы репарации. Мутационная и комбинативная изменчивость прокариот и эукариот Покормите птиц!
Покормите птиц!