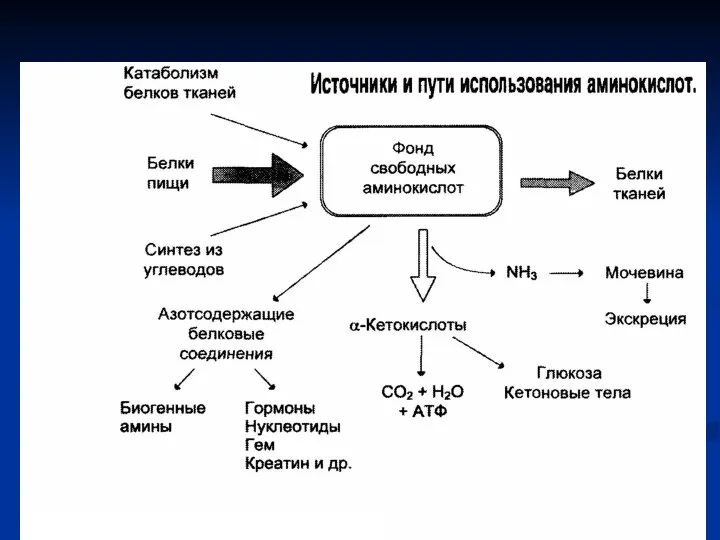
- Фонд свободных аминокислот

Содержание
- 2. Источники аминокислот 1. Белки корма (экзогенные аминокислоты). 2. Белки тканей при распаде в процессе их обновления
- 3. Пути использования аминокислот в организме Биосинтез белков, специфичных для данного организма. Синтез биологически активных веществ (ферментов,
- 5. Синтез белков 1. Транскрипция 2. Трансляция В каждом этапе различают подэтапы: Инициация; Элонгация; Терминация.
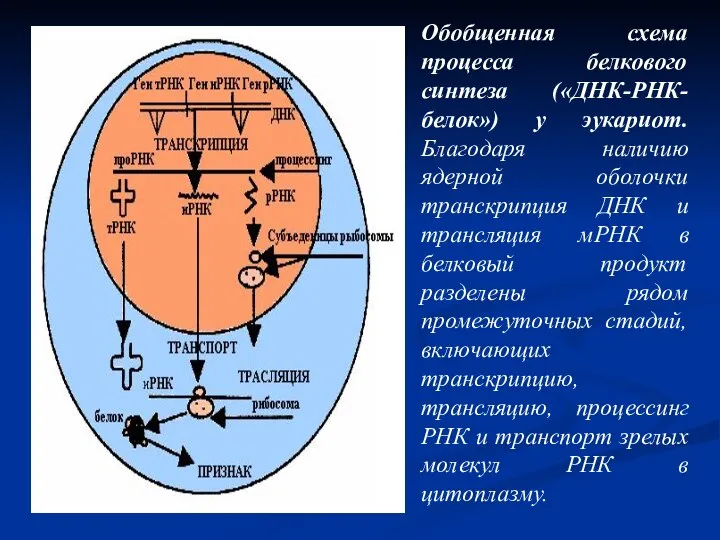
- 6. Обобщенная схема процесса белкового синтеза («ДНК-РНК-белок») у эукариот. Благодаря наличию ядерной оболочки транскрипция ДНК и трансляция
- 7. Транскрипция. Синтез РНК
- 8. Общая характеристика процесса транскрипции Молекулы ДНК каждой клетки содержат информацию для синтеза всех необходимых ей белков.
- 9. Считывание наследственной информации с генов регулируется специальными молекулами белков. В частности, гистоновые белки не только обеспечивают
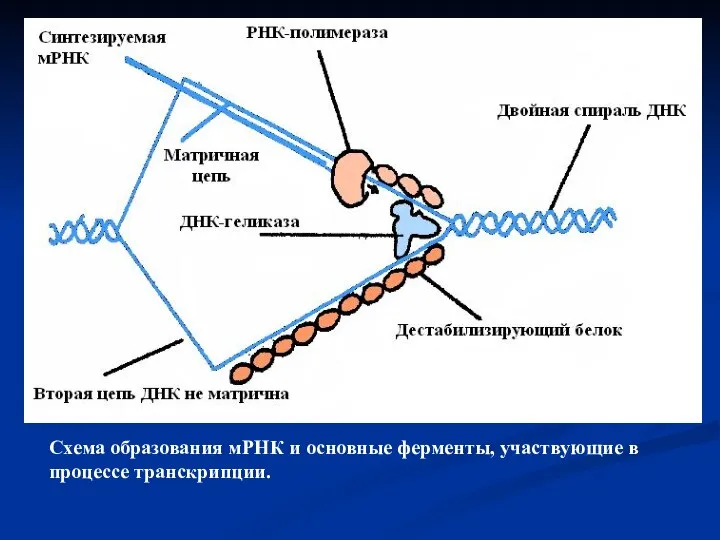
- 10. Схема образования мРНК и основные ферменты, участвующие в процессе транскрипции.
- 11. Инициация транскрипции По молекулярному сигналу из цитоплазмы определенный участок двойной спирали ДНК разворачивается и разделяется на
- 12. Элонгация цепи РНК. Трифосфаты рибонуклеотидов (АТФ, ГТФ, ЦТФ, УТФ), служат сырьем для синтеза РНК. Они присоединяются
- 13. Три различных типа РНК у эукариот, а именно: мРНК, рРНК и тРНК транскрибируются на различных участках
- 14. Терминация транскрипции РНК-полимераза движется вдоль цепи ДНК и переписывает информацию в виде про-РНК. Синтез РНК завершается
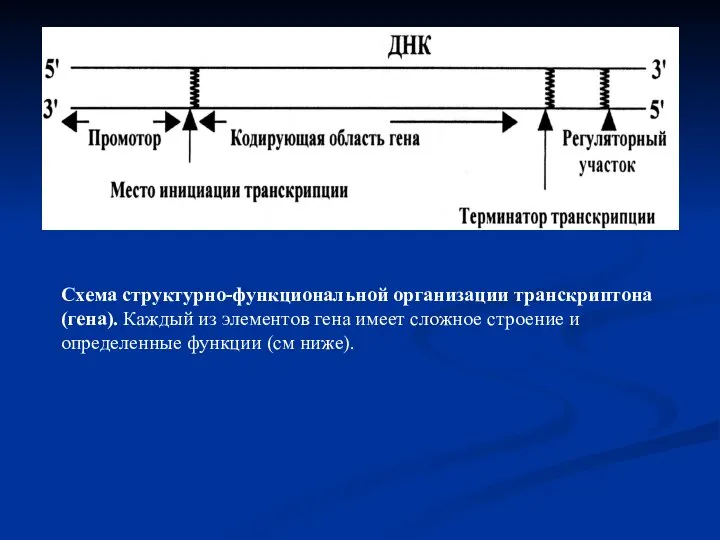
- 15. Схема структурно-функциональной организации транскриптона (гена). Каждый из элементов гена имеет сложное строение и определенные функции (см
- 16. Освободившийся от РНК участок молекулы ДНК вновь связывается водородными связями с комплементарной областью второй цепи. ДНК
- 17. Значение транскрипции: а) перенос информации с ДНК на РНК является первым этапом синтеза всех разновидностей белков
- 18. Процессинг. Экзоны и интроны. Сплайсинг. Молекулярные механизмы, связанные с «созреванием» различных типов РНК, называются процессингом. Они
- 19. Модификация про-мРНК начинается сразу после окончания его синтеза. На 5' конце образуется «колпачок» - КЭП, который
- 20. Схема процессинга мРНК и его место в процессе экспрессии гена. Транскрипция приводит к образованию крупной молекулы
- 21. Сплайсинг РНК. В процессе сплайсинга принимают участие различные мяРНП, которые формируют сплайсосому. мяРНП, взаимодействуя с РНК
- 22. Процессинг пре-тРНК. Определенные азотистые основания нуклеотидов тРНК в ходе процессинга метилируються под действием РНК-метилазы и превращаются,
- 23. В процессинге участвует целый ряд ферментов. Например, с помощью нуклеаз вырезаются интронные участки, а остающиеся экзонные
- 24. Существует также альтернативный процессинг, в результате которого из одной и той же молекулы про-мРНК могут удаляться
- 25. Альтернативный сплайсинг гена кальцитонина. В клетках щитовидной железы сплайсинг первичного транскрипта приводит к образованию кальцитониновой мРНК,
- 26. Итак, процессинг включает в себя несколько важных процессов: а) кэппирование РНК, б) удаление части нуклеотидов на
- 27. Трансляция. Синтез белков
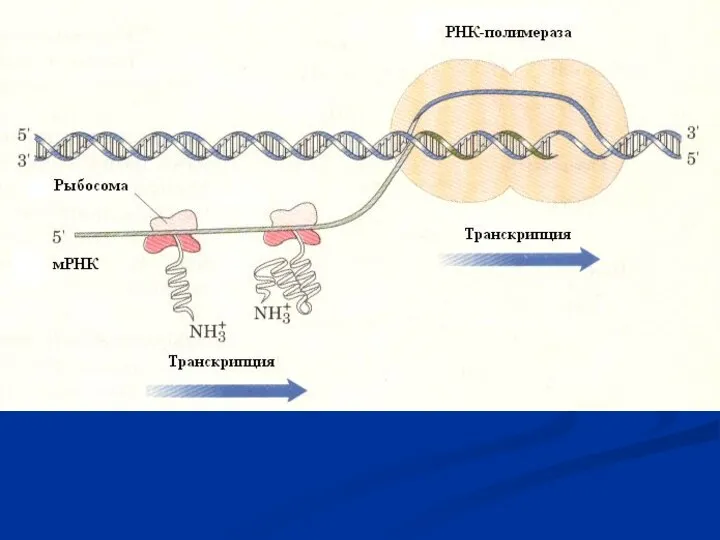
- 28. Общая характеристика процесса Информация, содержащаяся в ДНК, передается синтезируемому белку через РНК. Процесс переноса информации с
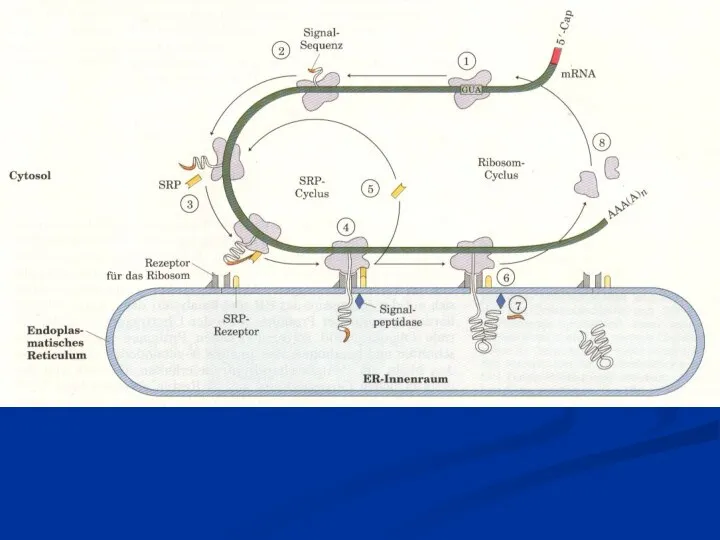
- 29. После «созревания» мРНК (процессинг) переносится в цитоплазму. Кэппированный конец мРНК, распознается малой субъединицей рибосом, так что
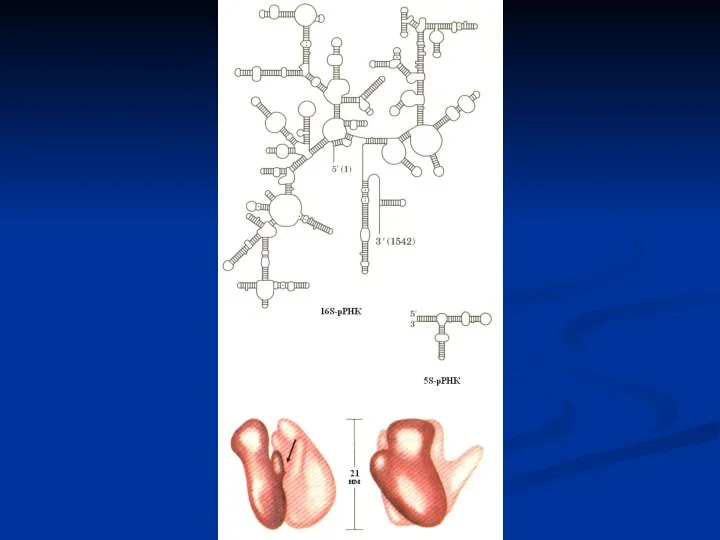
- 30. Рибосома эукариот. А – рибосома 8ОS состоит из большой и малой субъединицы. Большая субъединица содержит 5S,
- 31. Рибосомы – это органеллы клеток, обеспечивающие синтез полипептидов. Они объединяют в пространстве и времени все вещества,
- 32. Схематическое изображение полирибосомы показывающее, как ряд рибосом одновременно осуществляет трансляцию на одной и той же молекуле
- 33. Образование и выход из ядра субъединиц рибосом.
- 34. С цепью мРНК одновременно может связаться много рибосом. Так образуются полирибосомы (полисомы), что существенно увеличивает эффективность
- 35. Модель молекулярного механизма работы рибосом была предложена А. Спириным в 1968 году. Рибосома имеет две бороздки:
- 36. Основные компоненты белоксинтезирующей системы
- 37. Инициация синтеза белков. Активация аминокислот. Аминокислоты (АК) в цитозоле клетки вступают в реакцию с АТФ. Так
- 38. Схема инициации 1. Активация аминокислот – их связывание с соответствующей тРНК с помощью фермента аминоацил-тРНК-синтетазы; 2.
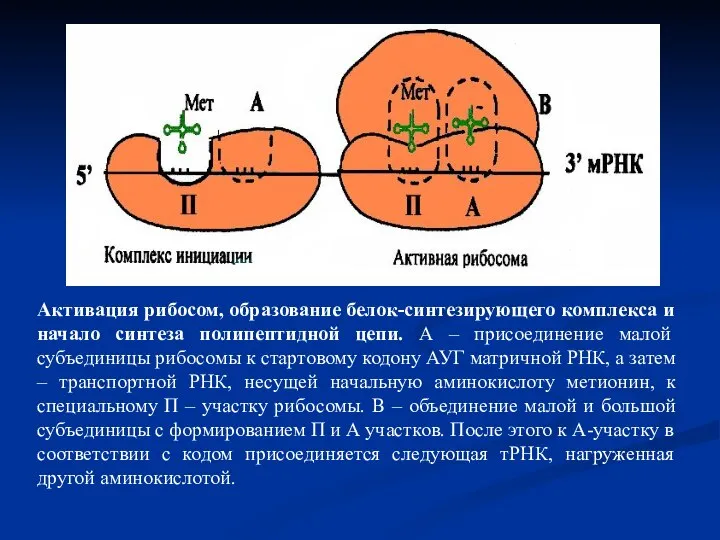
- 39. Активация рибосом, образование белок-синтезирующего комплекса и начало синтеза полипептидной цепи. А – присоединение малой субъединицы рибосомы
- 40. Комплементарность – строгое соответствие азотистых оснований одной цепи ДНК другой (А – Т, Г – Ц)
- 41. Генетический код – строгая последовательность триплетов (кодонов) в ДНК и м-РНК, контролирует порядок расположения аминокислот в
- 42. Характеристика генетического кода 1. Триплетность. 2. Специфичность. 3. Неперекрываемость. 4. Универсальность. 5. Избыточность. 6. Колинеарность. 7.
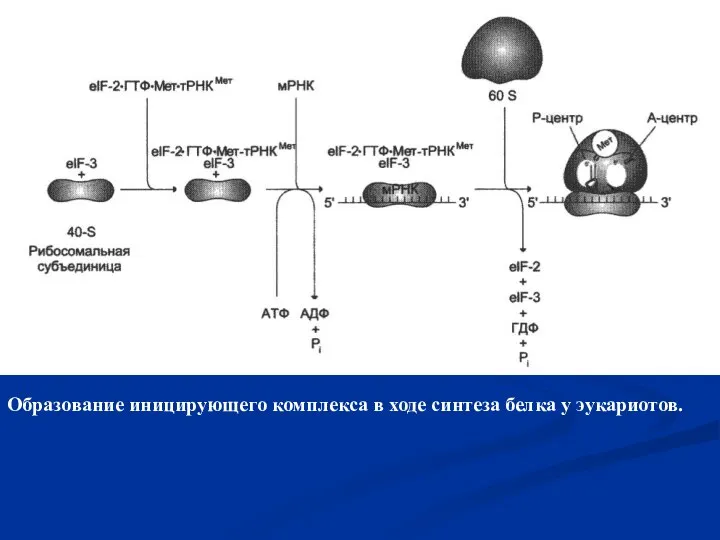
- 43. Образование иницирующего комплекса в ходе синтеза белка у эукариотов.
- 44. Активация рибосом и начало синтеза полипептидной цепи. Цепь мРНК соединяется с малой рибосомальной субъединицей при помощи
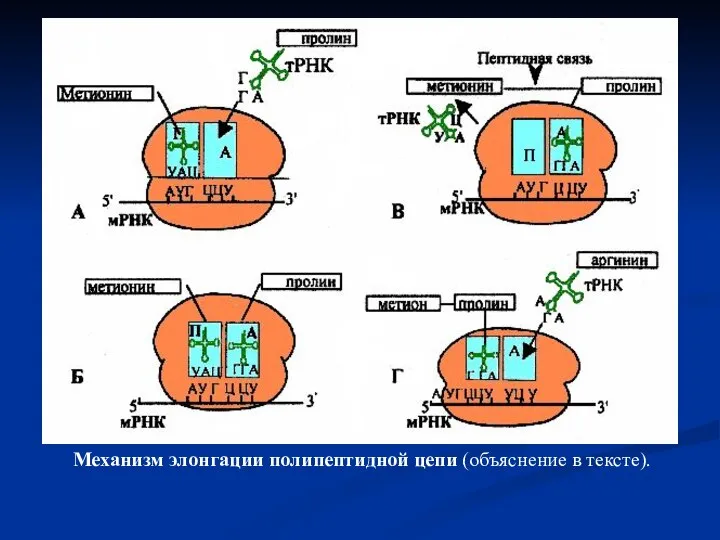
- 47. Элонгация полипептидной цепи Удлинение полипептида. Вторая нагруженная, например пролином, тРНК соединяется с рибосомой на участке А.
- 48. тРНК-дипептидный комплекс на участке А рибосомы вместе с мРНК перемещается относительно рибосомы в направлении участка П.
- 49. Механизм элонгации полипептидной цепи (объяснение в тексте).
- 50. Весь процесс, включающий поступление тРНК-аминокислотного комплекса, образования пептидной связи и транслокации, многократно повторяется. По мере продвижения
- 52. Терминация синтеза белков. Окончание синтеза и высвобождение полипептидной цепи. В конце цепи мРНК находится один из
- 53. Посттрансляционная модификация полипептидной цепи белка («созревание») Синтезированый из аминокислот полипептид - это практически прямолинейная молекула, не
- 55. Значение синтеза белков Синтез белков является основным процессом в жизни клетки и организма. Жизнь многоклеточного индивидуума
- 56. Основная догма молекулярной биологии Хромосомные ДНК содержат полную информацию обо всех белках, синтезируемых в клетке. Эта
- 58. Скачать презентацию
Слайд 2Источники аминокислот
1. Белки корма (экзогенные аминокислоты).
2. Белки тканей при распаде в процессе
Источники аминокислот
1. Белки корма (экзогенные аминокислоты).
2. Белки тканей при распаде в процессе

Слайд 3Пути использования аминокислот в организме
Биосинтез белков, специфичных для данного организма.
Синтез биологически активных
Пути использования аминокислот в организме
Биосинтез белков, специфичных для данного организма.
Синтез биологически активных

Слайд 5Синтез белков
1. Транскрипция
2. Трансляция
В каждом этапе различают подэтапы:
Инициация;
Элонгация;
Терминация.
Синтез белков
1. Транскрипция
2. Трансляция
В каждом этапе различают подэтапы:
Инициация;
Элонгация;
Терминация.

Слайд 6Обобщенная схема процесса белкового синтеза («ДНК-РНК-белок») у эукариот. Благодаря наличию ядерной оболочки
Слайд 7Транскрипция. Синтез РНК
Транскрипция. Синтез РНК

Слайд 8 Общая характеристика процесса транскрипции
Молекулы ДНК каждой клетки содержат информацию для
Общая характеристика процесса транскрипции
Молекулы ДНК каждой клетки содержат информацию для

Слайд 9 Считывание наследственной информации с генов регулируется специальными молекулами белков. В частности,
Считывание наследственной информации с генов регулируется специальными молекулами белков. В частности,

Слайд 10Схема образования мРНК и основные ферменты, участвующие в процессе транскрипции.
Схема образования мРНК и основные ферменты, участвующие в процессе транскрипции.
Слайд 11 Инициация транскрипции
По молекулярному сигналу из цитоплазмы определенный участок двойной спирали
Инициация транскрипции
По молекулярному сигналу из цитоплазмы определенный участок двойной спирали

Слайд 12 Элонгация цепи РНК.
Трифосфаты рибонуклеотидов (АТФ, ГТФ, ЦТФ, УТФ), служат сырьем для
Элонгация цепи РНК.
Трифосфаты рибонуклеотидов (АТФ, ГТФ, ЦТФ, УТФ), служат сырьем для

Слайд 13 Три различных типа РНК у эукариот, а именно: мРНК, рРНК и
Три различных типа РНК у эукариот, а именно: мРНК, рРНК и

Слайд 14 Терминация транскрипции
РНК-полимераза движется вдоль цепи ДНК и переписывает информацию в
Терминация транскрипции
РНК-полимераза движется вдоль цепи ДНК и переписывает информацию в

Слайд 15Схема структурно-функциональной организации транскриптона (гена). Каждый из элементов гена имеет сложное строение
Слайд 16 Освободившийся от РНК участок молекулы ДНК вновь связывается водородными связями с
Освободившийся от РНК участок молекулы ДНК вновь связывается водородными связями с

Слайд 17 Значение транскрипции:
а) перенос информации с ДНК на РНК является первым
Значение транскрипции:
а) перенос информации с ДНК на РНК является первым

Слайд 18 Процессинг. Экзоны и интроны. Сплайсинг.
Молекулярные механизмы, связанные с «созреванием» различных
Процессинг. Экзоны и интроны. Сплайсинг.
Молекулярные механизмы, связанные с «созреванием» различных

Слайд 19 Модификация про-мРНК начинается сразу после окончания его синтеза. На 5' конце
Модификация про-мРНК начинается сразу после окончания его синтеза. На 5' конце

Слайд 20Схема процессинга мРНК и его место в процессе экспрессии гена. Транскрипция приводит
Схема процессинга мРНК и его место в процессе экспрессии гена. Транскрипция приводит
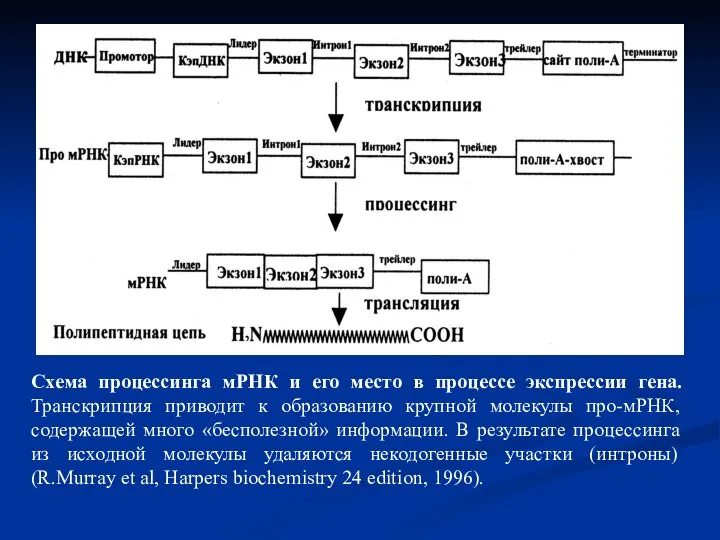
Слайд 21 Сплайсинг РНК. В процессе сплайсинга принимают участие различные мяРНП, которые формируют
Сплайсинг РНК. В процессе сплайсинга принимают участие различные мяРНП, которые формируют
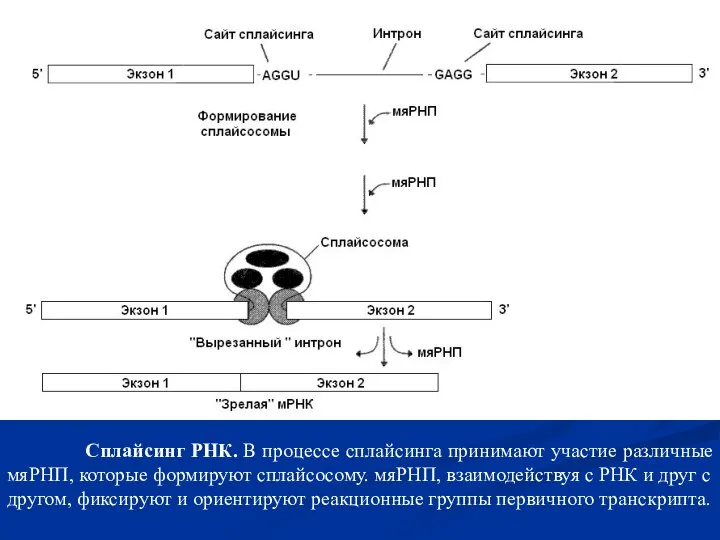
Слайд 22 Процессинг пре-тРНК. Определенные азотистые основания нуклеотидов тРНК в ходе процессинга метилируються
Процессинг пре-тРНК. Определенные азотистые основания нуклеотидов тРНК в ходе процессинга метилируються
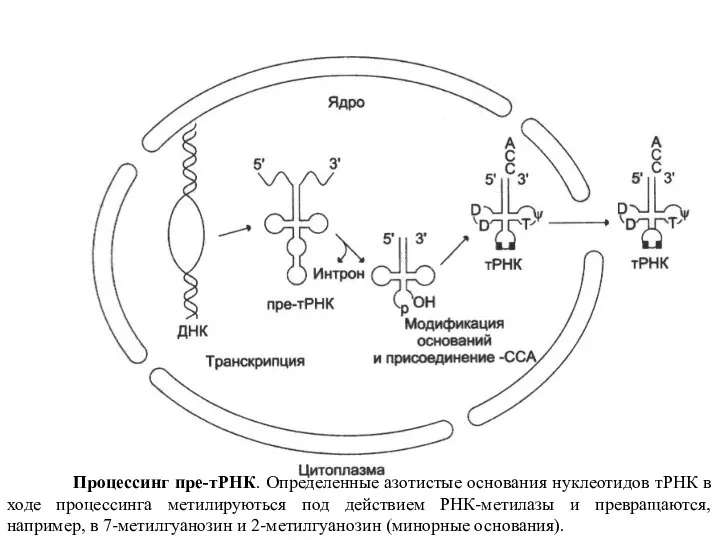
Слайд 23 В процессинге участвует целый ряд ферментов. Например, с помощью нуклеаз вырезаются
В процессинге участвует целый ряд ферментов. Например, с помощью нуклеаз вырезаются

Слайд 24 Существует также альтернативный процессинг, в результате которого из одной и той
Существует также альтернативный процессинг, в результате которого из одной и той

Слайд 25 Альтернативный сплайсинг гена кальцитонина. В клетках щитовидной железы сплайсинг первичного транскрипта
Альтернативный сплайсинг гена кальцитонина. В клетках щитовидной железы сплайсинг первичного транскрипта

Слайд 26 Итак, процессинг включает в себя несколько важных процессов: а) кэппирование РНК,
Итак, процессинг включает в себя несколько важных процессов: а) кэппирование РНК,

Слайд 27Трансляция. Синтез белков
Трансляция. Синтез белков

Слайд 28 Общая характеристика процесса
Информация, содержащаяся в ДНК, передается синтезируемому белку через РНК.
Общая характеристика процесса
Информация, содержащаяся в ДНК, передается синтезируемому белку через РНК.

Слайд 29 После «созревания» мРНК (процессинг) переносится в цитоплазму. Кэппированный конец мРНК, распознается малой
После «созревания» мРНК (процессинг) переносится в цитоплазму. Кэппированный конец мРНК, распознается малой

Слайд 30Рибосома эукариот. А – рибосома 8ОS состоит из большой и малой субъединицы.
Рибосома эукариот. А – рибосома 8ОS состоит из большой и малой субъединицы.
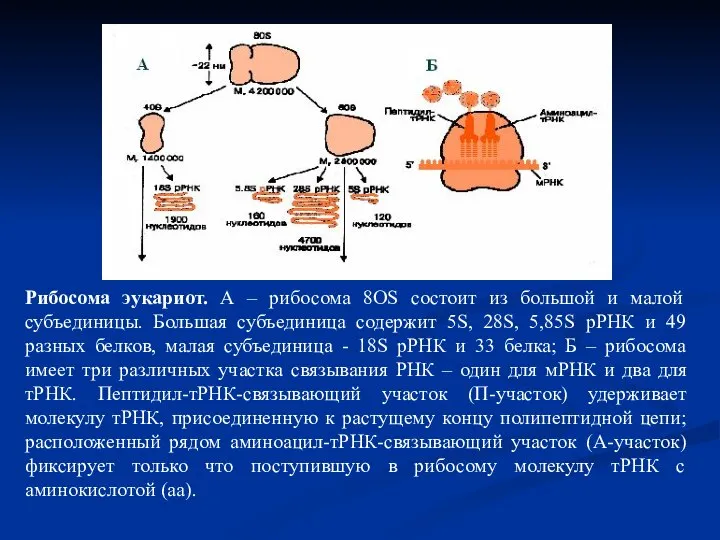
Слайд 31 Рибосомы – это органеллы клеток, обеспечивающие синтез полипептидов. Они объединяют в
Рибосомы – это органеллы клеток, обеспечивающие синтез полипептидов. Они объединяют в

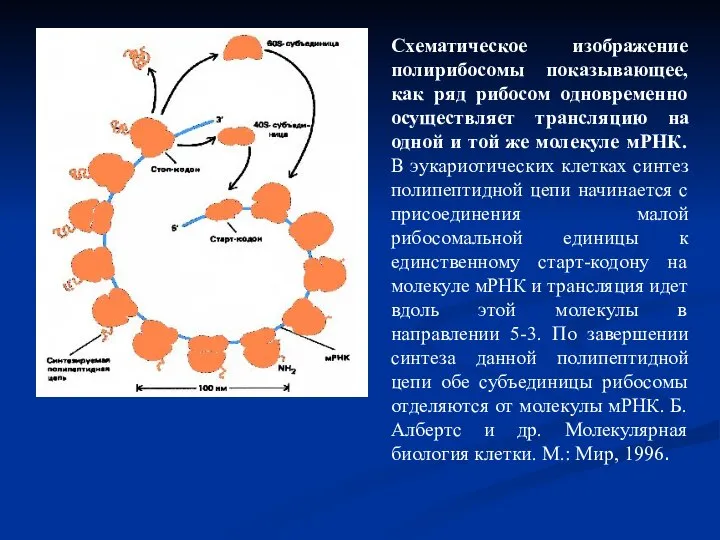
Слайд 32Схематическое изображение полирибосомы показывающее, как ряд рибосом одновременно осуществляет трансляцию на одной
Схематическое изображение полирибосомы показывающее, как ряд рибосом одновременно осуществляет трансляцию на одной
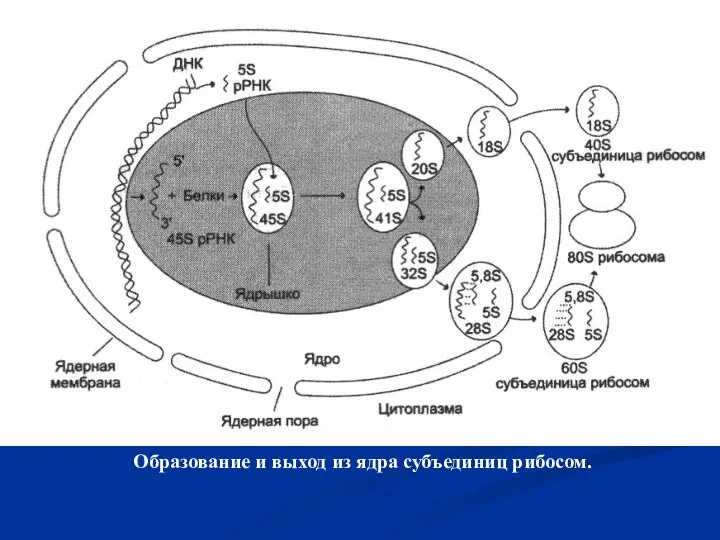
Слайд 33 Образование и выход из ядра субъединиц рибосом.
Образование и выход из ядра субъединиц рибосом.
Слайд 34 С цепью мРНК одновременно может связаться много рибосом. Так образуются полирибосомы
С цепью мРНК одновременно может связаться много рибосом. Так образуются полирибосомы

Слайд 35 Модель молекулярного механизма работы рибосом была предложена А. Спириным в 1968
Модель молекулярного механизма работы рибосом была предложена А. Спириным в 1968

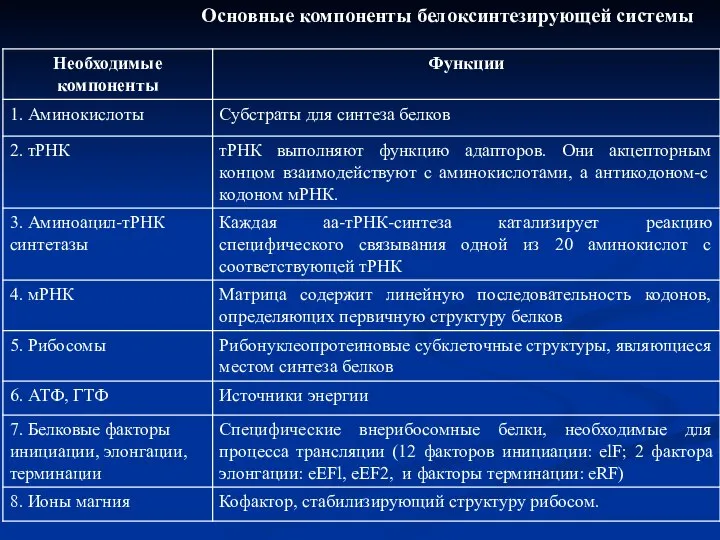
Слайд 36Основные компоненты белоксинтезирующей системы
Основные компоненты белоксинтезирующей системы
Слайд 37 Инициация синтеза белков.
Активация аминокислот. Аминокислоты (АК) в цитозоле клетки вступают
Инициация синтеза белков.
Активация аминокислот. Аминокислоты (АК) в цитозоле клетки вступают

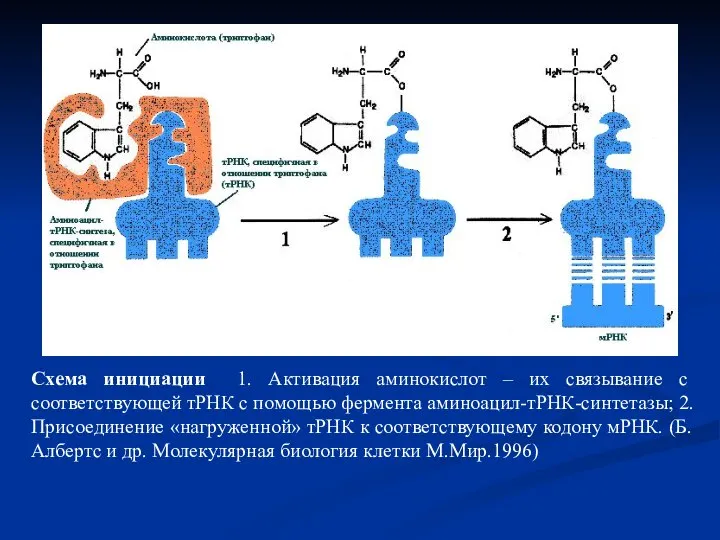
Слайд 38Схема инициации 1. Активация аминокислот – их связывание с соответствующей тРНК с
Слайд 39Активация рибосом, образование белок-синтезирующего комплекса и начало синтеза полипептидной цепи. А –
Активация рибосом, образование белок-синтезирующего комплекса и начало синтеза полипептидной цепи. А –
Слайд 40Комплементарность
– строгое соответствие азотистых оснований одной цепи ДНК другой (А –
Комплементарность
– строгое соответствие азотистых оснований одной цепи ДНК другой (А –

Слайд 41Генетический код
– строгая последовательность триплетов (кодонов) в ДНК и м-РНК, контролирует
Генетический код
– строгая последовательность триплетов (кодонов) в ДНК и м-РНК, контролирует

Слайд 42Характеристика генетического кода
1. Триплетность.
2. Специфичность.
3. Неперекрываемость.
4. Универсальность.
5. Избыточность.
6. Колинеарность.
7. Однонаправленность.
8. Наличие
Характеристика генетического кода
1. Триплетность.
2. Специфичность.
3. Неперекрываемость.
4. Универсальность.
5. Избыточность.
6. Колинеарность.
7. Однонаправленность.
8. Наличие

Слайд 43Образование иницирующего комплекса в ходе синтеза белка у эукариотов.
Образование иницирующего комплекса в ходе синтеза белка у эукариотов.
Слайд 44 Активация рибосом и начало синтеза полипептидной цепи.
Цепь мРНК соединяется
Активация рибосом и начало синтеза полипептидной цепи.
Цепь мРНК соединяется

Слайд 47 Элонгация полипептидной цепи
Удлинение полипептида. Вторая нагруженная, например пролином, тРНК соединяется
Элонгация полипептидной цепи
Удлинение полипептида. Вторая нагруженная, например пролином, тРНК соединяется

Слайд 48 тРНК-дипептидный комплекс на участке А рибосомы вместе с мРНК перемещается относительно
тРНК-дипептидный комплекс на участке А рибосомы вместе с мРНК перемещается относительно

Слайд 49Механизм элонгации полипептидной цепи (объяснение в тексте).
Механизм элонгации полипептидной цепи (объяснение в тексте).
Слайд 50 Весь процесс, включающий поступление тРНК-аминокислотного комплекса, образования пептидной связи и транслокации,
Весь процесс, включающий поступление тРНК-аминокислотного комплекса, образования пептидной связи и транслокации,

Слайд 52 Терминация синтеза белков.
Окончание синтеза и высвобождение полипептидной цепи. В конце
Терминация синтеза белков.
Окончание синтеза и высвобождение полипептидной цепи. В конце

Слайд 53 Посттрансляционная модификация полипептидной цепи белка («созревание»)
Синтезированый из аминокислот полипептид -
Посттрансляционная модификация полипептидной цепи белка («созревание»)
Синтезированый из аминокислот полипептид -

Слайд 55 Значение синтеза белков
Синтез белков является основным процессом в жизни клетки
Значение синтеза белков
Синтез белков является основным процессом в жизни клетки

Слайд 56 Основная догма молекулярной биологии
Хромосомные ДНК содержат полную информацию обо всех
Основная догма молекулярной биологии
Хромосомные ДНК содержат полную информацию обо всех

 Лабораторная работа. Внешнее строение моллюсков
Лабораторная работа. Внешнее строение моллюсков Презентация на тему Морские рыбы
Презентация на тему Морские рыбы  Tsns
Tsns Ромашка аптечная (ромашка лекарственная) Matricaria charnomilla L. (M. recutita L.)
Ромашка аптечная (ромашка лекарственная) Matricaria charnomilla L. (M. recutita L.) Elasipodida. Царство Животные. Подцарство Многоклеточные. Тип Иглокожие
Elasipodida. Царство Животные. Подцарство Многоклеточные. Тип Иглокожие Морковь - овощ
Морковь - овощ Многообразие Простейших. Одноклеточные. 4
Многообразие Простейших. Одноклеточные. 4 Фибриллярные белки
Фибриллярные белки Подъязычный нерв (XII пара)
Подъязычный нерв (XII пара) Химия жизни
Химия жизни Биоэлементы для животных и людей
Биоэлементы для животных и людей Бразильский странствующий паук
Бразильский странствующий паук Класс Земноводные
Класс Земноводные Класс Пресмыкающиеся, или Рептилии
Класс Пресмыкающиеся, или Рептилии Изучение растений в курсе естествознания начальной школы
Изучение растений в курсе естествознания начальной школы Презентация на тему Кто такие звери (1 класс)
Презентация на тему Кто такие звери (1 класс)  Мочевыделительная система
Мочевыделительная система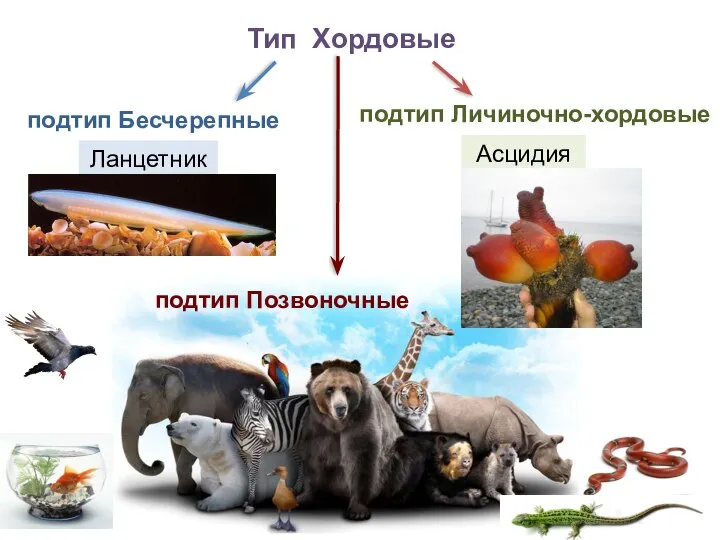 Рыбы. Внутренний костный скелет
Рыбы. Внутренний костный скелет Яблоко. Самый полезный и любимый мой фрукт
Яблоко. Самый полезный и любимый мой фрукт Тип Кольчатые черви Annellida
Тип Кольчатые черви Annellida Хохлатый жаворонок - птица Красной книги
Хохлатый жаворонок - птица Красной книги Дисахариды. Физические свойства
Дисахариды. Физические свойства Сердечно-сосудистая система. Лекция 8
Сердечно-сосудистая система. Лекция 8 Строение и функции нервной системы. Принципы изучения механизмов ее деятельности
Строение и функции нервной системы. Принципы изучения механизмов ее деятельности Большая подборка по второй части. 22 задание
Большая подборка по второй части. 22 задание Возрастные особенности сердечно-сосудистой системы
Возрастные особенности сердечно-сосудистой системы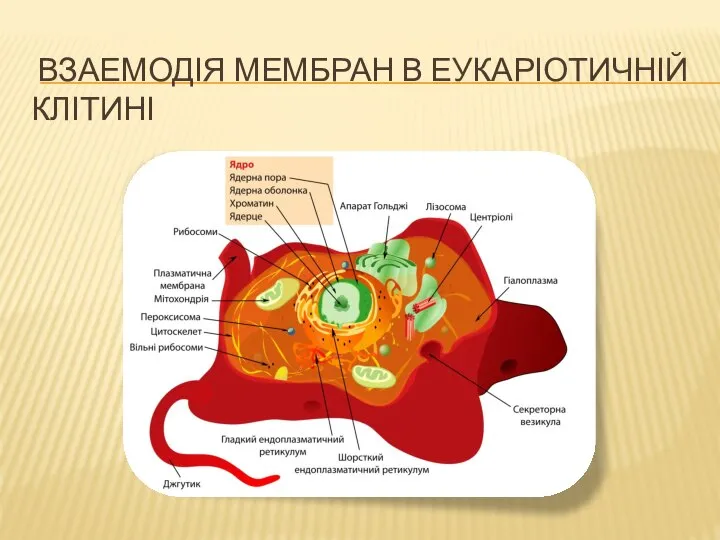 Взаемодія мембран в еукаріотичній клітині
Взаемодія мембран в еукаріотичній клітині Размножение растений
Размножение растений