- С. С. Холод Зональность в растительном покрове острова Врангеля: синтаксономический подход

Содержание
- 2. Остров Врангеля: Протяженность с запада на восток – 110 км,с севера на юг – 80 км
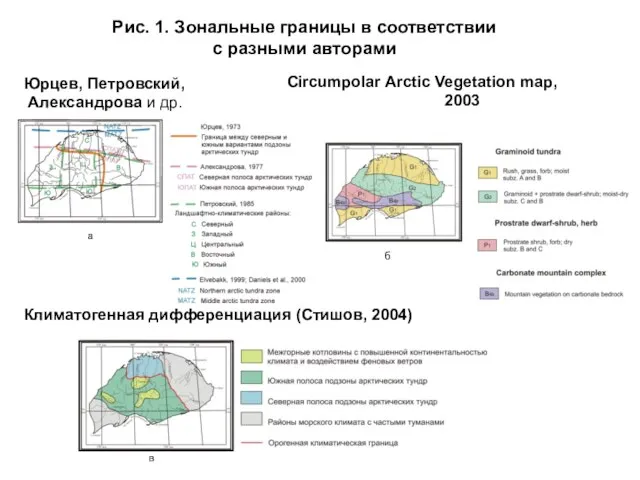
- 3. На рис. 1 представлены схемы геоботанического районирования и климатогенной дифференциации острова Врангеля, выполненные ранее. Одним из
- 4. Рис. 1. Зональные границы в соответствии с разными авторами Сircumpolar Arctic Vegetation map, 2003 Климатогенная дифференциация
- 5. Одна из основных проблем при изучении зональности – нахождение тех позиций в ландшафте, которые можно считать
- 6. Рис. 2. Обобщенный профиль равнины (0-180 м. над у. м.). Местоположение: 1 – прибрежный овраг, 2
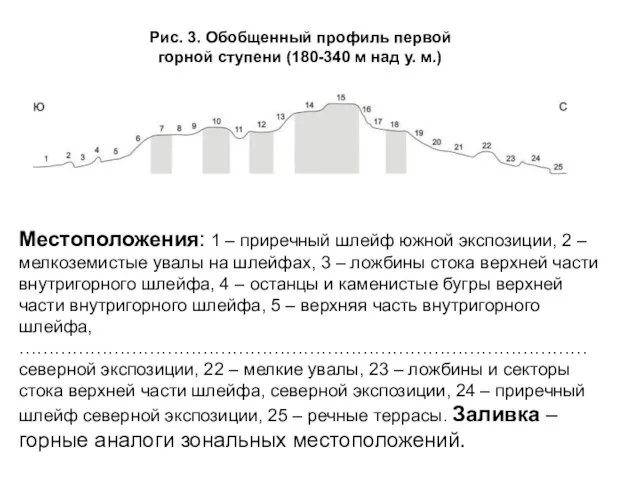
- 7. Для горных стран особенно актуально нахождение тех позиций в ландшафте, которые можно было бы считать условно
- 8. Местоположения: 1 – приречный шлейф южной экспозиции, 2 – мелкоземистые увалы на шлейфах, 3 – ложбины
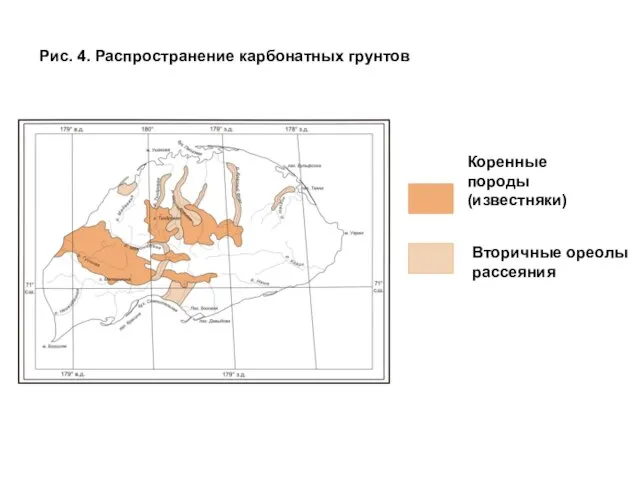
- 9. Существенное значение для определения зональных позиций ландшафта на рассматриваемой территории имеют широко распространенные здесь карбонатные грунты
- 10. Рис. 4. Распространение карбонатных грунтов Коренные породы (известняки) Вторичные ореолы рассеяния
- 11. В основу зонального деления острова Врангеля положен синтаксономический подход. Использовано 46 синтаксонов, выделенных в специальной работе
- 12. При построении таблицы зональной и оро-зональной дифференциации растительности использован метод табличной обработки данных. В строках этой
- 13. Табл. 1. Зональная дифференциация растительности
- 14. Табл. 2. Дифференцирующие синтаксоны северного варианта подзоны типичных тундр
- 15. Табл. 3. Дифференцирующие синтаксоны северных зональных полос
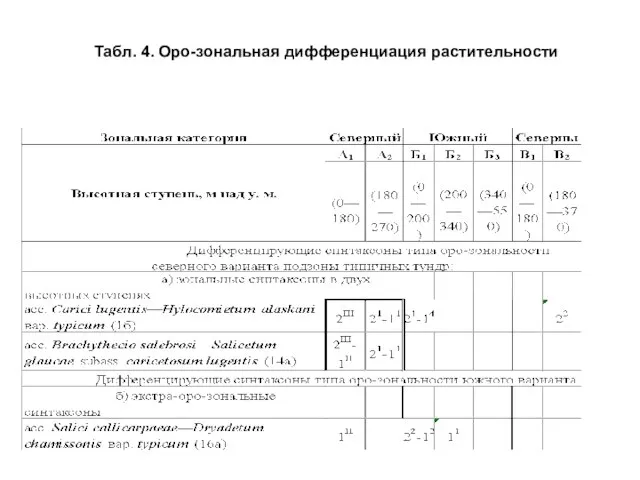
- 16. Аналогично широтной зональности, выявление дифференцирующих блоков проведено и для случая оро-зональной дифференциации растительности (табл. 4) .
- 17. Табл. 4. Оро-зональная дифференциация растительности
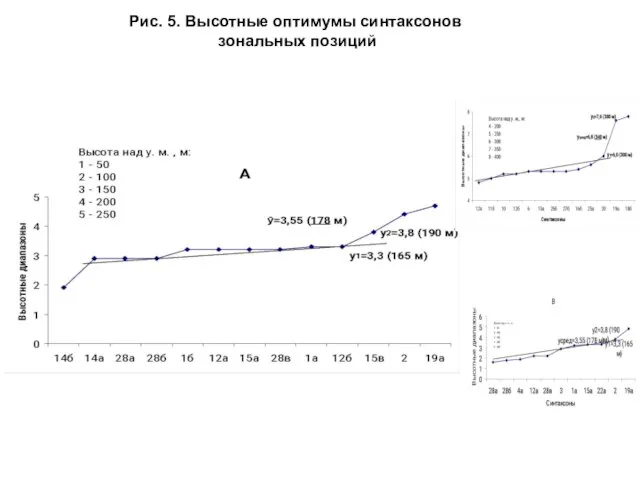
- 18. При расчете высотных оптимумов синтаксонов высотная шкала (в метрах над у. м.) разбивалась на диапазоны по
- 19. Рис. 5. Высотные оптимумы синтаксонов зональных позиций
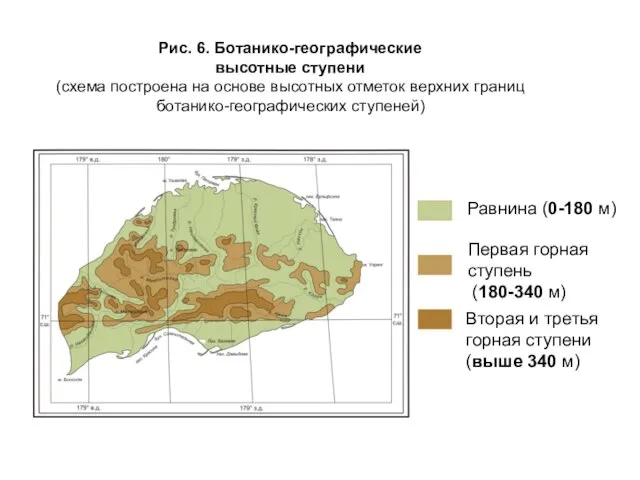
- 20. Рис. 6. Ботанико-географические высотные ступени (схема построена на основе высотных отметок верхних границ ботанико-географических ступеней) Равнина
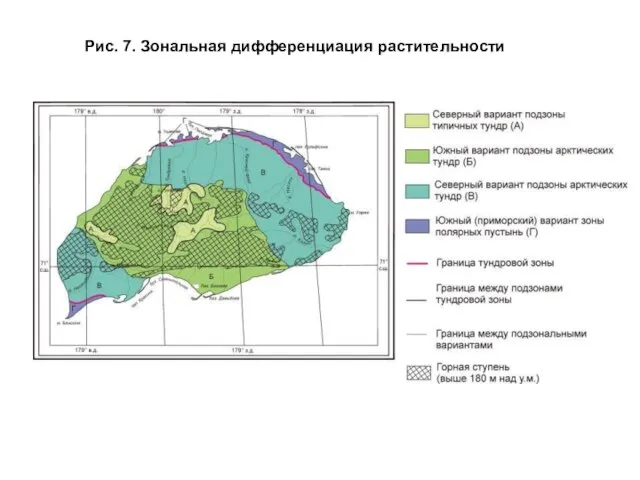
- 21. Рассмотренные выше диагностические признаки зональных полос разного ранга позволили построить схему зональной дифференциации растительности, представленную на
- 22. Рис. 7. Зональная дифференциация растительности
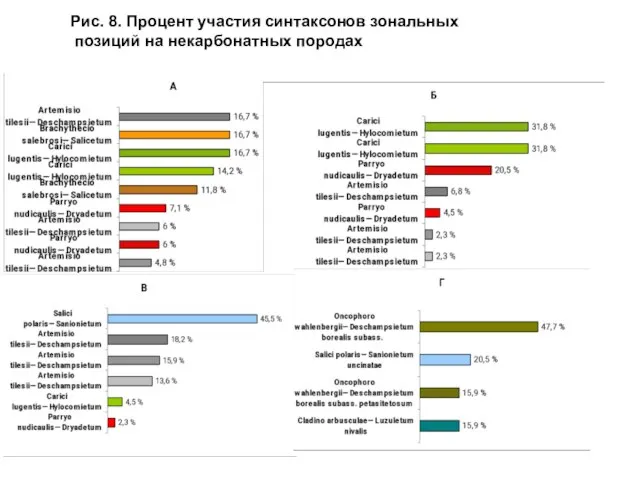
- 23. Показатель числа синтаксонов – одна из характеристик ценотического разнообразия. Этот показатель рассмотрен дифференцированно для зональных и
- 24. Рис. 8. Процент участия синтаксонов зональных позиций на некарбонатных породах
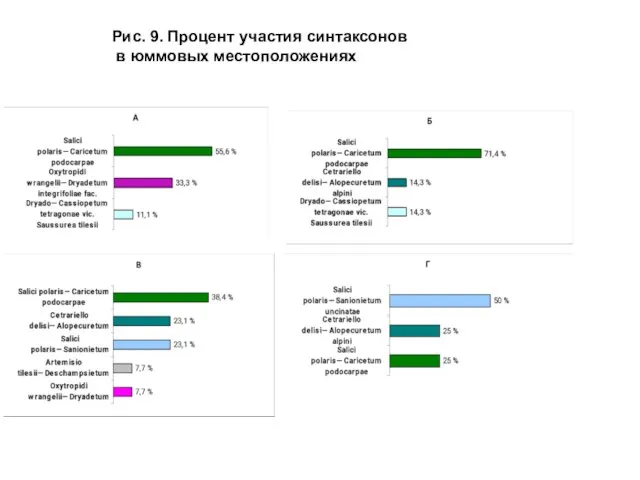
- 25. Рис. 9. Процент участия синтаксонов в юммовых местоположениях
- 26. Соотношение числа синтаксонов на зональных и азональных позициях и индексы зональности отдельно для некарбонатных и всех
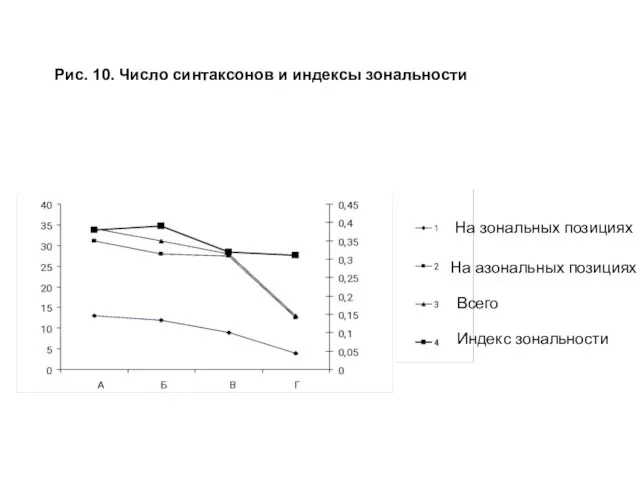
- 27. Рис. 10. Число синтаксонов и индексы зональности На зональных позициях На азональных позициях Всего Индекс зональности
- 28. Бета-разнообразие как несходство оценивалось через среднее арифметическое между значениями несходства, получаемыми при попарном сравнении видового состава
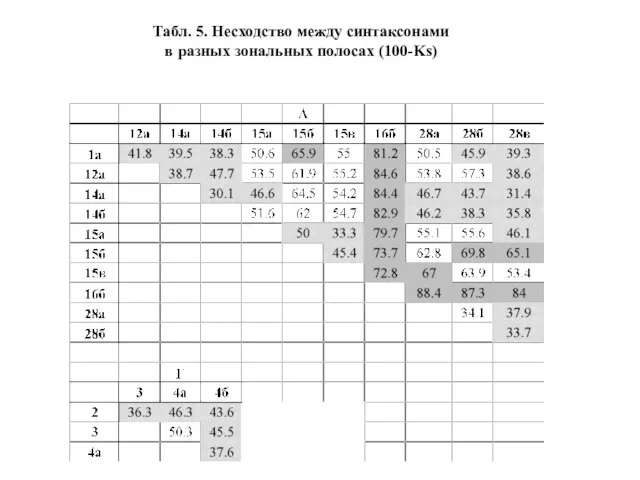
- 29. Табл. 5. Несходство между синтаксонами в разных зональных полосах (100-Ks)
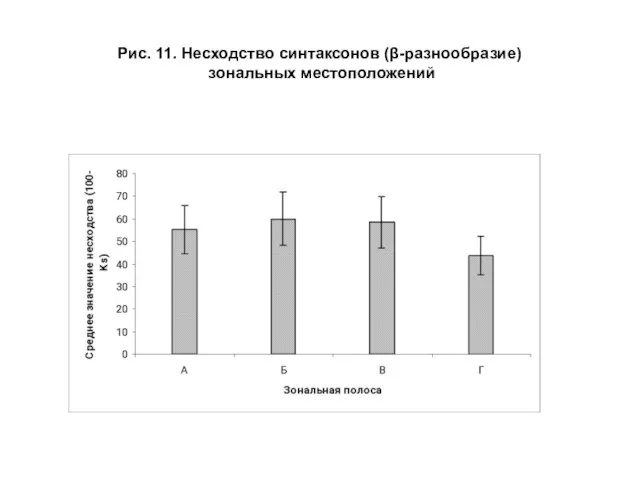
- 30. Рис. 11. Несходство синтаксонов (β-разнообразие) зональных местоположений
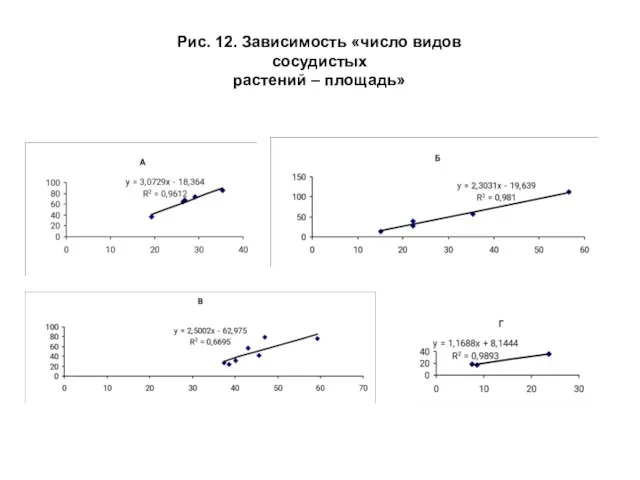
- 31. Оценка и сопоставление значений гамма (γ)- разнообразия сосудистых растений на зональных местоположениях производились на основе линейной
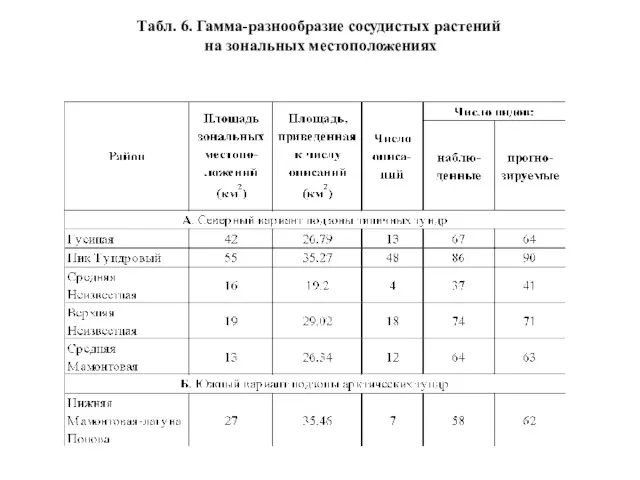
- 32. Табл. 6. Гамма-разнообразие сосудистых растений на зональных местоположениях
- 33. Рис. 12. Зависимость «число видов сосудистых растений – площадь»
- 34. Все многообразие конкретных вариантов горизонтальной структуры сообществ зональных позиций, встречающихся на острове Врангеля, сведено к нескольким
- 35. Рис. 13. Типы горизонтальной структуры
- 36. Рис. 15. Северный вариант подзоны типичных тундр (А) Заросли Salix lanata subsp. richardsonii – aсс. Parryo
- 37. Рис. 16. Южный вариант подзоны арктических тундр (В) aсс. Carici lugentis – Hylocomietum alaskani Sekretareva 1998
- 38. Рис. 17. Северный вариант подзоны арктических тундр (С) aсс. Artemisio tilesii – Deschampsietum borealis, асс. Salici
- 39. Рис. 18. Южный (приморский) вариант зоны полярных пустынь (D) aсс. Oncophoro wahlenbergii – Deschampsietum borealis; зональные
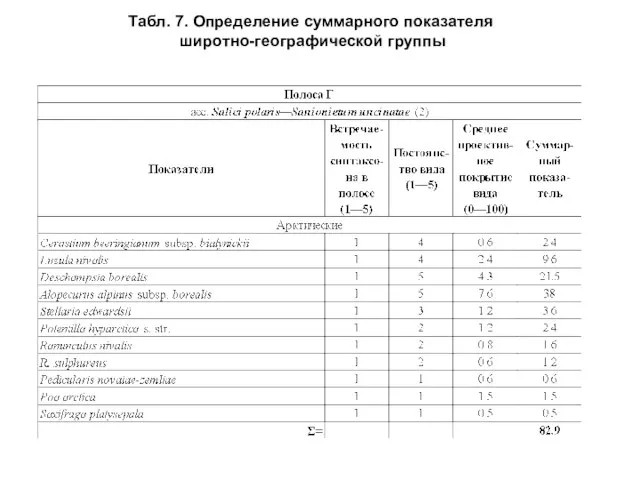
- 40. Для оценки соотношения широтно-географических групп видов использовались показатели встречаемости синтаксона, постоянства и среднего проективного покрытия вида
- 41. Табл. 7. Определение суммарного показателя широтно-географической группы
- 42. Анализ изменения спектров широтно-географических групп сосудистых растений на зональном градиенте приведен ниже (рис. 19). От полосы
- 43. Рис. 19. Широтно-географические группы сосудистых растений
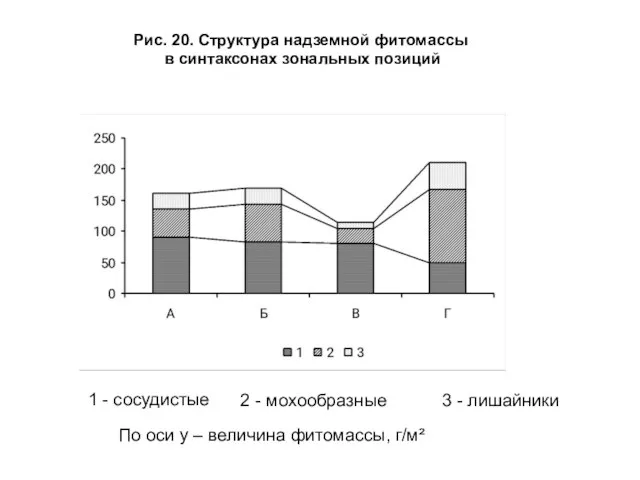
- 44. Значения общей надземной фитомассы в выборочных синтаксонах составляют: для полосы А — 161.5 г/м2, Б —
- 45. Рис. 20. Структура надземной фитомассы в синтаксонах зональных позиций 1 - сосудистые 2 - мохообразные 3
- 46. Для сосудистых растений отмечено уменьшение максимальных значений встречаемости этого показателя от полосы А к полосе Г
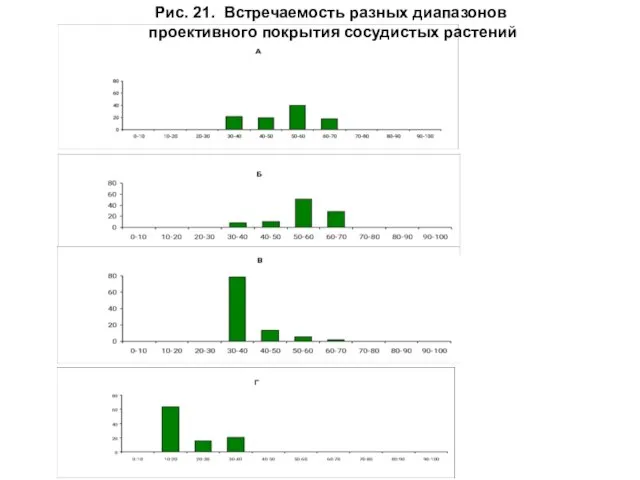
- 47. Рис. 21. Встречаемость разных диапазонов проективного покрытия сосудистых растений
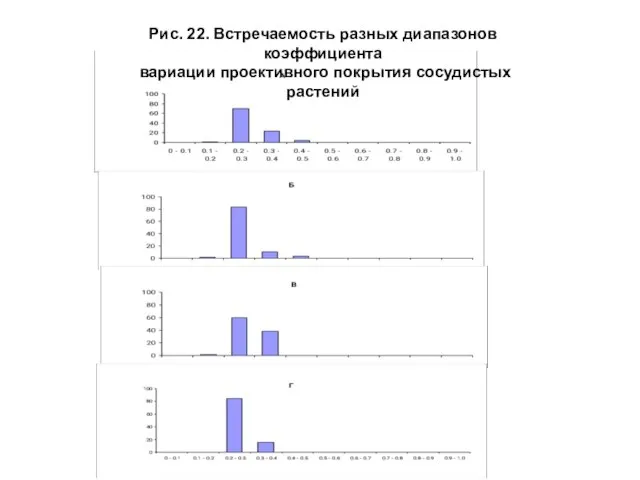
- 48. Изменение значений встречаемости коэффициента вариации проективного покрытия сосудистых растений на зональном градиенте выглядит следующим образом (рис.
- 49. Рис. 22. Встречаемость разных диапазонов коэффициента вариации проективного покрытия сосудистых растений
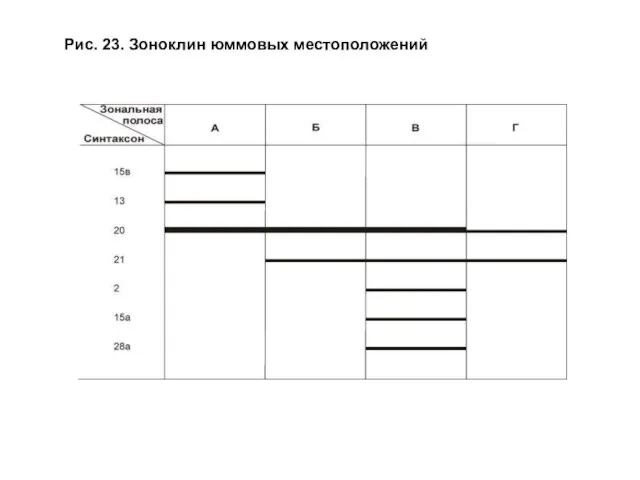
- 50. Зоноклины строились на основе методики, предложенной в работе Б. М. Миркина и Л. Г. Наумовой (1983).
- 51. Рис. 23. Зоноклин юммовых местоположений
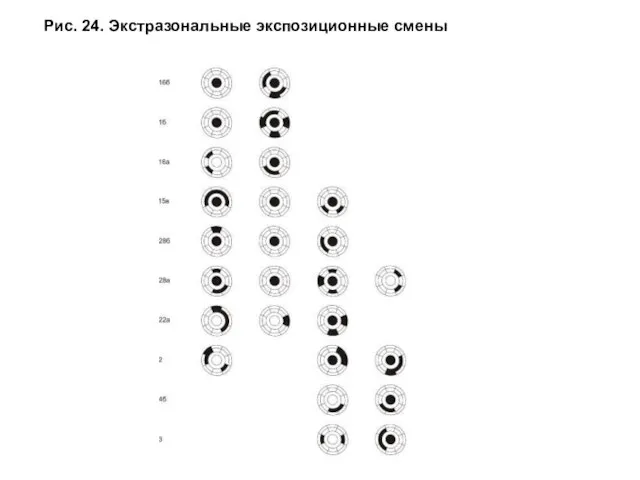
- 52. Для отображения характера экстразональных экспозиционных смен предложена круговая схема, разные секторы которой соответствуют экспозициям местоположений по
- 53. Рис. 24. Экстразональные экспозиционные смены
- 54. Проведенный выше анализ экстразональных смен позволил выявить тренд изменения экспозиции синтаксонов на зональном градиенте (рис. 25).
- 56. Скачать презентацию

Слайд 3 На рис. 1 представлены схемы геоботанического районирования и климатогенной дифференциации острова
На рис. 1 представлены схемы геоботанического районирования и климатогенной дифференциации острова

Слайд 4Рис. 1. Зональные границы в соответствии
с разными авторами
Сircumpolar Arctic Vegetation map,
Рис. 1. Зональные границы в соответствии
с разными авторами
Сircumpolar Arctic Vegetation map,
Слайд 5 Одна из основных проблем при изучении зональности – нахождение тех позиций
Одна из основных проблем при изучении зональности – нахождение тех позиций

Слайд 6Рис. 2. Обобщенный профиль равнины (0-180 м. над у. м.).
Местоположение: 1 –
Рис. 2. Обобщенный профиль равнины (0-180 м. над у. м.).
Местоположение: 1 –

Слайд 7 Для горных стран особенно актуально нахождение тех позиций в ландшафте, которые
Для горных стран особенно актуально нахождение тех позиций в ландшафте, которые

Слайд 8Местоположения: 1 – приречный шлейф южной экспозиции, 2 – мелкоземистые увалы на
Местоположения: 1 – приречный шлейф южной экспозиции, 2 – мелкоземистые увалы на
Слайд 9 Существенное значение для определения зональных позиций ландшафта на рассматриваемой территории имеют
Существенное значение для определения зональных позиций ландшафта на рассматриваемой территории имеют

Слайд 10Рис. 4. Распространение карбонатных грунтов
Коренные
породы
(известняки)
Вторичные ореолы
рассеяния
Рис. 4. Распространение карбонатных грунтов
Коренные
породы
(известняки)
Вторичные ореолы
рассеяния
Слайд 11 В основу зонального деления острова Врангеля положен синтаксономический подход. Использовано 46
В основу зонального деления острова Врангеля положен синтаксономический подход. Использовано 46

Слайд 12 При построении таблицы зональной и оро-зональной дифференциации растительности использован метод табличной
При построении таблицы зональной и оро-зональной дифференциации растительности использован метод табличной

Слайд 13Табл. 1. Зональная дифференциация растительности
Табл. 1. Зональная дифференциация растительности

Слайд 14Табл. 2. Дифференцирующие синтаксоны северного варианта
подзоны типичных тундр
Табл. 2. Дифференцирующие синтаксоны северного варианта
подзоны типичных тундр

Слайд 15Табл. 3. Дифференцирующие синтаксоны
северных зональных полос
Табл. 3. Дифференцирующие синтаксоны
северных зональных полос

Слайд 16 Аналогично широтной зональности, выявление дифференцирующих блоков проведено и для случая оро-зональной
Аналогично широтной зональности, выявление дифференцирующих блоков проведено и для случая оро-зональной

Слайд 17Табл. 4. Оро-зональная дифференциация растительности
Табл. 4. Оро-зональная дифференциация растительности
Слайд 18 При расчете высотных оптимумов синтаксонов высотная шкала (в метрах над у.
При расчете высотных оптимумов синтаксонов высотная шкала (в метрах над у.

Слайд 19Рис. 5. Высотные оптимумы синтаксонов
зональных позиций
Рис. 5. Высотные оптимумы синтаксонов
зональных позиций
Слайд 20Рис. 6. Ботанико-географические
высотные ступени
(схема построена на основе высотных отметок верхних
Рис. 6. Ботанико-географические
высотные ступени
(схема построена на основе высотных отметок верхних
Слайд 21 Рассмотренные выше диагностические признаки зональных полос разного ранга позволили построить схему
Рассмотренные выше диагностические признаки зональных полос разного ранга позволили построить схему

Слайд 22Рис. 7. Зональная дифференциация растительности
Рис. 7. Зональная дифференциация растительности
Слайд 23 Показатель числа синтаксонов – одна из характеристик ценотического разнообразия. Этот показатель
Показатель числа синтаксонов – одна из характеристик ценотического разнообразия. Этот показатель

Слайд 24Рис. 8. Процент участия синтаксонов зональных
позиций на некарбонатных породах
Рис. 8. Процент участия синтаксонов зональных
позиций на некарбонатных породах
Слайд 25Рис. 9. Процент участия синтаксонов
в юммовых местоположениях
Рис. 9. Процент участия синтаксонов
в юммовых местоположениях
Слайд 26 Соотношение числа синтаксонов на зональных и азональных позициях и индексы зональности
Соотношение числа синтаксонов на зональных и азональных позициях и индексы зональности

Слайд 27Рис. 10. Число синтаксонов и индексы зональности
На зональных позициях
На азональных позициях
Всего
Индекс
Рис. 10. Число синтаксонов и индексы зональности
На зональных позициях
На азональных позициях
Всего
Индекс
Слайд 28 Бета-разнообразие как несходство оценивалось через среднее арифметическое между значениями несходства, получаемыми
Бета-разнообразие как несходство оценивалось через среднее арифметическое между значениями несходства, получаемыми

Слайд 29Табл. 5. Несходство между синтаксонами
в разных зональных полосах (100-Ks)
Табл. 5. Несходство между синтаксонами
в разных зональных полосах (100-Ks)
Слайд 30Рис. 11. Несходство синтаксонов (β-разнообразие)
зональных местоположений
Рис. 11. Несходство синтаксонов (β-разнообразие)
зональных местоположений
Слайд 31 Оценка и сопоставление значений гамма (γ)- разнообразия сосудистых растений на зональных
Оценка и сопоставление значений гамма (γ)- разнообразия сосудистых растений на зональных

Слайд 32Табл. 6. Гамма-разнообразие сосудистых растений
на зональных местоположениях
Табл. 6. Гамма-разнообразие сосудистых растений
на зональных местоположениях
Слайд 33Рис. 12. Зависимость «число видов сосудистых
растений – площадь»
Рис. 12. Зависимость «число видов сосудистых
растений – площадь»
Слайд 34 Все многообразие конкретных вариантов горизонтальной структуры сообществ зональных позиций, встречающихся на
Все многообразие конкретных вариантов горизонтальной структуры сообществ зональных позиций, встречающихся на

Слайд 35Рис. 13. Типы горизонтальной структуры
Рис. 13. Типы горизонтальной структуры

Слайд 36Рис. 15. Северный вариант подзоны типичных тундр (А)
Заросли Salix lanata subsp. richardsonii
Рис. 15. Северный вариант подзоны типичных тундр (А)
Заросли Salix lanata subsp. richardsonii

Слайд 37Рис. 16. Южный вариант подзоны арктических тундр (В)
aсс. Carici lugentis – Hylocomietum
Рис. 16. Южный вариант подзоны арктических тундр (В)
aсс. Carici lugentis – Hylocomietum

Слайд 38Рис. 17. Северный вариант подзоны арктических тундр (С)
aсс. Artemisio tilesii – Deschampsietum
Рис. 17. Северный вариант подзоны арктических тундр (С)
aсс. Artemisio tilesii – Deschampsietum

Слайд 39Рис. 18. Южный (приморский) вариант зоны полярных пустынь (D)
aсс. Oncophoro wahlenbergii –
Рис. 18. Южный (приморский) вариант зоны полярных пустынь (D)
aсс. Oncophoro wahlenbergii –

Слайд 40 Для оценки соотношения широтно-географических групп видов использовались показатели встречаемости синтаксона, постоянства
Для оценки соотношения широтно-географических групп видов использовались показатели встречаемости синтаксона, постоянства

Слайд 41Табл. 7. Определение суммарного показателя
широтно-географической группы
Табл. 7. Определение суммарного показателя
широтно-географической группы
Слайд 42 Анализ изменения спектров широтно-географических групп сосудистых растений на зональном градиенте приведен
Анализ изменения спектров широтно-географических групп сосудистых растений на зональном градиенте приведен

Слайд 43Рис. 19. Широтно-географические группы
сосудистых растений
Рис. 19. Широтно-географические группы
сосудистых растений

Слайд 44 Значения общей надземной фитомассы в выборочных синтаксонах составляют: для полосы А
Значения общей надземной фитомассы в выборочных синтаксонах составляют: для полосы А

Слайд 45Рис. 20. Структура надземной фитомассы
в синтаксонах зональных позиций
1 - сосудистые
2
Рис. 20. Структура надземной фитомассы
в синтаксонах зональных позиций
1 - сосудистые
2
Слайд 46 Для сосудистых растений отмечено уменьшение максимальных значений встречаемости этого показателя от
Для сосудистых растений отмечено уменьшение максимальных значений встречаемости этого показателя от

Слайд 47Рис. 21. Встречаемость разных диапазонов
проективного покрытия сосудистых растений
Рис. 21. Встречаемость разных диапазонов
проективного покрытия сосудистых растений
Слайд 48 Изменение значений встречаемости коэффициента вариации проективного покрытия сосудистых растений на зональном
Изменение значений встречаемости коэффициента вариации проективного покрытия сосудистых растений на зональном

Слайд 49Рис. 22. Встречаемость разных диапазонов коэффициента
вариации проективного покрытия сосудистых растений
Рис. 22. Встречаемость разных диапазонов коэффициента
вариации проективного покрытия сосудистых растений
Слайд 50 Зоноклины строились на основе методики, предложенной в работе Б. М. Миркина
Зоноклины строились на основе методики, предложенной в работе Б. М. Миркина

Слайд 51Рис. 23. Зоноклин юммовых местоположений
Рис. 23. Зоноклин юммовых местоположений
Слайд 52 Для отображения характера экстразональных экспозиционных смен предложена круговая схема, разные секторы
Для отображения характера экстразональных экспозиционных смен предложена круговая схема, разные секторы

Слайд 53Рис. 24. Экстразональные экспозиционные смены
Рис. 24. Экстразональные экспозиционные смены
Слайд 54 Проведенный выше анализ экстразональных смен позволил выявить тренд изменения экспозиции синтаксонов
Проведенный выше анализ экстразональных смен позволил выявить тренд изменения экспозиции синтаксонов

 Правовое регулирование цен и ценообразования в Республике Беларусь
Правовое регулирование цен и ценообразования в Республике Беларусь Задание на практику
Задание на практику Полістирол
Полістирол Обработка и отделка фанеры. Фанерное и плитное производство
Обработка и отделка фанеры. Фанерное и плитное производство Технология переработки рыбных отходов
Технология переработки рыбных отходов Схемы охвата
Схемы охвата Документальный фильм-портрет “Раиса”
Документальный фильм-портрет “Раиса” Ежемесячное пожизненное содержание президента РФ
Ежемесячное пожизненное содержание президента РФ DIGITAL OUT-OF-HOME
DIGITAL OUT-OF-HOME Разработка урока с применением НИТавтор: Мараховский С. А. учитель физики МОУ «Европейский лицей»
Разработка урока с применением НИТавтор: Мараховский С. А. учитель физики МОУ «Европейский лицей» Назначение и устройство компьютера
Назначение и устройство компьютера ФИЗИКА
ФИЗИКА Презентация на тему Проект "Птицы"
Презентация на тему Проект "Птицы" Система правового регулирования отношений Россия - ЕС
Система правового регулирования отношений Россия - ЕС Трудовая деятельность
Трудовая деятельность  Индивидуальный проект обучающегося
Индивидуальный проект обучающегося Теория денег
Теория денег Святая Иулиания Новоторжская и Вяземская
Святая Иулиания Новоторжская и Вяземская Векторы в пространстве
Векторы в пространстве Преподавание истории в школах Болгарии
Преподавание истории в школах Болгарии Итоги и перспективы экономических реформ в Российской Федерации
Итоги и перспективы экономических реформ в Российской Федерации Презентация на тему Металлы
Презентация на тему Металлы Презентация на тему Методика расследования дел об административном правонарушении в области таможенного дела
Презентация на тему Методика расследования дел об административном правонарушении в области таможенного дела Эклектика в архитектуре
Эклектика в архитектуре Порядок и условия заключения договора о ТО и ремонте ВДГО и ВКГО
Порядок и условия заключения договора о ТО и ремонте ВДГО и ВКГО Изобразительное искусство. Орнамент
Изобразительное искусство. Орнамент История национального костюма
История национального костюма Die Liebesgeschichte von Adriano Celentano und Claudia Mori
Die Liebesgeschichte von Adriano Celentano und Claudia Mori