- Митохондрии грибов

Содержание
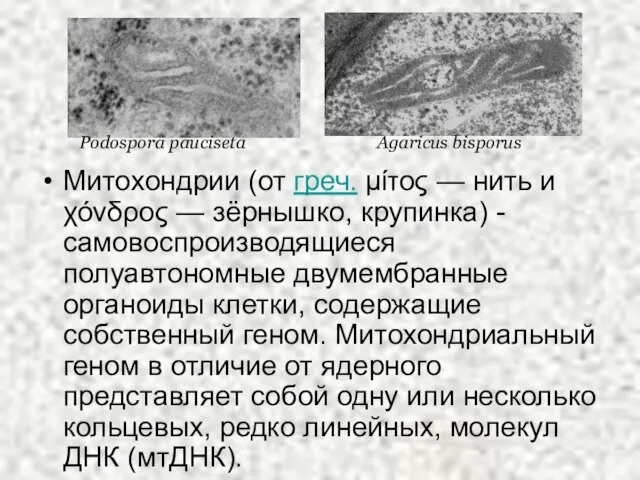
- 2. Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся полуавтономные двумембранные органоиды
- 3. Митохондрии характерны за малым исключением для всех эукариотических клеток, как аутотрофных (фотосинтезирующие растения), так и гетеротрофных
- 4. Митохондрии это мультифункциональные полиморфные органеллы, выполняющие функции обеспечения жизнедеятельности клетки. Динамика изменения морфологии митохондрий позволяет адаптировать
- 5. Функции митохондрий Основная функция митохондрий связана с окислением органических соединений и использованием освобождающейся при распаде этих
- 6. Биосинтез эргостерола у дрожжей Биосинтез эргостерола у дрожжей
- 7. Субкомпартменты внутренней мембраны митохондрий Митохондрии содержат около 800 белков/1500 у человека Субкомпартментализация внутренней мембраны митохондрий. Распространение
- 8. Комплекс I (НАДН дегидрогеназаКомплекс I (НАДН дегидрогеназа) окисляет НАД∙НКомплекс I (НАДН дегидрогеназа) окисляет НАД∙Н, отбирая у
- 9. Гипотетическая модель гибридной дыхательной системы у Fusarium oxysporum. Nar – нитратредуктаза Nir - нитритредуктаза
- 10. Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к 1911 году, несколько позже описаны
- 11. Методы наблюдения митохондрий у грибов Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды в геном митохондрий) Методы
- 12. Методы иммуноцитохимии (ТЭМ)
- 13. Морфология и распределение хондриома в клетке определяется следующими факторами: - положением клеток в колонии - условиями
- 14. Локализованные в апикальном кончике мицелия митохондрии (до 50 мкм) у Neurospora crassa возможно участвуют в устранении
- 15. Распределение и морфология митохондрий у мицелиальных грибов
- 16. Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006) . Тип 1. Характерен для:
- 17. Базидии шампиньона двуспорового
- 18. конидии мицелий
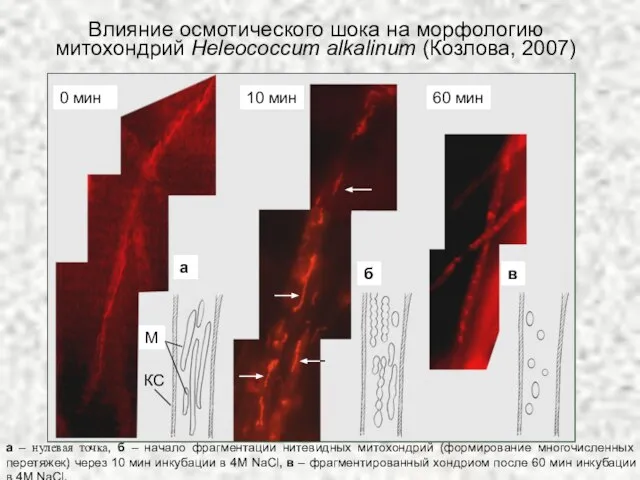
- 19. Влияние стрессового фактора на морфологию (фрагментацию) митохондрий Podospora pauciseta Родамин 6Ж GFP плазмида контроль опыт
- 20. а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование многочисленных перетяжек) через 10 мин
- 21. Распределение и морфология митохондрий у одноклеточных грибов
- 22. Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae
- 23. Miyakawa et al., 1984 Деление и слияние митохондрий в процессе клеточного цикла у дрожжей
- 25. Размеры митохондрий Размеры митохондрий непостоянны у разных видов. Обычно митохондрии представляют собой мелкие (длина 0.5-3 мкм,
- 26. Ультраструктура митохондрий в клетках поверхностно растущего вегетативного мицелия (А) и субгимения (Б) Agaricus bisporus А Б
- 28. Морфология крист у дрожжей Внутренняя митохондриальная мембрана способна в зависимости от напряженности энергетического обмена образовывать большее
- 30. Геном митохондрий грибов ДНК в митохондриях представлена циклическими молекулами, не образующими связь с гистонными белками. Митохондриальная
- 31. Наследование митохондрий Сохранение целостности митохондриального генома в процессе наследования очень важно для дыхательной функции. Факторы, контролирующие
- 32. Движение митохондрий В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans в транспорте митохондрий большую
- 33. Связь митохондрий с другими органеллами: ядра и пероксисомы (клетки мицелия Agaricus bisporus, связь митохондрий и пероксисомы)
- 34. Слияние (англ. fusion) и деление (англ.fission или division) митохондрий Морфология митохондрий и количество копий зависит от
- 35. Динамика митохондрий и ее роль в клетке Цитокинез Развитие Апоптоз Передача энергии – для метаболически активных
- 36. Динамика изменения плотности митохондриальных профилей в зависимости от продолжительности культивирования P. anserina в поверхностных и в
- 37. Westermann, 2008 Westermann, 2008
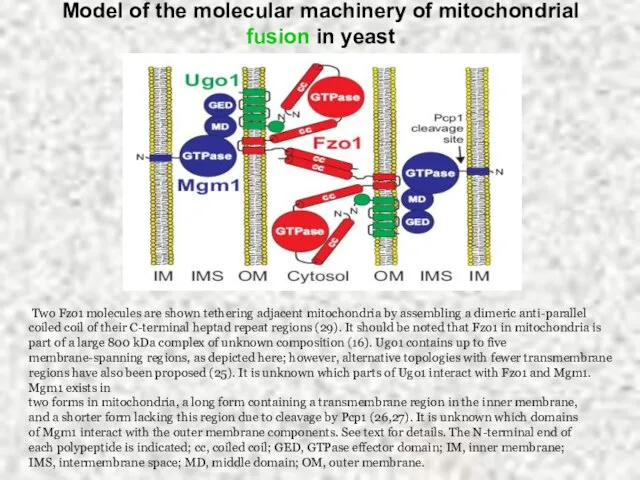
- 38. Model of the molecular machinery of mitochondrial fusion in yeast Two Fzo1 molecules are shown tethering
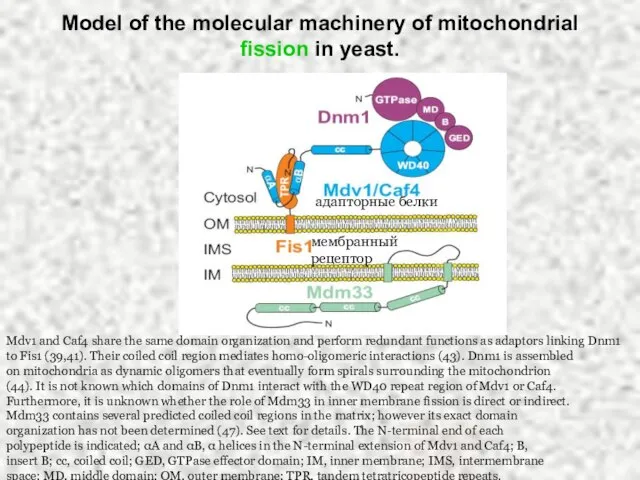
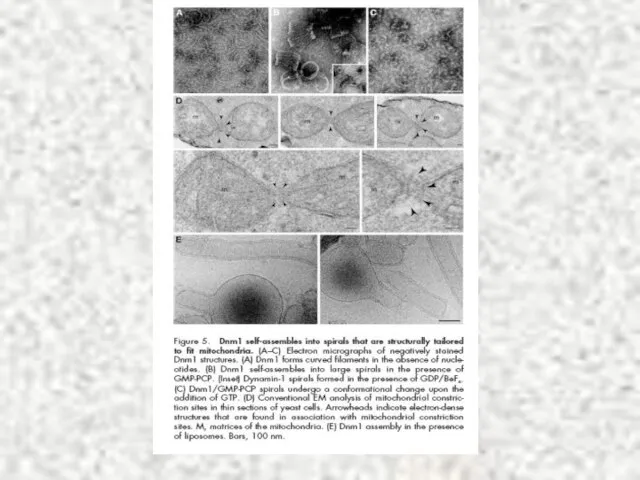
- 39. Model of the molecular machinery of mitochondrial fission in yeast. Mdv1 and Caf4 share the same
- 42. Продолжительность жизни, старение и смерть грибов
- 43. Долгоживущий мицелий базидиомицетов
- 44. Armillaria bulbosa (Smith , M., J. Bruhn and J. Anderson, 1992. The fungus Armillaria bulbosa is
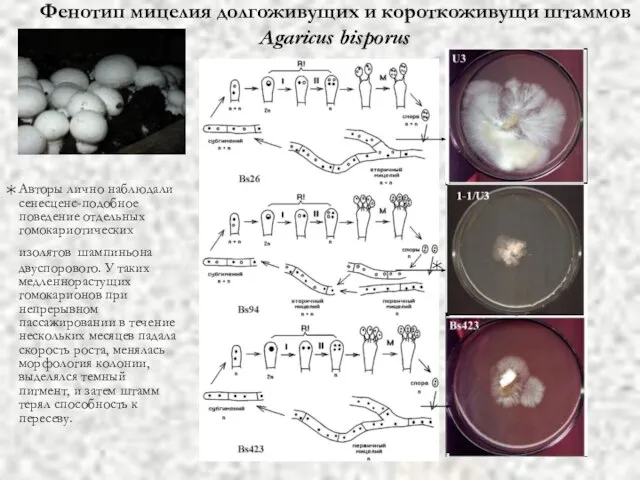
- 45. Фенотип мицелия долгоживущих и короткоживущи штаммов Agaricus bisporus Авторы лично наблюдали сенесценс-подобное поведение отдельных гомокариотических изолятов
- 46. Podospora anserina жизненный цикл природных штаммов составляет в среднем 25 суток senescent phenotype
- 47. Продолжительность жизни Saccharomyces cerevisiae Средняя продолжительность жизни клетки лабораторных штаммов дрожжей Saccharomyces cerevisiae составляет в среднем
- 48. Какие факторы влияют на продолжительность жизни модельных объектов Podospora anserina и Saccharomyces cerevisiae ? Источник и
- 49. На продолжительность жизни оказывают влияние не только факторы окружающей среды и доступность питательных веществ, но и
- 50. Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех биологических систем и может быть определен
- 51. Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992, Osiewacz and Hamman, 2006), которые
- 52. У грибов процесс старения включает несколько возможных базисных механизмов (Мажейка и сотр., 2011): хронологическое старение репликативное
- 53. Хронологическое и репликативное (митотическое) старение Хронологическое старение - потеря жизнеспособности индивидуальных клеток дрожжей при культивировании в
- 54. Накопление факторов старения у дрожжей Модельная схема накопления и распределения между клетками условного фактора старения у
- 55. Накопление факторов старения у Рodospora anserina Модельная схема накопления и распределения между клеток условного фактора старения.
- 56. Митохондриальные факторы старения P. anserina Схема вырезания и накапливания последовательности α-sen в стареющих культурах P. anserina
- 57. Какие факторы влияют на продление или сокращение продолжительности жизни у грибов? Рассмотрим один фактор - условия
- 58. Культивирование мицелия Podospora anserina в глубинных условиях (Turker, Cummings, 1987; Кудрявцева с сотр., 2011) Культивирование в
- 59. Культивирование мицелия Agaricus bisporus в глубинных условиях Культивирование в жидкой среде вызывает гибель клеток мицелия через
- 60. Роль митохондрий в апоптозе клетки
- 61. Apoptosis pathways in fungal growth, development and ageing Hamman et al., 2008
- 62. Основная молекулярная машина апоптоза у дрожжей Saccharomyces cerevisiae
- 63. Дрожжи Метакаспаза (Yca1p) Uth1p Dnm1p Fis1 Компоненты, участвующие в программируемой клеточной смерти у дрожжей STM1 AIF?
- 64. Феромон Увеличение концентрации цитоплазматического Ca2+ Увеличение ΔΨ в митохондриях Образование АФК Потеря целостности митохондрий Гибель клеток
- 65. Морфология митохондрий клеток глубинного мицелия Agaricus bisporus
- 66. Ультраструктура клетки глубинного мицелия Agaricus bisporus после длительного культивирования
- 68. Скачать презентацию
Слайд 2Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся полуавтономные
Митохондрии (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) - самовоспроизводящиеся полуавтономные
Слайд 3Митохондрии характерны за малым исключением для всех эукариотических клеток, как аутотрофных (фотосинтезирующие
Митохондрии характерны за малым исключением для всех эукариотических клеток, как аутотрофных (фотосинтезирующие

Слайд 4Митохондрии это мультифункциональные полиморфные органеллы, выполняющие функции обеспечения жизнедеятельности клетки.
Динамика изменения
Митохондрии это мультифункциональные полиморфные органеллы, выполняющие функции обеспечения жизнедеятельности клетки.
Динамика изменения

Слайд 5Функции митохондрий
Основная функция митохондрий связана с окислением органических соединений и использованием освобождающейся
Функции митохондрий
Основная функция митохондрий связана с окислением органических соединений и использованием освобождающейся

Слайд 6Биосинтез эргостерола у дрожжей
Биосинтез эргостерола у дрожжей
Биосинтез эргостерола у дрожжей
Биосинтез эргостерола у дрожжей

Слайд 7Субкомпартменты внутренней мембраны митохондрий
Митохондрии содержат около 800 белков/1500 у человека
Субкомпартментализация внутренней мембраны
Субкомпартменты внутренней мембраны митохондрий
Митохондрии содержат около 800 белков/1500 у человека
Субкомпартментализация внутренней мембраны

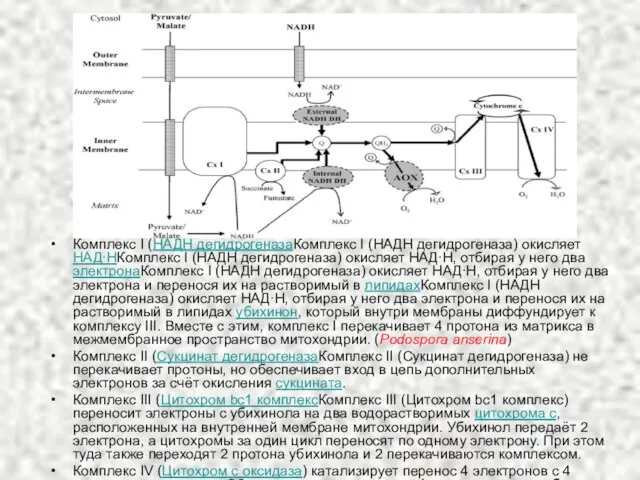
Слайд 8Комплекс I (НАДН дегидрогеназаКомплекс I (НАДН дегидрогеназа) окисляет НАД∙НКомплекс I (НАДН дегидрогеназа)
Комплекс I (НАДН дегидрогеназаКомплекс I (НАДН дегидрогеназа) окисляет НАД∙НКомплекс I (НАДН дегидрогеназа)
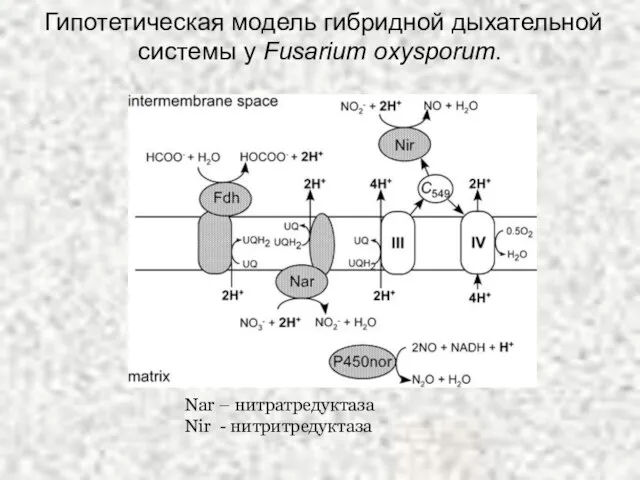
Слайд 9Гипотетическая модель гибридной дыхательной системы у Fusarium oxysporum.
Nar – нитратредуктаза
Nir - нитритредуктаза
Гипотетическая модель гибридной дыхательной системы у Fusarium oxysporum.
Nar – нитратредуктаза Nir - нитритредуктаза
Слайд 10Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к 1911
Первые наблюдения митохондрий у гриба Pustularia versispora (Tarzetta catinus) относятся к 1911

Слайд 11Методы наблюдения митохондрий у грибов
Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды в
Методы наблюдения митохондрий у грибов
Флуоресцентные (потенциалзависимые красители или встраивание GFP плазмиды в

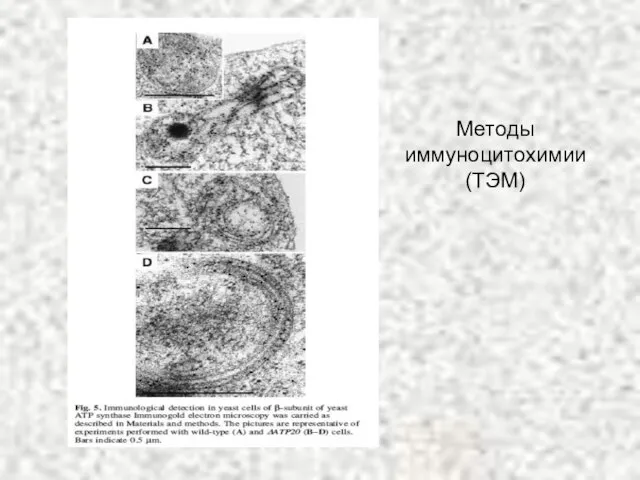
Слайд 12Методы иммуноцитохимии (ТЭМ)
Методы иммуноцитохимии (ТЭМ)
Слайд 13Морфология и распределение хондриома в клетке
определяется следующими факторами:
- положением клеток в
Морфология и распределение хондриома в клетке определяется следующими факторами: - положением клеток в

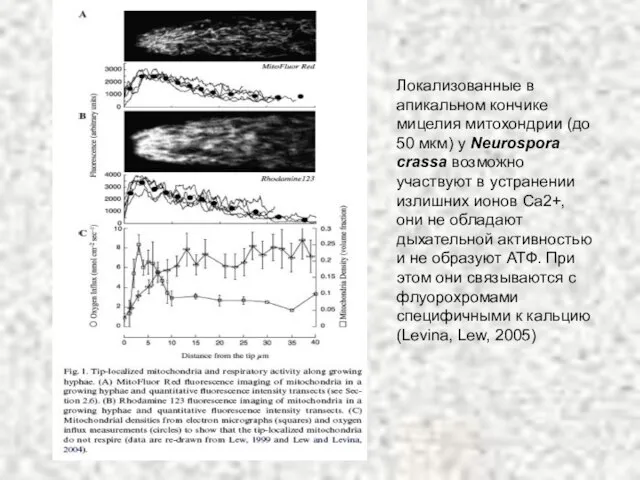
Слайд 14Локализованные в апикальном кончике мицелия митохондрии (до 50 мкм) у Neurospora crassa
Локализованные в апикальном кончике мицелия митохондрии (до 50 мкм) у Neurospora crassa
Слайд 15Распределение и морфология митохондрий у мицелиальных грибов
Распределение и морфология митохондрий у мицелиальных грибов

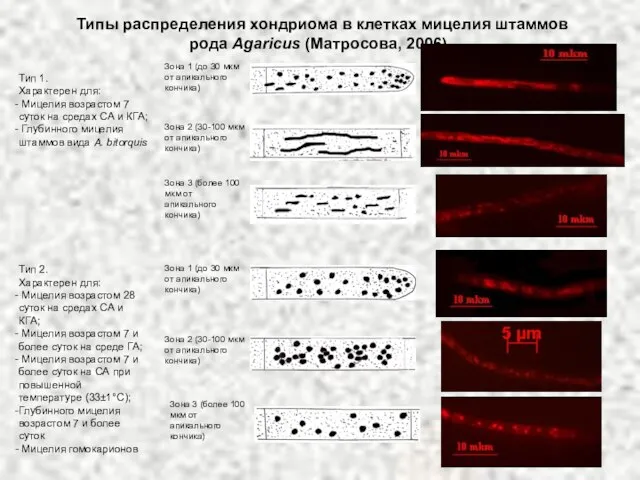
Слайд 16Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006) .
Тип
Типы распределения хондриома в клетках мицелия штаммов рода Agaricus (Матросова, 2006) .
Тип
Слайд 17Базидии шампиньона двуспорового
Базидии шампиньона двуспорового

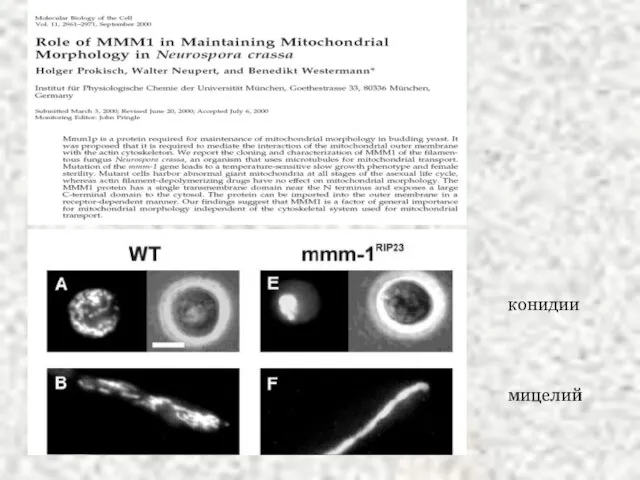
Слайд 18конидии
мицелий
конидии
мицелий
Слайд 19Влияние стрессового фактора на морфологию (фрагментацию) митохондрий Podospora pauciseta
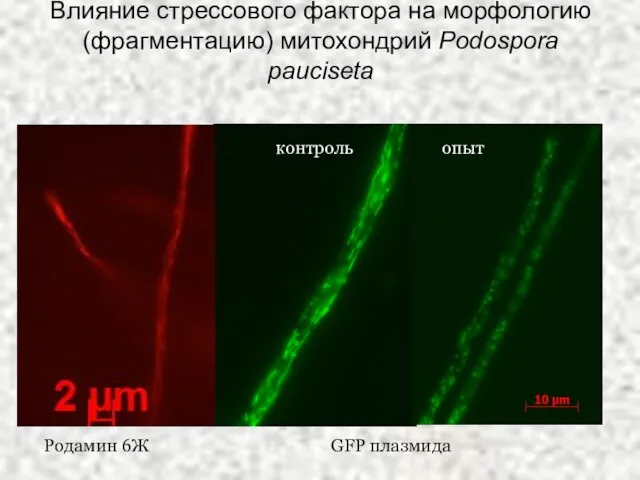
Родамин 6Ж
GFP плазмида
контроль
опыт
Влияние стрессового фактора на морфологию (фрагментацию) митохондрий Podospora pauciseta
Родамин 6Ж
GFP плазмида
контроль
опыт
Слайд 20а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование многочисленных
а – нулевая точка, б – начало фрагментации нитевидных митохондрий (формирование многочисленных
Слайд 21Распределение и морфология митохондрий у одноклеточных грибов
Распределение и морфология митохондрий у одноклеточных грибов

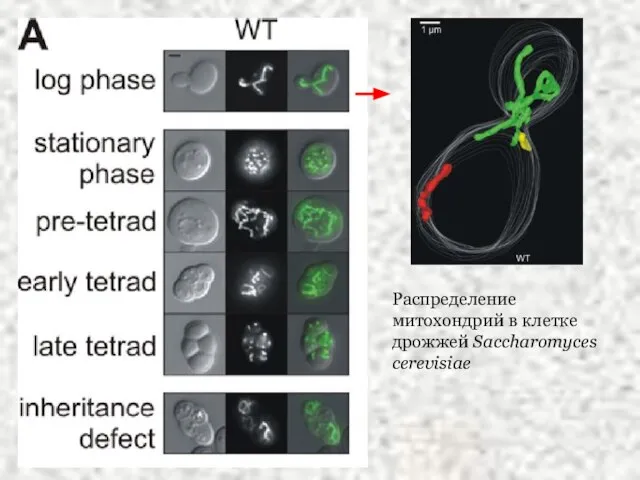
Слайд 22Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae
Распределение митохондрий в клетке дрожжей Saccharomyces cerevisiae
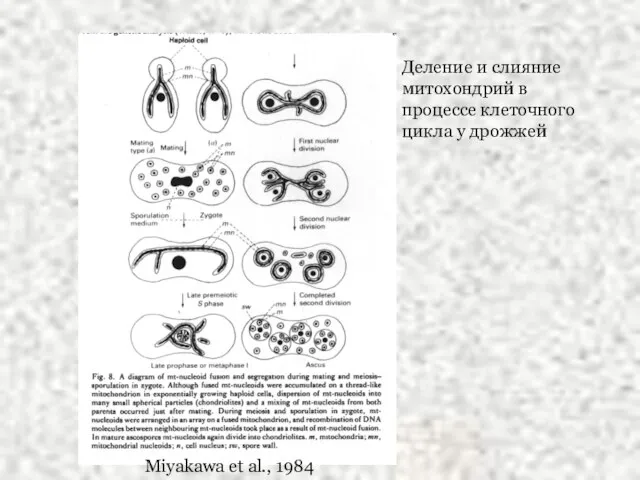
Слайд 23Miyakawa et al., 1984
Деление и слияние митохондрий в процессе клеточного цикла у
Miyakawa et al., 1984
Деление и слияние митохондрий в процессе клеточного цикла у

Слайд 25Размеры митохондрий
Размеры митохондрий непостоянны у разных видов. Обычно митохондрии представляют собой
Размеры митохондрий
Размеры митохондрий непостоянны у разных видов. Обычно митохондрии представляют собой

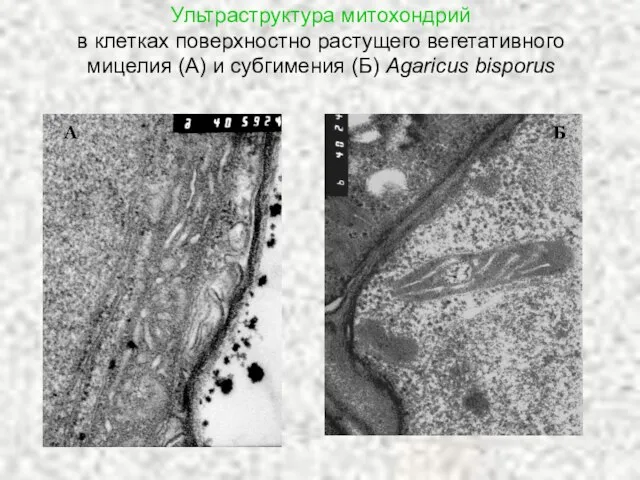
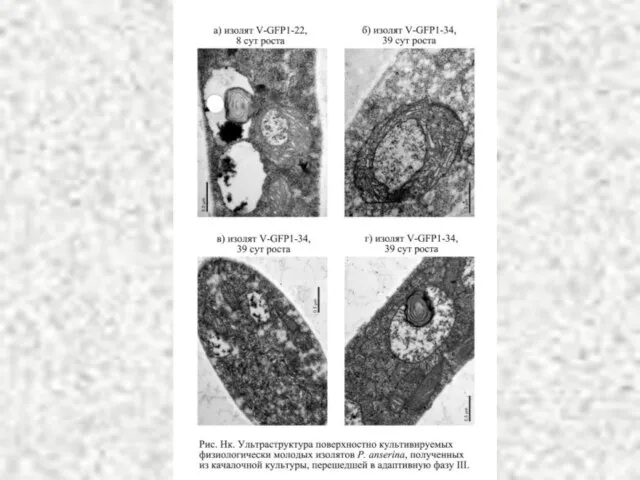
Слайд 26Ультраструктура митохондрий
в клетках поверхностно растущего вегетативного мицелия (А) и субгимения (Б)
Ультраструктура митохондрий в клетках поверхностно растущего вегетативного мицелия (А) и субгимения (Б)
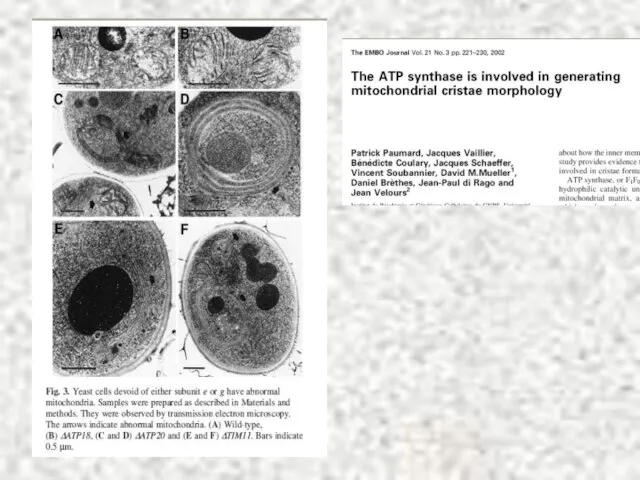
Слайд 28Морфология крист у дрожжей
Внутренняя митохондриальная мембрана способна в зависимости от напряженности
Морфология крист у дрожжей
Внутренняя митохондриальная мембрана способна в зависимости от напряженности

Слайд 30Геном митохондрий грибов
ДНК в митохондриях представлена циклическими молекулами, не образующими связь с
Геном митохондрий грибов
ДНК в митохондриях представлена циклическими молекулами, не образующими связь с

Слайд 31Наследование митохондрий
Сохранение целостности митохондриального генома в процессе наследования очень важно для дыхательной
Наследование митохондрий
Сохранение целостности митохондриального генома в процессе наследования очень важно для дыхательной

Слайд 32Движение митохондрий
В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans в
Движение митохондрий
В клетках Saccharomyces cerevisiae, как и в клетках Aspergillus nidulans в

Слайд 33Связь митохондрий с другими органеллами: ядра и пероксисомы
(клетки мицелия Agaricus bisporus,
Связь митохондрий с другими органеллами: ядра и пероксисомы (клетки мицелия Agaricus bisporus,

Слайд 34Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Морфология митохондрий и количество
Слияние (англ. fusion) и деление (англ.fission или division) митохондрий
Морфология митохондрий и количество

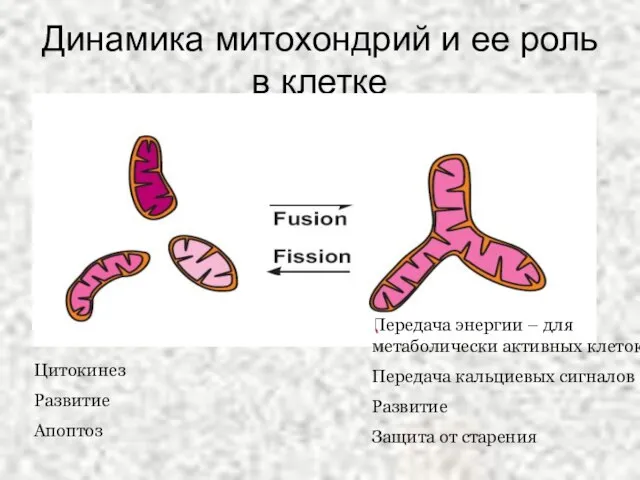
Слайд 35Динамика митохондрий и ее роль в клетке
Цитокинез
Развитие
Апоптоз
Передача энергии – для метаболически активных
Динамика митохондрий и ее роль в клетке
Цитокинез
Развитие
Апоптоз
Передача энергии – для метаболически активных
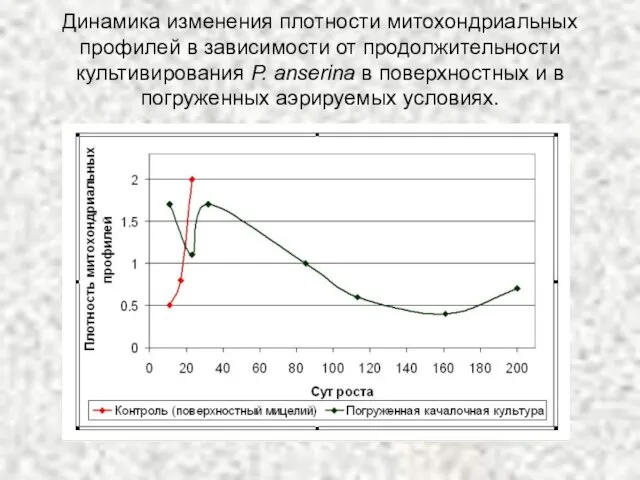
Слайд 36Динамика изменения плотности митохондриальных профилей в зависимости от продолжительности культивирования P. anserina
Динамика изменения плотности митохондриальных профилей в зависимости от продолжительности культивирования P. anserina
Слайд 37 Westermann, 2008
Westermann, 2008
Westermann, 2008
Westermann, 2008

Слайд 38Model of the molecular machinery of mitochondrial fusion in yeast
Two Fzo1
Model of the molecular machinery of mitochondrial fusion in yeast
Two Fzo1
Слайд 39Model of the molecular machinery of mitochondrial fission in yeast.
Mdv1 and Caf4
Model of the molecular machinery of mitochondrial fission in yeast.
Mdv1 and Caf4

Слайд 42Продолжительность жизни, старение и смерть грибов
Продолжительность жизни, старение и смерть грибов

Слайд 43Долгоживущий мицелий базидиомицетов
Долгоживущий мицелий базидиомицетов

Слайд 44Armillaria bulbosa (Smith , M., J. Bruhn and J. Anderson, 1992. The
Armillaria bulbosa (Smith , M., J. Bruhn and J. Anderson, 1992. The

Слайд 45Фенотип мицелия долгоживущих и короткоживущи штаммов Agaricus bisporus
Авторы лично наблюдали сенесценс-подобное поведение
Фенотип мицелия долгоживущих и короткоживущи штаммов Agaricus bisporus
Авторы лично наблюдали сенесценс-подобное поведение
Слайд 46Podospora anserina
жизненный цикл природных штаммов составляет
в среднем 25 суток
senescent phenotype
Podospora anserina
жизненный цикл природных штаммов составляет
в среднем 25 суток
senescent phenotype
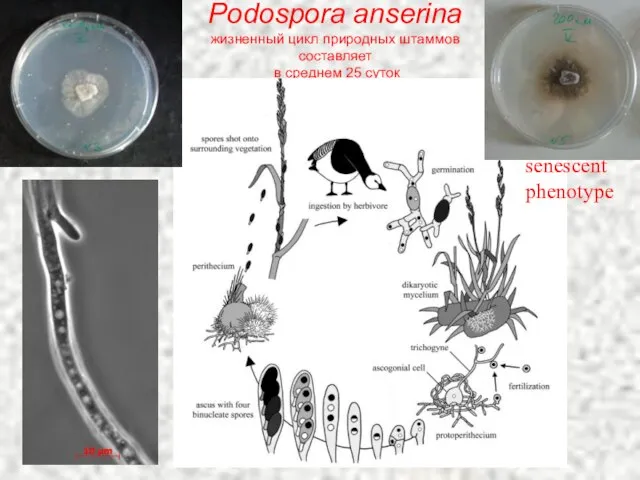
Слайд 47Продолжительность жизни Saccharomyces cerevisiae
Средняя продолжительность жизни клетки лабораторных штаммов дрожжей Saccharomyces
Продолжительность жизни Saccharomyces cerevisiae
Средняя продолжительность жизни клетки лабораторных штаммов дрожжей Saccharomyces

Слайд 48Какие факторы влияют на продолжительность жизни модельных объектов Podospora anserina и Saccharomyces
Какие факторы влияют на продолжительность жизни модельных объектов Podospora anserina и Saccharomyces

Слайд 49На продолжительность жизни оказывают влияние не только факторы окружающей среды и доступность
На продолжительность жизни оказывают влияние не только факторы окружающей среды и доступность

Слайд 50 Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех биологических
Старение (англ. аging) это фундаментальный процесс обнаруженный почти у всех биологических

Слайд 51Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992, Osiewacz
Признаки старения Podospora anserina and Neurospora crassa (senescent strains) (Griffiths, 1992, Osiewacz

Слайд 52У грибов процесс старения включает несколько возможных базисных механизмов (Мажейка и сотр.,
У грибов процесс старения включает несколько возможных базисных механизмов (Мажейка и сотр.,

Слайд 53Хронологическое и репликативное (митотическое) старение
Хронологическое старение - потеря жизнеспособности индивидуальных клеток дрожжей
Хронологическое и репликативное (митотическое) старение
Хронологическое старение - потеря жизнеспособности индивидуальных клеток дрожжей

Слайд 54Накопление факторов старения у дрожжей
Модельная схема накопления и распределения между клетками условного
Накопление факторов старения у дрожжей
Модельная схема накопления и распределения между клетками условного
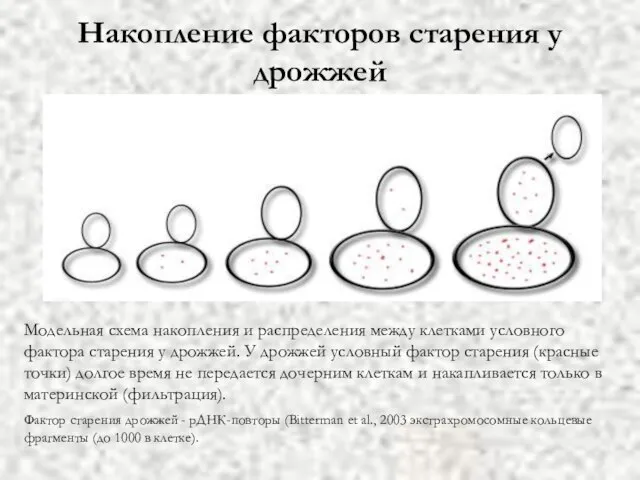
Слайд 55Накопление факторов старения у Рodospora anserina
Модельная схема накопления и распределения между клеток
Накопление факторов старения у Рodospora anserina
Модельная схема накопления и распределения между клеток

Слайд 56Митохондриальные факторы старения
P. anserina
Схема вырезания и накапливания последовательности α-sen
в стареющих
Митохондриальные факторы старения
P. anserina
Схема вырезания и накапливания последовательности α-sen
в стареющих
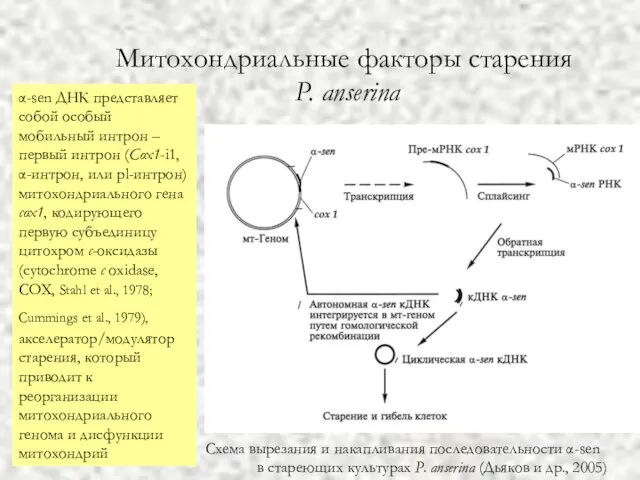
Слайд 57Какие факторы влияют на продление или сокращение продолжительности жизни у грибов?
Рассмотрим один
Какие факторы влияют на продление или сокращение продолжительности жизни у грибов?
Рассмотрим один

Слайд 58Культивирование мицелия Podospora anserina в глубинных условиях (Turker, Cummings, 1987; Кудрявцева с
Культивирование мицелия Podospora anserina в глубинных условиях (Turker, Cummings, 1987; Кудрявцева с

Слайд 59Культивирование мицелия Agaricus bisporus в глубинных условиях
Культивирование в жидкой среде вызывает гибель
Культивирование мицелия Agaricus bisporus в глубинных условиях
Культивирование в жидкой среде вызывает гибель

Слайд 60Роль митохондрий в апоптозе клетки
Роль митохондрий в апоптозе клетки

Слайд 61Apoptosis pathways in fungal growth,
development and ageing
Hamman et al., 2008
Apoptosis pathways in fungal growth,
development and ageing
Hamman et al., 2008
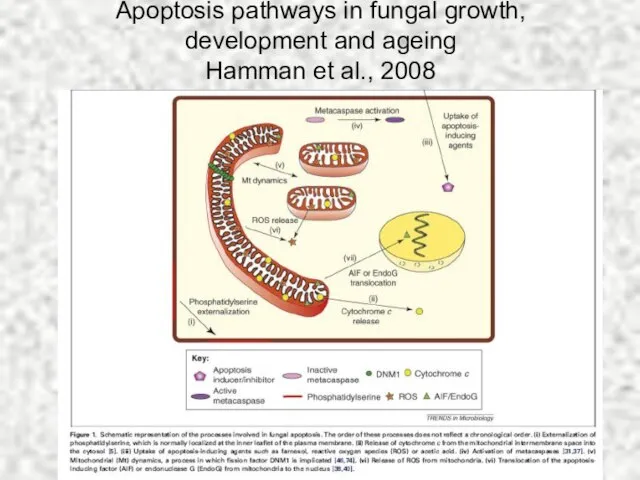
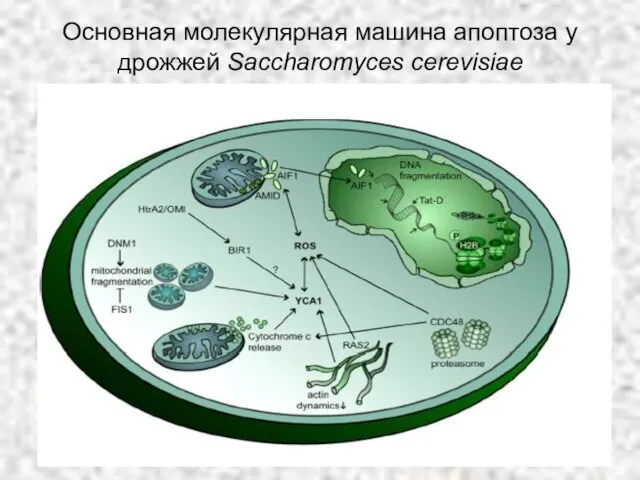
Слайд 62Основная молекулярная машина апоптоза у дрожжей Saccharomyces cerevisiae
Основная молекулярная машина апоптоза у дрожжей Saccharomyces cerevisiae
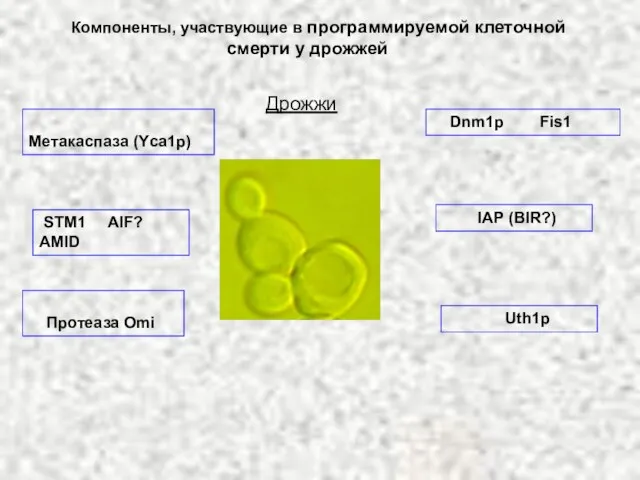
Слайд 63Дрожжи
Метакаспаза (Yca1p)
Uth1p
Dnm1p Fis1
Компоненты, участвующие в программируемой клеточной смерти у
Дрожжи
Метакаспаза (Yca1p)
Uth1p
Dnm1p Fis1
Компоненты, участвующие в программируемой клеточной смерти у
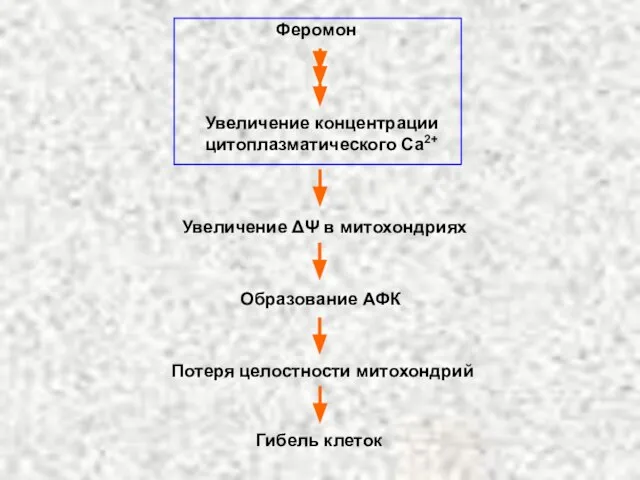
Слайд 64Феромон
Увеличение концентрации
цитоплазматического Ca2+
Увеличение ΔΨ в митохондриях
Образование АФК
Потеря целостности митохондрий
Гибель клеток
Феромон
Увеличение концентрации
цитоплазматического Ca2+
Увеличение ΔΨ в митохондриях
Образование АФК
Потеря целостности митохондрий
Гибель клеток
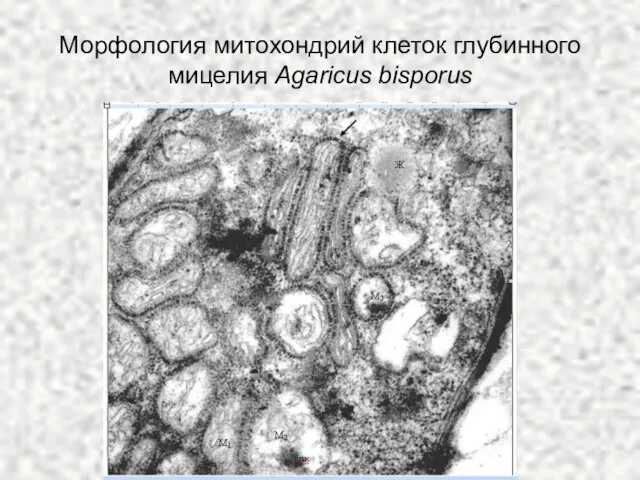
Слайд 65Морфология митохондрий клеток глубинного мицелия Agaricus bisporus
Морфология митохондрий клеток глубинного мицелия Agaricus bisporus
Слайд 66Ультраструктура клетки глубинного мицелия Agaricus bisporus после длительного культивирования
Ультраструктура клетки глубинного мицелия Agaricus bisporus после длительного культивирования

 Обеспечение гарантий прав граждан на накопительную часть трудовой пенсии
Обеспечение гарантий прав граждан на накопительную часть трудовой пенсии Герои Великой Отечественной Войны
Герои Великой Отечественной Войны Использование разнообразных форм и средств контроля за качеством образования на уроках химии
Использование разнообразных форм и средств контроля за качеством образования на уроках химии Маркетинговые исследования
Маркетинговые исследования Опорные конспекты
Опорные конспекты Аннотация проекта
Аннотация проекта Эпоха Просвещения
Эпоха Просвещения ПОЭЗИЯ СЕРЕБРЯНОГО ВЕКА
ПОЭЗИЯ СЕРЕБРЯНОГО ВЕКА Relictum Lab. Крипто-бизнес
Relictum Lab. Крипто-бизнес Органы чувств
Органы чувств Учебно - тренировочный тест по математике 2
Учебно - тренировочный тест по математике 2 Культура народов мира
Культура народов мира Презентация на тему Способы повышения интереса к учебе
Презентация на тему Способы повышения интереса к учебе «Чем люди живы?»
«Чем люди живы?» Таможенные сборы
Таможенные сборы Godes_A_I
Godes_A_I Социальный эксперимент
Социальный эксперимент Художественная обработка материала, мозаика Царевна-Лебедь
Художественная обработка материала, мозаика Царевна-Лебедь Вводные слова и предложения как грамматические явления
Вводные слова и предложения как грамматические явления Как написать сочинение
Как написать сочинение Проектирование общественного здания зального типа
Проектирование общественного здания зального типа Прогрессивная Партия Большой Страны
Прогрессивная Партия Большой Страны Первый день творения (земля, вода, небо)
Первый день творения (земля, вода, небо) Наука и философия 10 класс
Наука и философия 10 класс Шаги DIRECTUM для реализации облачной стратегии
Шаги DIRECTUM для реализации облачной стратегии Тв. деят
Тв. деят История Linux и ПСПО
История Linux и ПСПО Презентация на тему Компьютерные сети Коммуникационные технологии
Презентация на тему Компьютерные сети Коммуникационные технологии