- Экология фотосинтеза

Содержание
- 2. Эколого-физиологический аспект процесса фотосинтеза важен для понимания закономерностей формирования продуктивности растений и роли фотосинтеза в адаптации
- 3. В условиях недостатка углекислого газа, при водном дефиците и перегреве листовой пластинки может проявляться полуденная депрессия
- 4. На принципиальные промежуточные процессы фотосинтеза, такие, как работа реакционных центров фотосистем, транспорт электронов, фотофосфорилирование, восстановление ФГК,
- 5. Важным показателем, характеризующим эффективность работы фото-синтетического аппарата растений, является квантовый расход фотосинтеза – число квантов света,
- 6. У древесных и кустарниковых растений онтогенез листа почти не зави- сит от возраста самого растения. Весной
- 7. Хорошо исследовано изменение фотосинтеза в онтогенезе хвойных растений. Наиболее интенсивный фотосинтез наблюдают в период интенсивного роста
- 9. Скачать презентацию
Слайд 2Эколого-физиологический аспект процесса фотосинтеза важен для понимания закономерностей формирования продуктивности растений и
Эколого-физиологический аспект процесса фотосинтеза важен для понимания закономерностей формирования продуктивности растений и
Слайд 3В условиях недостатка углекислого газа, при водном дефиците и перегреве листовой пластинки
В условиях недостатка углекислого газа, при водном дефиците и перегреве листовой пластинки

Слайд 4На принципиальные промежуточные процессы фотосинтеза, такие, как работа реакционных центров фотосистем, транспорт
На принципиальные промежуточные процессы фотосинтеза, такие, как работа реакционных центров фотосистем, транспорт

Слайд 5Важным показателем, характеризующим эффективность работы фото-синтетического аппарата растений, является квантовый расход фотосинтеза
Важным показателем, характеризующим эффективность работы фото-синтетического аппарата растений, является квантовый расход фотосинтеза

Слайд 6У древесных и кустарниковых растений онтогенез листа почти не зави-
сит от возраста
У древесных и кустарниковых растений онтогенез листа почти не зави- сит от возраста

Слайд 7Хорошо исследовано изменение фотосинтеза в онтогенезе хвойных растений. Наиболее интенсивный фотосинтез наблюдают
Хорошо исследовано изменение фотосинтеза в онтогенезе хвойных растений. Наиболее интенсивный фотосинтез наблюдают

 Ткани растений. Физиология растения. Агротехнические приемы
Ткани растений. Физиология растения. Агротехнические приемы Презентация на тему Викторина по биологии "Как называется"
Презентация на тему Викторина по биологии "Как называется"  Все о собаках
Все о собаках Синдромы поражения краниальных нервов
Синдромы поражения краниальных нервов Полезная еда. Проростки гречки
Полезная еда. Проростки гречки Топография поверхностей (рельеф) полушарий мозга
Топография поверхностей (рельеф) полушарий мозга Медицинская гельминтология
Медицинская гельминтология Соцветия. Схемы соцветий
Соцветия. Схемы соцветий Геном про – та еукаріот
Геном про – та еукаріот Обмен веществ и энергии
Обмен веществ и энергии Биологическое действие радиации. Закон радиоактивного распада. Термоядерные реакции
Биологическое действие радиации. Закон радиоактивного распада. Термоядерные реакции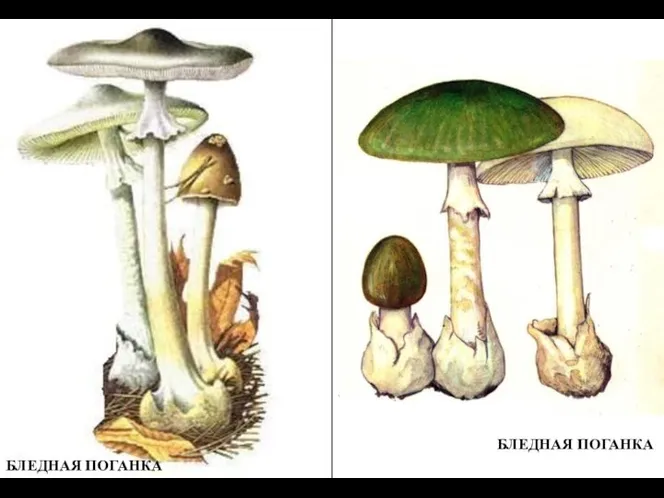 Наглядный материал к предложенным занятиям - Грибы
Наглядный материал к предложенным занятиям - Грибы Влияние комнатных растений
Влияние комнатных растений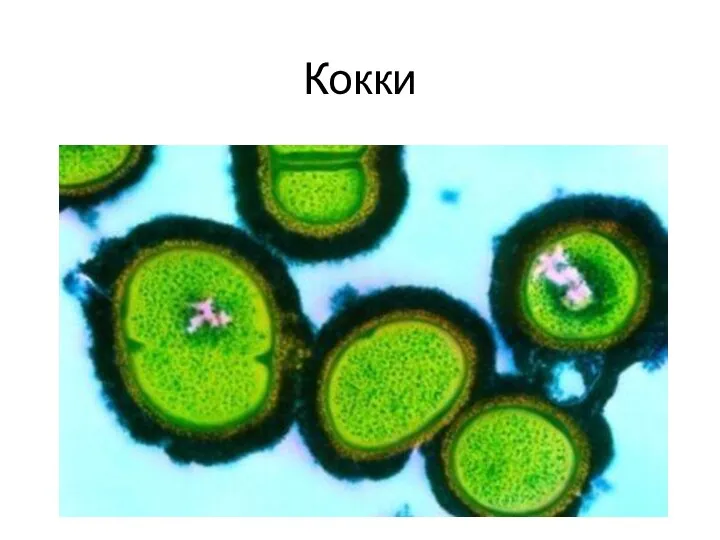 Виды бактерий
Виды бактерий Занятие 6. Основные и выделительные ткани
Занятие 6. Основные и выделительные ткани Состав, строение и рост костей. 8 класс
Состав, строение и рост костей. 8 класс Platinum. 70% парного мяса для вашего питомца
Platinum. 70% парного мяса для вашего питомца Многообразие млекопитающих. 7 класс
Многообразие млекопитающих. 7 класс Саванна
Саванна Животный мир Песчанокопского района
Животный мир Песчанокопского района Анатомия и физиология кожи
Анатомия и физиология кожи Значение опорно-двигательного аппарата, его состав . Строение костей. 8 класс
Значение опорно-двигательного аппарата, его состав . Строение костей. 8 класс Контрольное тестирование по теме Анализаторы. Значение анализаторов в жизни человека
Контрольное тестирование по теме Анализаторы. Значение анализаторов в жизни человека Комнатные растения
Комнатные растения Мутации. Виды мутаций
Мутации. Виды мутаций Утомление при статической и динамической нагрузке
Утомление при статической и динамической нагрузке Бактерии, их строение
Бактерии, их строение Олигосахариды и их классификация
Олигосахариды и их классификация