- Фотосинтез

Содержание
- 2. ФОТОСИНТЕЗ – образование органических веществ зелеными растениями и некоторыми бактериями с использованием энергии солнечного света. Основной
- 3. Джозеф Пристли В 1771 году открыл фотосинтез В 1778 году доказал, что при фотосинтезе растения поглощают
- 4. Вильгельм Пфеффер в 1877 году предложил термин «фотосинтез» Пьер Жозеф Пеллетье Жозеф Бьенеме Каванту Термин «хлорофилл»
- 5. Фотосинтез
- 6. Фотосинтетические пигменты Хлорофиллы Каратиноиды Ксантофиллы Пигменты зеленого листа: • хлорофилл а • хлорофилл b • β-каротин
- 7. Хлорофилл Единственная молекула, которая может поглощать солнечный свет и трансформировать его в энергию электронов, а также
- 8. Фотосинтетические пигменты Хлорофиллы Содержат катион Mg2+ Тетрапиррольное кольцо Хлорофилл a Входит в реакционные центры обеих фотосистем
- 9. Фотосинтетические пигменты Хлорофилл b Входит в светособирающие комплексы обеих фотосистем Спектры поглощения хлорофиллов а и b.
- 10. Различия между хлорофиллами Хл. а Хл. с2 Хл. d Бхл. а Бхл. с Хл. с1 2
- 11. Возбуждение хлорофилла Красный свет – наиболее эффективен для фотосинтеза, т.к. не перевозбуждает молекулы хлорофилла При поглощении
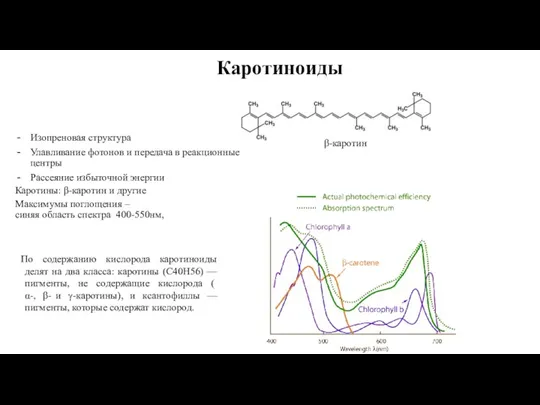
- 12. Каротиноиды Изопреновая структура Улавливание фотонов и передача в реакционные центры Рассеяние избыточной энергии Каротины: β-каротин и
- 13. Фотосинтетические пигменты Ксантофиллы: зеаксантин, антераксантин, виолаксантин → виолаксантиновый цикл (защита от переосвещения)
- 14. Фотовыцветание
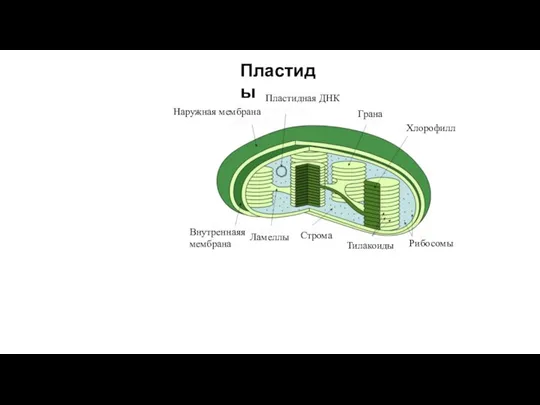
- 15. Наружная мембрана Внутреннаяя мембрана Ламеллы Строма Тилакоиды Рибосомы Пластидная ДНК Грана Хлорофилл Пластиды
- 16. Хлоропласты большинства растений способны перемещаться в клетке в зависимости от интенсивности освещения и его направления. Сильный
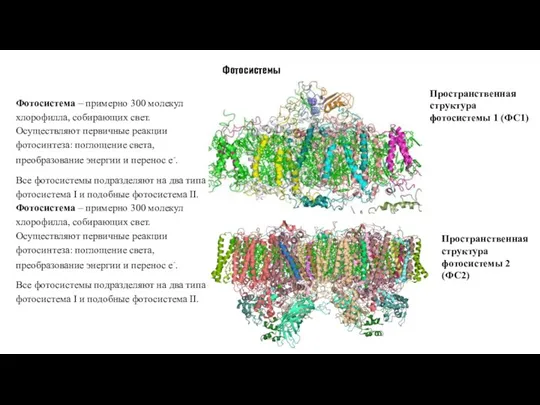
- 17. Фотосистемы Фотосистема – примерно 300 молекул хлорофилла, собирающих свет. Осуществляют первичные реакции фотосинтеза: поглощение света, преобразование
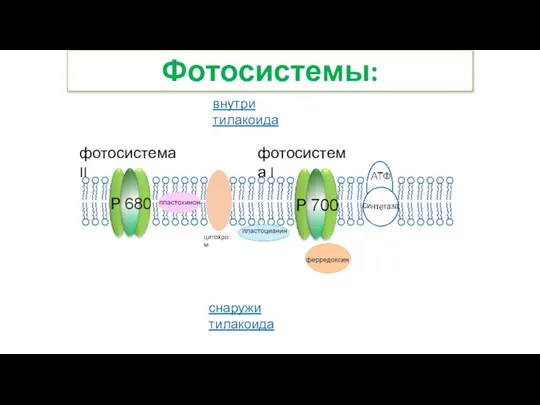
- 18. внутри тилакоида снаружи тилакоида цитохром фотосистема II фотосистема I Фотосистемы:
- 19. Фотосистема I. Фотосистема I высших растений и водорослей представляет собой интегральный пигмент-белковый комплекс молекулярной массой около
- 20. Фотосистема II. Реакционный центр фотосистемы II включает первичный донор электронов — димер хлорофилла а с максимумом
- 21. ФАЗЫ ФОТОСИНТЕЗА СВЕТОВАЯ ФАЗА. Происходит на свету в тилакоидах. Световая фаза – этап фотосинтеза, в течение
- 22. Световая фаза фотосинтеза Хлорофилл поглощает энергию солнечного света (hν), которая используется для: -синтеза молекул АТФ из
- 23. Световая фаза: 1. Молекула хлорофилла фотосистемы I поглощает квант света и переходит в возбужденное состояние. При
- 24. Световая фаза: снаружи тилакоида внутри тилакоида
- 25. Световая фаза: 3. Квант красного света, поглощенный хлорофиллом П680 фотосистемы ІІ, переводит электрон в возбужденное состояние
- 26. Световая фаза: АТФ
- 27. Световая фаза: 5. Электрон поступает в фотосистему I и восстанавливает молекулу П700. При этом молекула П70О
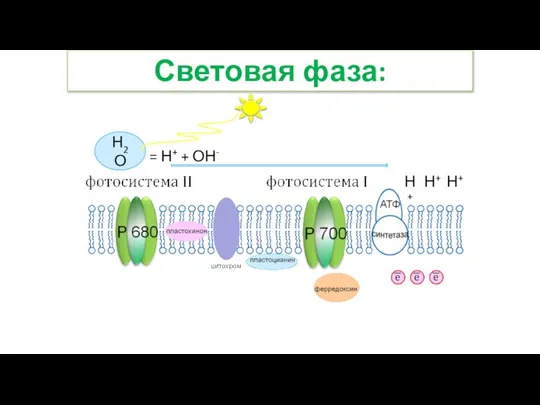
- 28. Световая фаза: Н2О = Н+ + ОН- Н+ Н+ Н+
- 29. Световая фаза: 7. Протоны водорода накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя поверхность мембраны заряжается
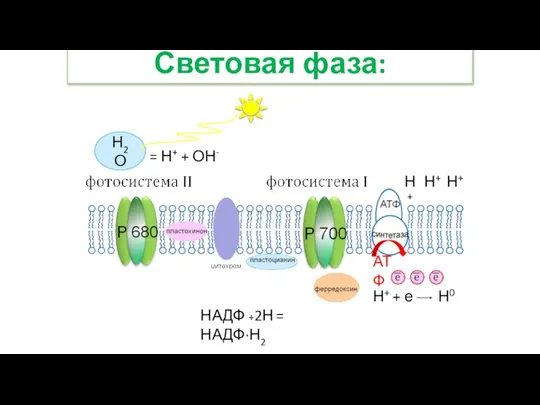
- 30. Световая фаза: Н2О = Н+ + ОН- Н+ Н+ Н+ АТФ НАДФ +2Н = НАДФ·Н2 Н+
- 31. Световая фаза: 9. Катионы водорода на наружной стороне мембраны присоединяют электроны молекулы хлорофилла, образуя атомарный водород,
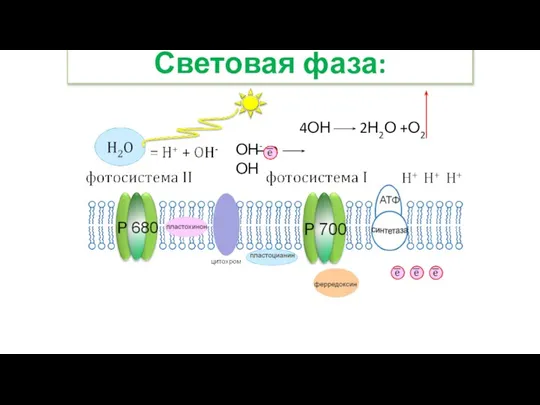
- 32. Световая фаза: ОН- е ОН 4ОН 2Н2О +О2
- 33. Световая фаза: Ионы гидроксильной группы отдают свои электроны, превращаясь в радикалы: ОН- е ОН. Этот электрон
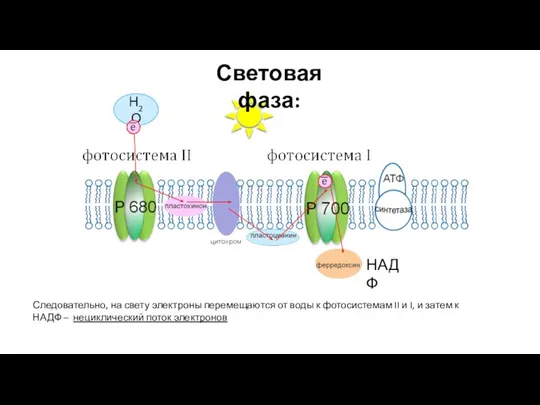
- 34. Н2О НАДФ Следовательно, на свету электроны перемещаются от воды к фотосистемам II и I, и затем
- 35. Световая фаза: Таким образом, энергия солнечного света порождает три процесса: 1) Образование кислорода вследствие фотолиза воды
- 36. Цепь переноса электронов в ФСII В реакционном центре – 2 молекулы хл a (P680) Электрон с
- 37. Фотолиз воды В ФСII должен проходить процесс регенерации пула электронов. Окисленный хл a отнимает электрон у
- 38. Общий план Z-схемы (линейный транспорт e-) Последовательность расположения отдельных элементов электронтранспортной цепи (ЭТЦ) в тилакоидных мембранах
- 39. Варианты транспорта электронов: • нециклический (основной путь); • циклический у ФС I; • циклический транспорт у
- 40. Циклический транспорт Циклический транспорт – образуется только АТФ, NADPH расходуется Циклический поток электронов в фотосистеме II
- 41. Циклическиe потоки электронов вокруг фотосистемы II
- 42. Реакция Меллера (псевдоциклический транспорт e-) высокий восстановительный потенциал, генерируемый на акцепторной стороне фотосистемы I, расходуется не
- 43. Циклический транспорт электронов вокруг фотосистемы I и хлоропластное дыхание НАД(Ф)Н Дегид- роге- наза АО О2 →Н2О
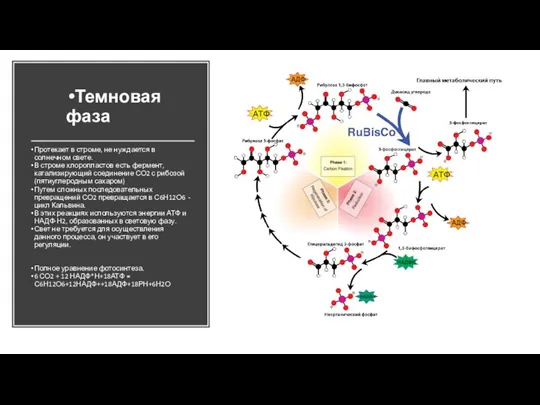
- 44. Протекает в строме, не нуждается в солнечном свете. В строме хлоропластов есть фермент, катализирующий соединение СО2
- 45. цикла Кальвина — в честь его первооткрывателя, лорда Мелвина Кальвина, который получил за это открытие Нобелевскую
- 46. RuBisCO Состоит из 8 димеров больших и малых субъединиц Класс: лиазы ДНК больших цепей – в
- 47. Темновая фаза фотосинтеза (цикл Кальвина) Продукты световой фазы: АТФ и NADPH Цикл Кальвина присутствует у всех
- 48. Темновая фаза фотосинтеза (цикл Кальвина) Второй этап цикла – восстановление 3-фосфоглицерата 2 реакции: фосфорилирование (АТФ) и
- 49. Темновая фаза фотосинтеза (цикл Кальвина)
- 50. Темновая фаза фотосинтеза (цикл Кальвина) Энергетический расчет на 1 молекулу глюкозы (6 оборотов цикла): 12х 3-фосфоглицерат→12х
- 51. Аноксигенный фотосинтез (англ. anoxygenic «бескислородный») — вариант фотосинтеза (процесса образования органических веществ на свету), при котором
- 52. 5 млрд-3млрд лет назад анаэробный этап жизни. 3 млрд лет назад на Земле возникают первые сине-зеленые
- 53. Динофлагелляты, или динофитовые водоросли Кокколитофориды Диатомовые водоросли
- 54. Эндосимбиоз в эволюции водорослей. Можно представить, что когда-то эукариот пытался съесть цианобактерию, но передумал и решил
- 56. Elysia viridis Elysia chlorotica
- 57. Восточный шершень (Vespa orientalis) преобразует солнечный свет в электроэнергию с помощью пигмента ксантоптерина
- 58. Поперечное сечение листа арабидопсиса — типичного С3-растения. Хорошо видно строение сосудистых пучков. С3-растения, как правило, процветают
- 59. С4-фотосинтез Используют около 7600 видов растений – 3% всех наземных видов растений. Механизм концентрации углерода: Температура
- 60. C4-фотосинтез Характерен для растений жаркого и сухого климата, необходим для более эффективного поглощения CO2 из атмосферы
- 61. NADPH-зависимый C4-фотосинтез Представители: кукуруза, сахарный тростник В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат + NADPH
- 62. NADPH-зависимый C4-фотосинтез
- 63. NAD-зависимый C4-фотосинтез Представители: просо В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат → аспартат В клетках
- 65. ФЕП-карбоксилазный C4-фотосинтез Представители: тропические злаковые В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат → аспартат В
- 67. САМ-фотосинтез Назван в честь семейства Crassulaceae - Толстянковые Crassulaceae acid metabolism — кислотный метаболизм толстянковых Зафиксирован
- 68. CAM-метаболизм Разделение поглощения и использования CO2 по времени Примеры: толстянковые, полушники Ночью устьица открыты, и CO2
- 69. CAM-метаболизм. Ночная фаза
- 70. CAM-метаболизм. Дневная фаза
- 71. Регуляция транспирации Интенсивность транспирации регулируется осмотическим давлением замыкающих клеток устьиц При повышении осмотического давления в замыкающих
- 72. Регуляция транспирации
- 73. Синтез сахарозы Глюкоза → глюкозо-6-фосфат + UTP → UDP-глюкоза UDP-глюкоза + фруктозо-6-фосфат (из гликолиза) → сахарозо-6-фосфат
- 74. Синтез сахарозы
- 75. Синтез крахмала Фруктозо-6-фосфат (из цикла Кальвина) → глюкозо-6-фосфат → глюкозо-1-фосфат + ATP → ADP-Glu → крахмал
- 76. Синтез крахмала
- 77. Синтез целлюлозы Сахароза→УДФ-глюкоза→целлюлоза Целлюлозосинтаза – розеточный комплекс, синтезирует сразу микрофибриллу целлюлозы (6 цепей)
- 78. Фотодыхание Роль: утилизация фосфогликолата, образующегося в ходе оксигеназной активности RubisCO Суммарное уравнение: 2 (2-фосфогликолат) → 3-фосфоглицерат
- 79. Фотодыхание Реакции в хлоропласте: 1,5-рибулозобисфосфат + O2 → 3-фосфоглицерат + 2-фосфогликолат 2-фосфогликолат → гликолат Обменник: 2
- 80. Фотодыхание Реакции в пероксисоме: 2 гликолат + 2O2 → 2 глиоксилат + 2H2O2 2 глиоксилат +
- 82. Фотодыхание Энергетика реакции: 2х рибулозо-1,5-бисфосфат → 2х 3-фосфоглицерат + 2х 2-фосфогликолат (самопроизвольно) 2х 2-фосфогликолат → 3-фосфоглицерат
- 83. ВТОРИЧНЫЕ МЕТАБОЛИТЫ
- 84. Различают первичные продукты обмена веществ клетки и вторичные, которые образуются из первичных в ходе б/х превращений
- 87. Существует несколько классификаций веществ вторичного происхождения. 1. ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ 3 Класса: 1. Фенольные соединения 2. Алкалоиды
- 88. 2. Классификация по функциям:
- 89. 3. Физиологически активные вещества: фитогормоны и витамины. 4. Вещества с неизвестным физиологическим значением: гликозиды, алкалоиды, таннины,
- 90. 1. ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ – вещества ароматической природы, содержащие одну (фенолы) или более (полифенолы) гидроксильных
- 91. Фенольные соединения накапливаются в разных частях растения в зависимости от вида: цветках (медуница), плодах (малина), корнеплодах
- 92. ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ
- 93. Примеры окраски антоцианами
- 94. Богаты катехинами: виноград, чай
- 95. 3. Полифенолы: лигнины (обеспечивают одревеснение, т.е. опору растению), таннины или дубильные вещества (защищают от животных, т.к.
- 96. Кора и листья дуба богаты таннинами
- 97. 2. АЛКАЛОИДЫ
- 98. Алкалоиды накапливаются в разных частях растения в зависимости от вида: листьях (табака – никотин, чая -
- 99. 3. ИЗОПРЕНОИДЫ
- 100. Представители изопреноидов:
- 102. ИСПОЛЬЗОВАНИЕ ВТОРИЧНЫХ МЕТАБОЛИТОВ
- 105. Эфирномасличные растения
- 106. Обзор класса терпенов и некоторые типичные представители
- 108. Скачать презентацию
Слайд 2ФОТОСИНТЕЗ – образование органических веществ зелеными растениями и некоторыми бактериями с использованием энергии
ФОТОСИНТЕЗ – образование органических веществ зелеными растениями и некоторыми бактериями с использованием энергии

Слайд 3Джозеф Пристли
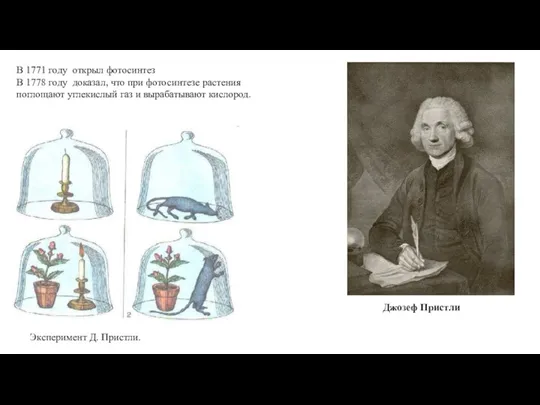
В 1771 году открыл фотосинтез
В 1778 году доказал, что при фотосинтезе растения поглощают углекислый газ и вырабатывают кислород.
Эксперимент
Джозеф Пристли
В 1771 году открыл фотосинтез
В 1778 году доказал, что при фотосинтезе растения поглощают углекислый газ и вырабатывают кислород.
Эксперимент
Слайд 4Вильгельм Пфеффер
в 1877 году предложил термин «фотосинтез»
Пьер Жозеф Пеллетье
Жозеф Бьенеме Каванту
Термин «хлорофилл» был
Вильгельм Пфеффер
в 1877 году предложил термин «фотосинтез»
Пьер Жозеф Пеллетье
Жозеф Бьенеме Каванту
Термин «хлорофилл» был

Слайд 5Фотосинтез
Фотосинтез

Слайд 6Фотосинтетические пигменты
Хлорофиллы
Каратиноиды
Ксантофиллы
Пигменты зеленого листа:
• хлорофилл а
• хлорофилл b
• β-каротин
• ксантофиллы (виолоксантин, лютеин, зеаксантин,
Фотосинтетические пигменты
Хлорофиллы
Каратиноиды
Ксантофиллы
Пигменты зеленого листа:
• хлорофилл а
• хлорофилл b
• β-каротин
• ксантофиллы (виолоксантин, лютеин, зеаксантин,

Слайд 7Хлорофилл
Единственная молекула, которая может поглощать солнечный свет и трансформировать его в энергию
Хлорофилл
Единственная молекула, которая может поглощать солнечный свет и трансформировать его в энергию

Слайд 8Фотосинтетические пигменты
Хлорофиллы
Содержат катион Mg2+
Тетрапиррольное кольцо
Хлорофилл a
Входит в реакционные центры обеих фотосистем
Имеется фитольный
Фотосинтетические пигменты
Хлорофиллы
Содержат катион Mg2+
Тетрапиррольное кольцо
Хлорофилл a
Входит в реакционные центры обеих фотосистем
Имеется фитольный

Слайд 9Фотосинтетические пигменты
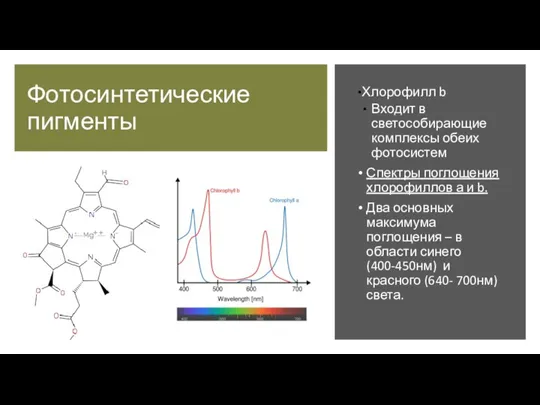
Хлорофилл b
Входит в светособирающие комплексы обеих
фотосистем
Спектры поглощения хлорофиллов а и b.
Два
Фотосинтетические пигменты
Хлорофилл b
Входит в светособирающие комплексы обеих
фотосистем
Спектры поглощения хлорофиллов а и b.
Два
Слайд 10Различия между хлорофиллами
Хл. а
Хл. с2 Хл. d
Бхл. а Бхл. с
Хл. с1
2
Бурые, диатомовые
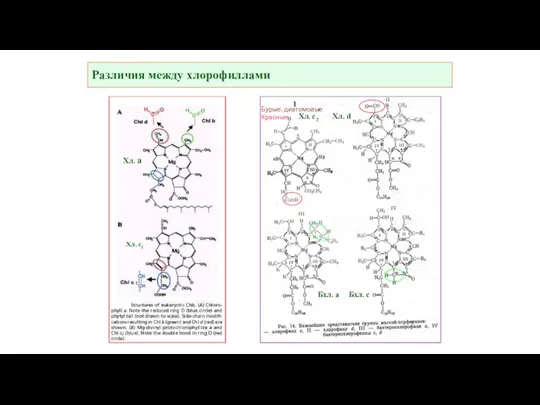
Различия между хлорофиллами
Хл. а
Хл. с2 Хл. d
Бхл. а Бхл. с
Хл. с1
2
Бурые, диатомовые
Слайд 11Возбуждение хлорофилла
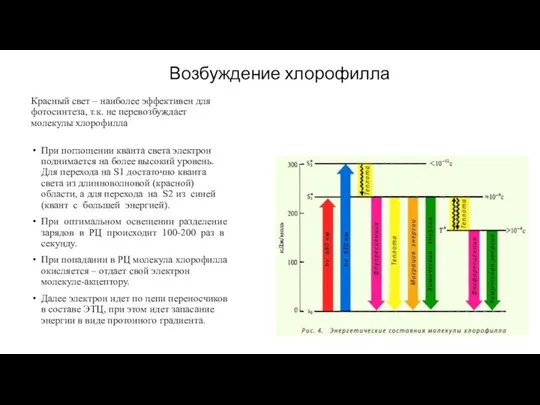
Красный свет – наиболее эффективен для фотосинтеза, т.к. не перевозбуждает молекулы
Возбуждение хлорофилла
Красный свет – наиболее эффективен для фотосинтеза, т.к. не перевозбуждает молекулы
Слайд 12Каротиноиды
Изопреновая структура
Улавливание фотонов и передача в реакционные центры
Рассеяние избыточной энергии
Каротины: β-каротин и
Каротиноиды
Изопреновая структура
Улавливание фотонов и передача в реакционные центры
Рассеяние избыточной энергии
Каротины: β-каротин и
Слайд 13Фотосинтетические пигменты
Ксантофиллы: зеаксантин, антераксантин, виолаксантин →
виолаксантиновый цикл (защита от переосвещения)
Фотосинтетические пигменты
Ксантофиллы: зеаксантин, антераксантин, виолаксантин →
виолаксантиновый цикл (защита от переосвещения)

Слайд 14Фотовыцветание
Фотовыцветание

Слайд 15Наружная мембрана
Внутреннаяя
мембрана
Ламеллы
Строма
Тилакоиды
Рибосомы
Пластидная ДНК
Грана
Хлорофилл
Пластиды
Наружная мембрана
Внутреннаяя
мембрана
Ламеллы
Строма
Тилакоиды
Рибосомы
Пластидная ДНК
Грана
Хлорофилл
Пластиды
Слайд 16Хлоропласты большинства растений способны перемещаться в клетке в зависимости от интенсивности освещения
Хлоропласты большинства растений способны перемещаться в клетке в зависимости от интенсивности освещения

Слайд 17Фотосистемы
Фотосистема – примерно 300 молекул хлорофилла, собирающих свет. Осуществляют первичные реакции фотосинтеза:
Фотосистемы
Фотосистема – примерно 300 молекул хлорофилла, собирающих свет. Осуществляют первичные реакции фотосинтеза:
Слайд 18внутри тилакоида
снаружи тилакоида
цитохром
фотосистема II
фотосистема I
Фотосистемы:
внутри тилакоида
снаружи тилакоида
цитохром
фотосистема II
фотосистема I
Фотосистемы:
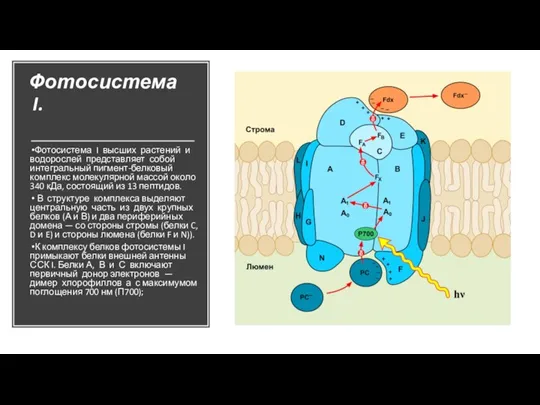
Слайд 19Фотосистема I.
Фотосистема I высших растений и водорослей представляет собой интегральный пигмент-белковый комплекс
Фотосистема I.
Фотосистема I высших растений и водорослей представляет собой интегральный пигмент-белковый комплекс
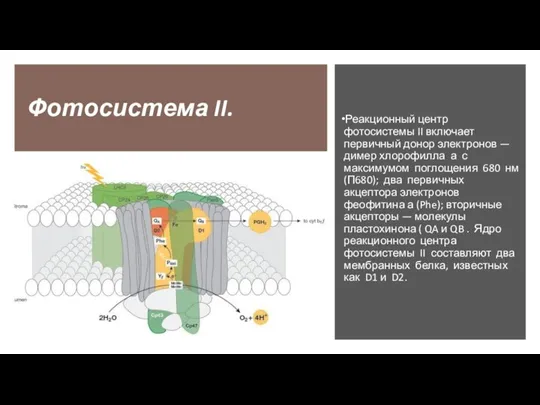
Слайд 20Фотосистема II.
Реакционный центр фотосистемы II включает первичный донор электронов — димер хлорофилла
Фотосистема II.
Реакционный центр фотосистемы II включает первичный донор электронов — димер хлорофилла
Слайд 21ФАЗЫ ФОТОСИНТЕЗА
СВЕТОВАЯ ФАЗА.
Происходит на свету в тилакоидах.
Световая фаза – этап фотосинтеза,
ФАЗЫ ФОТОСИНТЕЗА
СВЕТОВАЯ ФАЗА.
Происходит на свету в тилакоидах.
Световая фаза – этап фотосинтеза,

Слайд 22Световая фаза фотосинтеза
Хлорофилл поглощает энергию солнечного света (hν), которая используется для:
-синтеза молекул
Световая фаза фотосинтеза
Хлорофилл поглощает энергию солнечного света (hν), которая используется для:
-синтеза молекул

Слайд 23Световая фаза:
1. Молекула хлорофилла фотосистемы I поглощает квант света и переходит в
Световая фаза:
1. Молекула хлорофилла фотосистемы I поглощает квант света и переходит в

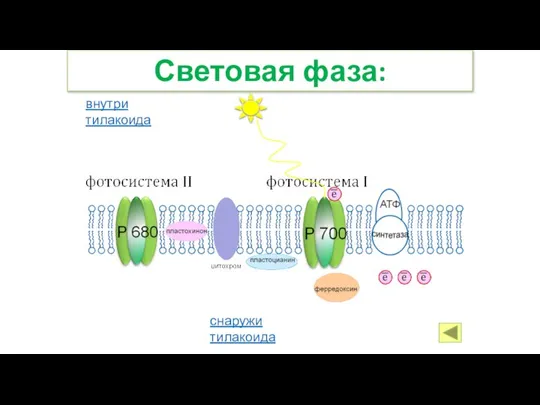
Слайд 24Световая фаза:
снаружи тилакоида
внутри тилакоида
Световая фаза:
снаружи тилакоида
внутри тилакоида
Слайд 25Световая фаза:
3. Квант красного света, поглощенный хлорофиллом П680 фотосистемы ІІ, переводит электрон
Световая фаза:
3. Квант красного света, поглощенный хлорофиллом П680 фотосистемы ІІ, переводит электрон

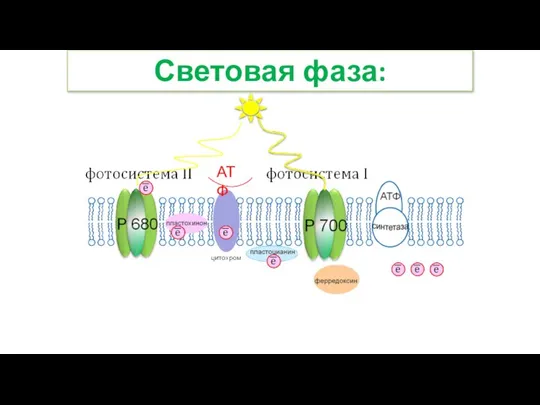
Слайд 26Световая фаза:
АТФ
Световая фаза:
АТФ
Слайд 27Световая фаза:
5. Электрон поступает в фотосистему I и восстанавливает молекулу П700. При
Световая фаза:
5. Электрон поступает в фотосистему I и восстанавливает молекулу П700. При

Слайд 28Световая фаза:
Н2О
= Н+ + ОН-
Н+
Н+
Н+
Световая фаза:
Н2О
= Н+ + ОН-
Н+
Н+
Н+
Слайд 29Световая фаза:
7. Протоны водорода накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя
Световая фаза:
7. Протоны водорода накапливаются внутри тилакоида, создавая Н+-резервуар. В результате внутренняя

Слайд 30Световая фаза:
Н2О
= Н+ + ОН-
Н+
Н+
Н+
АТФ
НАДФ +2Н = НАДФ·Н2
Н+ + е
Световая фаза:
Н2О
= Н+ + ОН-
Н+
Н+
Н+
АТФ
НАДФ +2Н = НАДФ·Н2
Н+ + е
Слайд 31Световая фаза:
9. Катионы водорода на наружной стороне мембраны присоединяют электроны молекулы хлорофилла,
Световая фаза:
9. Катионы водорода на наружной стороне мембраны присоединяют электроны молекулы хлорофилла,

Слайд 32Световая фаза:
ОН- е ОН
4ОН 2Н2О +О2
Световая фаза:
ОН- е ОН
4ОН 2Н2О +О2
Слайд 33Световая фаза:
Ионы гидроксильной группы отдают свои электроны, превращаясь в радикалы:
ОН- е
Световая фаза:
Ионы гидроксильной группы отдают свои электроны, превращаясь в радикалы:
ОН- е

Слайд 34Н2О
НАДФ
Следовательно, на свету электроны перемещаются от воды к фотосистемам II и I,
Н2О
НАДФ
Следовательно, на свету электроны перемещаются от воды к фотосистемам II и I,
Слайд 35Световая фаза:
Таким образом, энергия солнечного света порождает три процесса:
1) Образование кислорода
Световая фаза:
Таким образом, энергия солнечного света порождает три процесса:
1) Образование кислорода

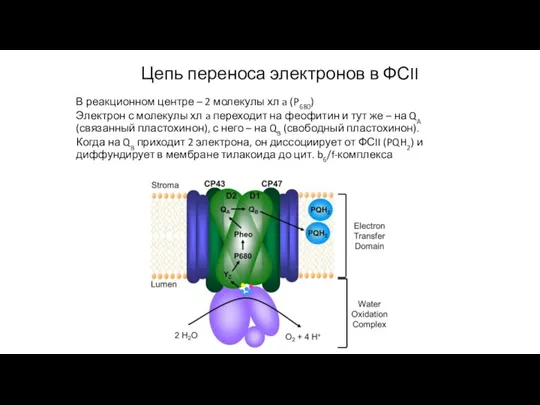
Слайд 36Цепь переноса электронов в ФСII
В реакционном центре – 2 молекулы хл a
Цепь переноса электронов в ФСII
В реакционном центре – 2 молекулы хл a
Слайд 37Фотолиз воды
В ФСII должен проходить процесс регенерации пула электронов.
Окисленный хл a отнимает
Фотолиз воды
В ФСII должен проходить процесс регенерации пула электронов.
Окисленный хл a отнимает

Слайд 38Общий план Z-схемы (линейный транспорт e-)
Последовательность расположения отдельных элементов электронтранспортной цепи (ЭТЦ)
Общий план Z-схемы (линейный транспорт e-)
Последовательность расположения отдельных элементов электронтранспортной цепи (ЭТЦ)

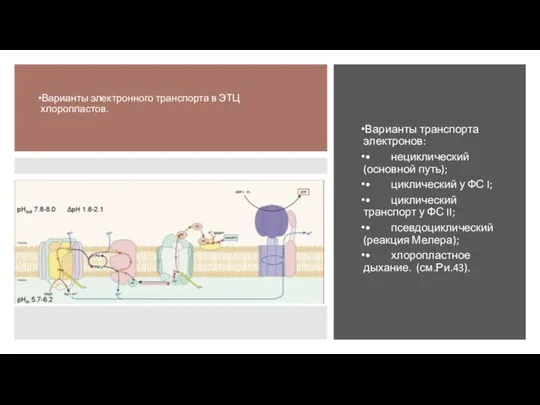
Слайд 39Варианты транспорта электронов:
• нециклический (основной путь);
• циклический у ФС I;
• циклический транспорт
Варианты транспорта электронов:
• нециклический (основной путь);
• циклический у ФС I;
• циклический транспорт
Слайд 40Циклический транспорт
Циклический транспорт – образуется только АТФ, NADPH расходуется
Циклический поток электронов в
Циклический транспорт
Циклический транспорт – образуется только АТФ, NADPH расходуется
Циклический поток электронов в

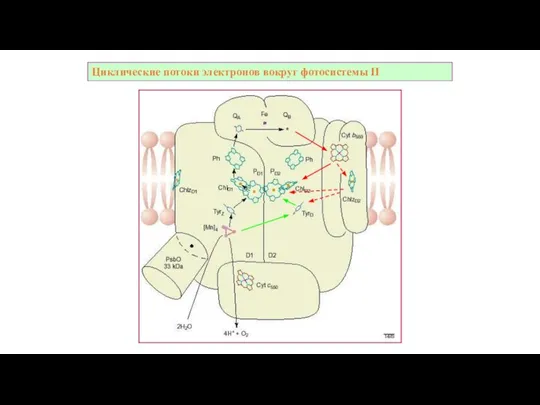
Слайд 41Циклическиe потоки электронов вокруг фотосистемы II
Циклическиe потоки электронов вокруг фотосистемы II
Слайд 42Реакция Меллера (псевдоциклический транспорт e-)
высокий восстановительный потенциал, генерируемый на акцепторной стороне фотосистемы
Реакция Меллера (псевдоциклический транспорт e-)
высокий восстановительный потенциал, генерируемый на акцепторной стороне фотосистемы

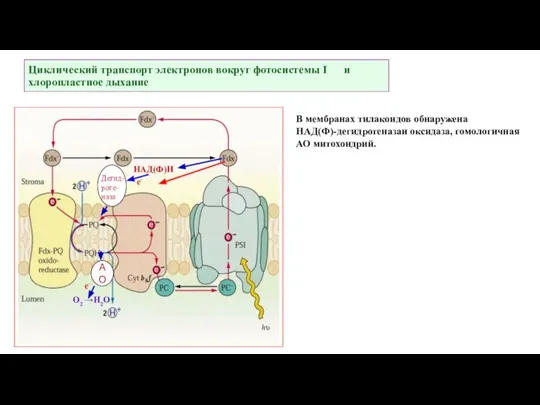
Слайд 43Циклический транспорт электронов вокруг фотосистемы I и хлоропластное дыхание
НАД(Ф)Н
Дегид-
роге-
наза
АО
О2 →Н2О
е-
е-
В мембранах тилакоидов
Циклический транспорт электронов вокруг фотосистемы I и хлоропластное дыхание
НАД(Ф)Н
Дегид-
роге-
наза
АО
О2 →Н2О
е-
е-
В мембранах тилакоидов
Слайд 44Протекает в строме, не нуждается в солнечном свете.
В строме хлоропластов есть фермент,
Протекает в строме, не нуждается в солнечном свете.
В строме хлоропластов есть фермент,
Слайд 45цикла Кальвина — в честь его первооткрывателя, лорда Мелвина Кальвина, который получил
цикла Кальвина — в честь его первооткрывателя, лорда Мелвина Кальвина, который получил

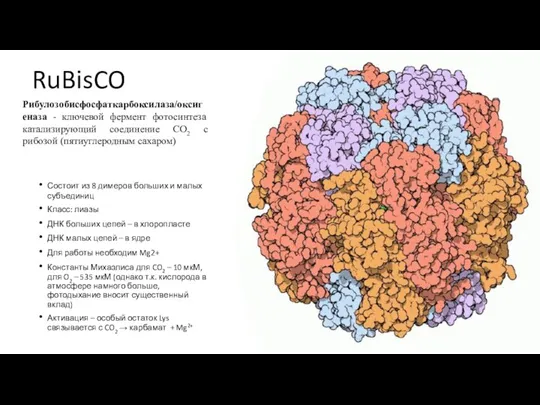
Слайд 46RuBisCO
Состоит из 8 димеров больших и малых субъединиц
Класс: лиазы
ДНК больших цепей –
RuBisCO
Состоит из 8 димеров больших и малых субъединиц
Класс: лиазы
ДНК больших цепей –
Слайд 47Темновая фаза фотосинтеза (цикл Кальвина)
Продукты световой фазы: АТФ и NADPH
Цикл Кальвина присутствует
Темновая фаза фотосинтеза (цикл Кальвина)
Продукты световой фазы: АТФ и NADPH
Цикл Кальвина присутствует

Слайд 48Темновая фаза фотосинтеза (цикл Кальвина)
Второй этап цикла – восстановление 3-фосфоглицерата
2 реакции: фосфорилирование
Темновая фаза фотосинтеза (цикл Кальвина)
Второй этап цикла – восстановление 3-фосфоглицерата
2 реакции: фосфорилирование

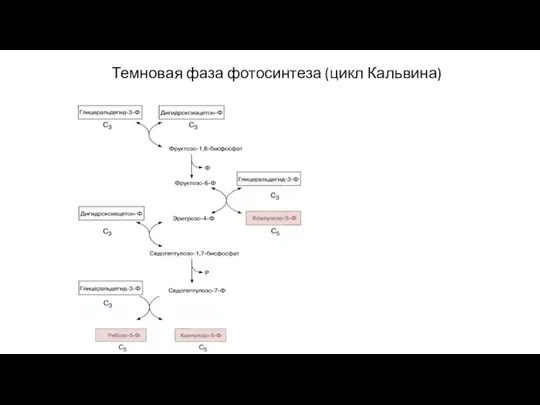
Слайд 49Темновая фаза фотосинтеза (цикл Кальвина)
Темновая фаза фотосинтеза (цикл Кальвина)
Слайд 50Темновая фаза фотосинтеза (цикл Кальвина)
Энергетический расчет на 1 молекулу глюкозы (6 оборотов
Темновая фаза фотосинтеза (цикл Кальвина)
Энергетический расчет на 1 молекулу глюкозы (6 оборотов

Слайд 51Аноксигенный фотосинтез (англ. anoxygenic «бескислородный») — вариант фотосинтеза (процесса образования органических веществ на свету),
Аноксигенный фотосинтез (англ. anoxygenic «бескислородный») — вариант фотосинтеза (процесса образования органических веществ на свету),

Слайд 525 млрд-3млрд лет назад анаэробный этап жизни.
3 млрд лет назад на Земле
5 млрд-3млрд лет назад анаэробный этап жизни.
3 млрд лет назад на Земле

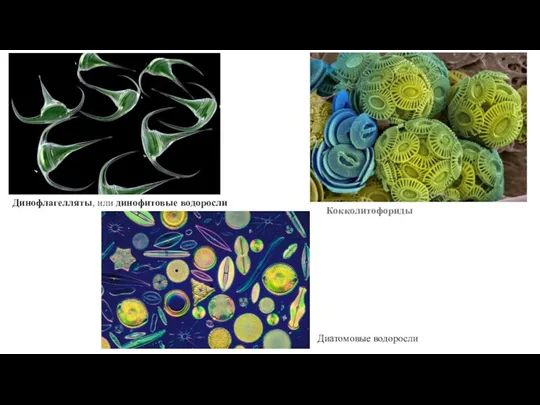
Слайд 53Динофлагелляты, или динофитовые водоросли
Кокколитофориды
Диатомовые водоросли
Динофлагелляты, или динофитовые водоросли
Кокколитофориды
Диатомовые водоросли
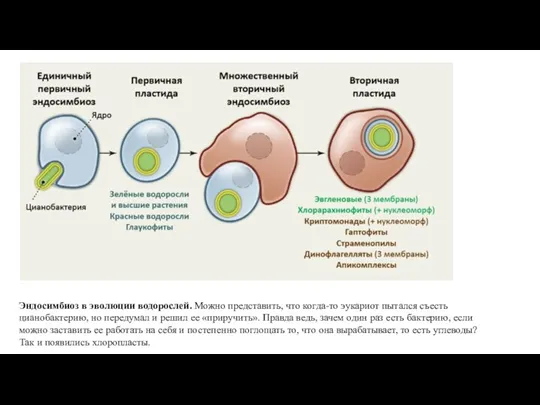
Слайд 54Эндосимбиоз в эволюции водорослей. Можно представить, что когда-то эукариот пытался съесть цианобактерию, но передумал и решил
Эндосимбиоз в эволюции водорослей. Можно представить, что когда-то эукариот пытался съесть цианобактерию, но передумал и решил

Слайд 56Elysia viridis Elysia chlorotica
Elysia viridis Elysia chlorotica

Слайд 57Восточный шершень (Vespa orientalis)
преобразует солнечный свет в электроэнергию с помощью пигмента
Восточный шершень (Vespa orientalis)
преобразует солнечный свет в электроэнергию с помощью пигмента

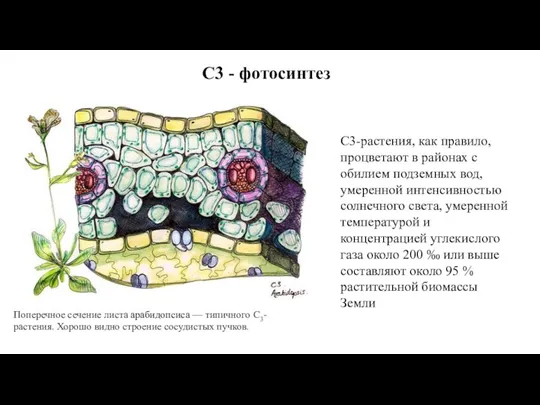
Слайд 58Поперечное сечение листа арабидопсиса — типичного С3-растения. Хорошо видно строение сосудистых пучков.
С3-растения, как правило,
Поперечное сечение листа арабидопсиса — типичного С3-растения. Хорошо видно строение сосудистых пучков.
С3-растения, как правило,
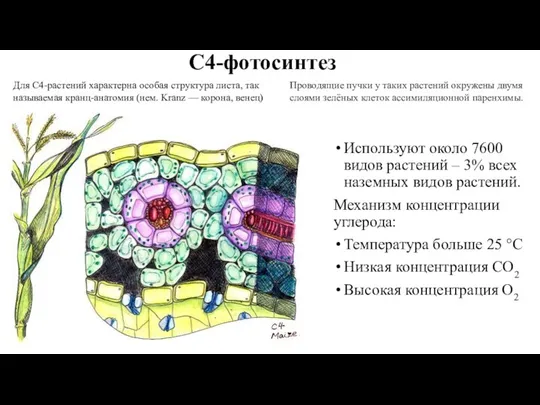
Слайд 59С4-фотосинтез
Используют около 7600 видов растений – 3% всех наземных видов растений.
Механизм концентрации
С4-фотосинтез
Используют около 7600 видов растений – 3% всех наземных видов растений.
Механизм концентрации
Слайд 60C4-фотосинтез
Характерен для растений жаркого и сухого климата, необходим для более эффективного поглощения
C4-фотосинтез
Характерен для растений жаркого и сухого климата, необходим для более эффективного поглощения

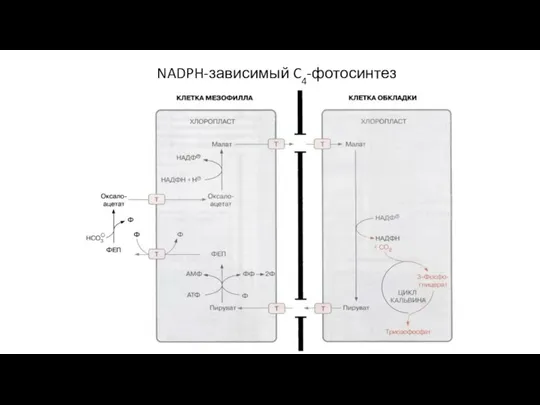
Слайд 61NADPH-зависимый C4-фотосинтез
Представители: кукуруза, сахарный тростник
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат
NADPH-зависимый C4-фотосинтез
Представители: кукуруза, сахарный тростник
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат

Слайд 62NADPH-зависимый C4-фотосинтез
NADPH-зависимый C4-фотосинтез
Слайд 63NAD-зависимый C4-фотосинтез
Представители: просо
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат → аспартат
В
NAD-зависимый C4-фотосинтез
Представители: просо
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат → аспартат
В

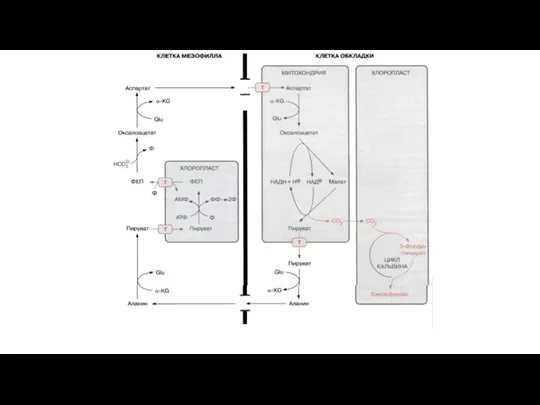
Слайд 65ФЕП-карбоксилазный C4-фотосинтез
Представители: тропические злаковые
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат →
ФЕП-карбоксилазный C4-фотосинтез
Представители: тропические злаковые
В корончатом мезофилле: ФЕП + HCO3- → оксалоацетат →

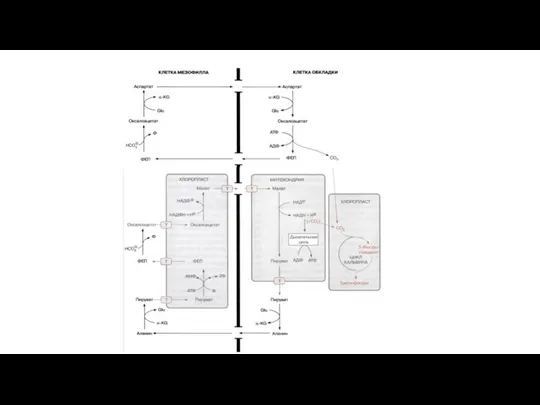
Слайд 67САМ-фотосинтез
Назван в честь семейства Crassulaceae - Толстянковые
Crassulaceae acid metabolism — кислотный метаболизм толстянковых
Зафиксирован
САМ-фотосинтез
Назван в честь семейства Crassulaceae - Толстянковые
Crassulaceae acid metabolism — кислотный метаболизм толстянковых
Зафиксирован

Слайд 68CAM-метаболизм
Разделение поглощения и использования CO2 по времени
Примеры: толстянковые, полушники
Ночью устьица открыты, и
CAM-метаболизм
Разделение поглощения и использования CO2 по времени
Примеры: толстянковые, полушники
Ночью устьица открыты, и

Слайд 69CAM-метаболизм. Ночная фаза
CAM-метаболизм. Ночная фаза

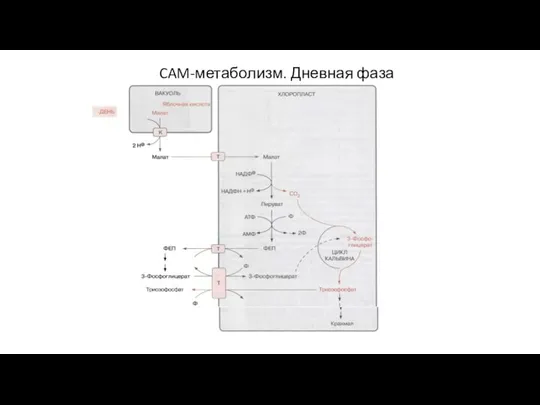
Слайд 70CAM-метаболизм. Дневная фаза
CAM-метаболизм. Дневная фаза
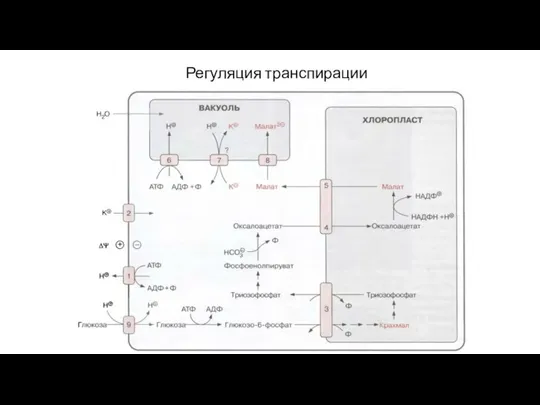
Слайд 71Регуляция транспирации
Интенсивность транспирации регулируется осмотическим давлением замыкающих клеток устьиц
При повышении осмотического давления
Регуляция транспирации
Интенсивность транспирации регулируется осмотическим давлением замыкающих клеток устьиц
При повышении осмотического давления

Слайд 72Регуляция транспирации
Регуляция транспирации
Слайд 73Синтез сахарозы
Глюкоза → глюкозо-6-фосфат + UTP → UDP-глюкоза
UDP-глюкоза + фруктозо-6-фосфат (из гликолиза)
Синтез сахарозы
Глюкоза → глюкозо-6-фосфат + UTP → UDP-глюкоза
UDP-глюкоза + фруктозо-6-фосфат (из гликолиза)

Слайд 74Синтез сахарозы
Синтез сахарозы

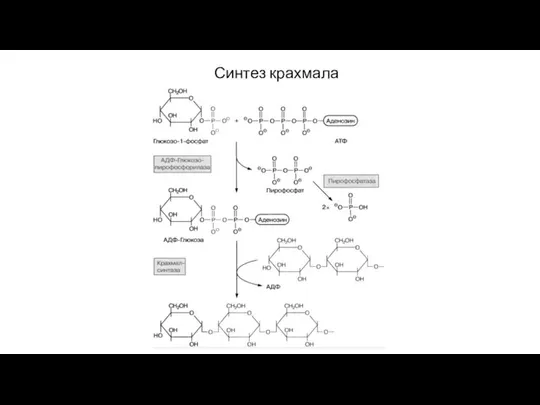
Слайд 75Синтез крахмала
Фруктозо-6-фосфат (из цикла Кальвина) → глюкозо-6-фосфат → глюкозо-1-фосфат + ATP →
Синтез крахмала
Фруктозо-6-фосфат (из цикла Кальвина) → глюкозо-6-фосфат → глюкозо-1-фосфат + ATP →

Слайд 76Синтез крахмала
Синтез крахмала
Слайд 77Синтез целлюлозы
Сахароза→УДФ-глюкоза→целлюлоза
Целлюлозосинтаза – розеточный комплекс, синтезирует сразу микрофибриллу целлюлозы (6 цепей)
Синтез целлюлозы
Сахароза→УДФ-глюкоза→целлюлоза
Целлюлозосинтаза – розеточный комплекс, синтезирует сразу микрофибриллу целлюлозы (6 цепей)

Слайд 78Фотодыхание
Роль: утилизация фосфогликолата, образующегося в ходе оксигеназной активности RubisCO
Суммарное уравнение:
2 (2-фосфогликолат)
Фотодыхание
Роль: утилизация фосфогликолата, образующегося в ходе оксигеназной активности RubisCO
Суммарное уравнение:
2 (2-фосфогликолат)

Слайд 79Фотодыхание
Реакции в хлоропласте:
1,5-рибулозобисфосфат + O2 → 3-фосфоглицерат + 2-фосфогликолат
2-фосфогликолат → гликолат
Обменник: 2
Фотодыхание
Реакции в хлоропласте:
1,5-рибулозобисфосфат + O2 → 3-фосфоглицерат + 2-фосфогликолат
2-фосфогликолат → гликолат
Обменник: 2

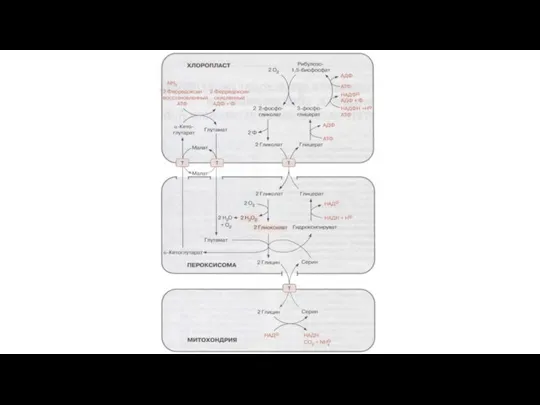
Слайд 80Фотодыхание
Реакции в пероксисоме:
2 гликолат + 2O2 → 2 глиоксилат + 2H2O2
2 глиоксилат
Фотодыхание
Реакции в пероксисоме:
2 гликолат + 2O2 → 2 глиоксилат + 2H2O2
2 глиоксилат

Слайд 82Фотодыхание
Энергетика реакции:
2х рибулозо-1,5-бисфосфат → 2х 3-фосфоглицерат + 2х 2-фосфогликолат (самопроизвольно)
2х 2-фосфогликолат →
Фотодыхание
Энергетика реакции:
2х рибулозо-1,5-бисфосфат → 2х 3-фосфоглицерат + 2х 2-фосфогликолат (самопроизвольно)
2х 2-фосфогликолат →

Слайд 83ВТОРИЧНЫЕ МЕТАБОЛИТЫ
ВТОРИЧНЫЕ МЕТАБОЛИТЫ

Слайд 84Различают первичные продукты обмена веществ клетки и вторичные, которые образуются из первичных
Различают первичные продукты обмена веществ клетки и вторичные, которые образуются из первичных



Слайд 87Существует несколько классификаций веществ вторичного происхождения.
1. ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ
3 Класса:
1. Фенольные соединения
2.
Существует несколько классификаций веществ вторичного происхождения.
1. ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ
3 Класса:
1. Фенольные соединения
2.

Слайд 882. Классификация по функциям:
2. Классификация по функциям:

Слайд 893. Физиологически активные вещества: фитогормоны и витамины.
4. Вещества с неизвестным физиологическим значением:
3. Физиологически активные вещества: фитогормоны и витамины.
4. Вещества с неизвестным физиологическим значением:

Слайд 901. ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ
ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ – вещества ароматической природы, содержащие одну (фенолы)
1. ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ
ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ – вещества ароматической природы, содержащие одну (фенолы)

Слайд 91Фенольные соединения накапливаются в разных частях растения в зависимости от вида:
цветках (медуница),
Фенольные соединения накапливаются в разных частях растения в зависимости от вида:
цветках (медуница),

Слайд 92ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ
ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ

Слайд 93Примеры окраски антоцианами
Примеры окраски антоцианами

Слайд 94Богаты катехинами: виноград, чай
Богаты катехинами: виноград, чай

Слайд 953. Полифенолы:
лигнины (обеспечивают одревеснение, т.е. опору растению),
таннины или
3. Полифенолы:
лигнины (обеспечивают одревеснение, т.е. опору растению),
таннины или

Слайд 96Кора и листья дуба богаты таннинами
Кора и листья дуба богаты таннинами

Слайд 972. АЛКАЛОИДЫ
2. АЛКАЛОИДЫ
Слайд 98Алкалоиды накапливаются в разных частях растения в зависимости от вида:
листьях (табака –
Алкалоиды накапливаются в разных частях растения в зависимости от вида:
листьях (табака –

Слайд 993. ИЗОПРЕНОИДЫ
3. ИЗОПРЕНОИДЫ

Слайд 100Представители изопреноидов:
Представители изопреноидов:


Слайд 102ИСПОЛЬЗОВАНИЕ ВТОРИЧНЫХ МЕТАБОЛИТОВ
ИСПОЛЬЗОВАНИЕ ВТОРИЧНЫХ МЕТАБОЛИТОВ



Слайд 105Эфирномасличные растения
Эфирномасличные растения

Слайд 106Обзор класса терпенов и некоторые типичные представители
Обзор класса терпенов и некоторые типичные представители

 Строение клетки
Строение клетки Теория катастроф Ж. Кювье
Теория катастроф Ж. Кювье Репликация ДНК
Репликация ДНК Жизнедеятельность бактерий. Питание
Жизнедеятельность бактерий. Питание Биология в естествознании
Биология в естествознании Ботаника. Введение. Корень
Ботаника. Введение. Корень 3. Урок - РНК и АТФ
3. Урок - РНК и АТФ Мутационная изменчивость
Мутационная изменчивость Урбопопуляции птиц
Урбопопуляции птиц Создание природного индикатора для определения pH-среды
Создание природного индикатора для определения pH-среды Изготовление кровяной муки
Изготовление кровяной муки Царство бактерии
Царство бактерии Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Презентация на тему Класс Двудольные, семейство Паслёновые
Презентация на тему Класс Двудольные, семейство Паслёновые  Ткани растений. Виды тканей. Обмен веществ
Ткани растений. Виды тканей. Обмен веществ Женская половая система - 1. Яичник. Жёлтое тело
Женская половая система - 1. Яичник. Жёлтое тело Презентация на тему Віруси. Пріони. Бактерії
Презентация на тему Віруси. Пріони. Бактерії  Отдел Покрытосеменные Класс Двудольные
Отдел Покрытосеменные Класс Двудольные Элемент металлического происхождения марганец
Элемент металлического происхождения марганец Fiziologia_1
Fiziologia_1 Биоэлектрогенез
Биоэлектрогенез Высшая нервная деятельность
Высшая нервная деятельность Животные и растения степей
Животные и растения степей Презентация на тему Зона степей (4 класс)
Презентация на тему Зона степей (4 класс)  Класс млекопитающие или звери
Класс млекопитающие или звери Репарация
Репарация Презентация на тему Главные части и органоиды клеток
Презентация на тему Главные части и органоиды клеток  Лишайники. Строение и питание лишайников
Лишайники. Строение и питание лишайников