- Практический курс морфологии архегониальных растений

Содержание
- 2. К.И. Мейер. Практический курс морфологии архегониальных растений. М, 1982
- 4. Takakia Род был описан в 1958 году. Его относили к печеночникам (а иногда даже сближали с
- 5. Wollemia nobilis – третий род семейства Araucariaceae Новый род, описан из Австралии в 1994 г.
- 6. Ticodendron incognitum http://www.plantsystematics.org
- 7. Число образцов, хранящихся в мировых ботанических коллекциях, для 317 б.м. случайно выбранных видов древесных тропических растений
- 8. Vaccinium (Ericaceae): около 450 видов, большое разнообразие в горных районах тропиков узкий ареал Vaccinium uliginosum -
- 9. Shortia rotata Gaddy & Nuraliev (Diapensiaceae). Вьетнам. Новые для науки виды, подвиды и разновидности, описанные сотрудниками
- 10. Aspidistra elatior
- 11. Aspidistra 1998 – 11 видов 2000 – 49 видов 2005 – 80 видов 2015 – 130
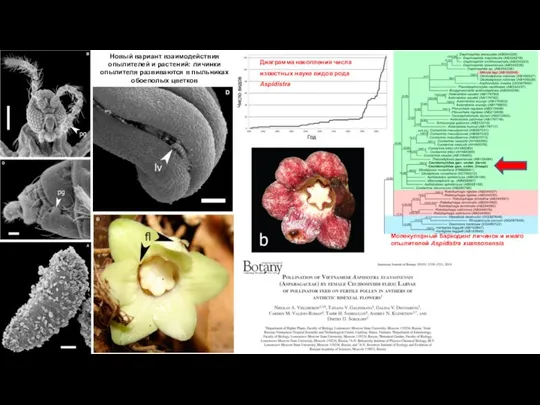
- 12. Молекулярный баркодинг личинок и имаго опылителей Aspidistra xuansonensis Диаграмма накопления числа известных науке видов рода Aspidistra
- 16. Ginkgo ♀
- 18. Типы систем живых организмов Искусственные – произвольный выбор небольшого числа признаков как основы системы Естественные –
- 19. Монофилия Полифилия Старое понимание терминов Монофилия Парафилия Полифилия «не-монофилия» Кладистическое понимание терминов
- 20. Soltis et al. 2011 17 генов Valerianaceae Dipsacaceae Morinaceae Caprifoliaceae s.l. Caprifoliaceae s.l. Adoxaceae Порядок Dipsacales
- 21. Soltis et al. 2011 17 генов Caprifoliaceae Adoxaceae Порядок Dipsacales Семейства по системе Stevens (2007, APWEB)
- 22. Кладизм, или «филогенетическая систематика» W. Hennig (1950, 1966) – основатель кладизма монофилия – парафилия – полифилия
- 23. Апоморфия – производное состояние признака Плезиоморфия – исходное состояние признака Родство определяют только по апоморфиям!!! Самое
- 24. Апоморфия – производное состояние признака Плезиоморфия – исходное состояние признака Родство определяют только по апоморфиям!!! Самое
- 25. Хара Сфагнум Плаун Дуб Ряска Мужские гаметы Сперматозоиды Спермии Ксилема Отсутствует Развита Конфликт признаков: в этом
- 34. Общий вид первичных данных, используемых в молекулярно-филогенетическом анализе Признак – позиция нуклеотида Состояние признака – A,
- 35. Однодольные Примитивные двудольные Высшие двудольные Филогения цветковых растений из работы Angiosperm Phylogeny Group (2003), данные в
- 36. Однодольные Примитивные двудольные Высшие двудольные Филогения покрытосеменных по Hans Hallier (1893-1912)
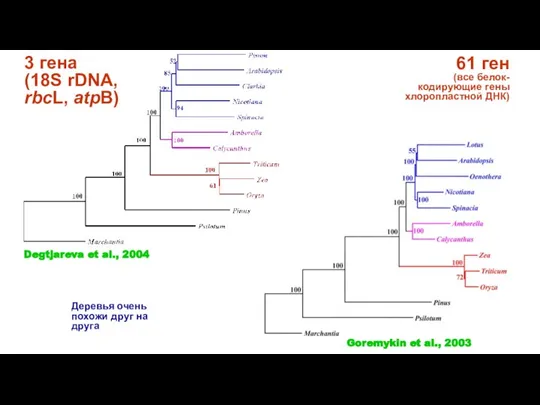
- 37. 61 ген (все белок-кодирующие гены хлоропластной ДНК) Примитивные двудольные Высшие двудольные Однодольные Goremykin et al., 2004
- 38. 3 гена (18S rDNA, rbcL, atpB) 61 ген (все белок-кодирующие гены хлоропластной ДНК) Деревья очень похожи
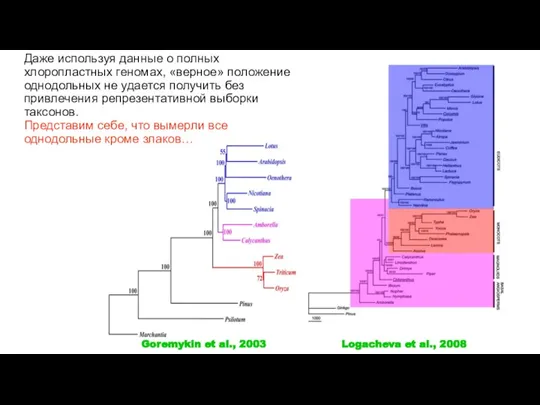
- 39. Logacheva et al., 2008 Goremykin et al., 2003 Даже используя данные о полных хлоропластных геномах, «верное»
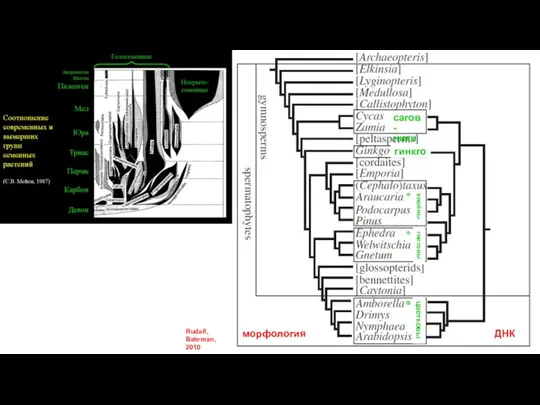
- 40. Rudall, Bateman, 2010 морфология ДНК сагов- ники гинкго хвойные гнетовые цветковые
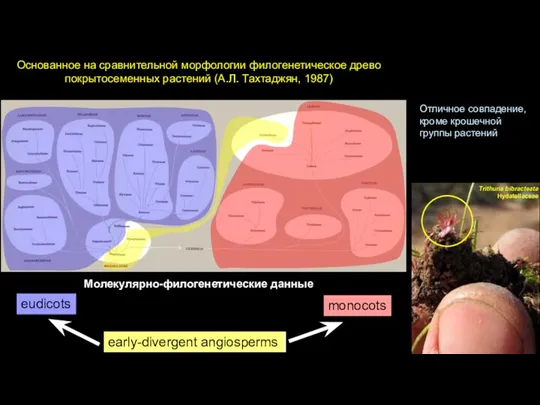
- 41. Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений (А.Л. Тахтаджян, 1987) Молекулярно-филогенетические данные Early-divergent angiosperms monocots
- 42. Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений (А.Л. Тахтаджян, 1987). Цветом показана выделяемая сейчас группа
- 43. subkingdom Embryophyta OK superdivision Tracheophyta OK division Spermatophyta OK subdivision Angiospermae ОК class Dicotyledones almost ОК
- 44. Platanus, Nelumbo, Proteaceae
- 45. А.Л.Тахтаджян (1966) Platanaceae
- 46. Все прочие высшие двудольные Все прочие высшие двудольные Soltis et al. 2011 17 генов Savolainen et
- 47. СПОРОФИТ СПОРАНГИИ СПОРЫ ГАМЕТОФИТ ГАМЕТАНГИИ (антеридии и архегонии) ГАМЕТЫ ЗИГОТА МЕЙОЗ ОПЛОДОТВОРЕНИЕ
- 48. Антеридии Архегонии яйцеклетка шейковые канальцевые клетки брюшная канальцевая клетка шейка шейка брюшко брюшко стенка сперматоцит сперматоцит
- 49. Уродливые стерильные гаметангии печеночника Corsinia marchantioides, сочетающие признаки антеридия и архегония
- 53. Скачать презентацию

Слайд 4Takakia
Род был описан в 1958 году. Его относили к печеночникам
(а иногда даже
Takakia
Род был описан в 1958 году. Его относили к печеночникам (а иногда даже

Слайд 5Wollemia nobilis – третий род семейства Araucariaceae
Новый род, описан из Австралии в
Wollemia nobilis – третий род семейства Araucariaceae
Новый род, описан из Австралии в

Слайд 6Ticodendron incognitum
http://www.plantsystematics.org
Ticodendron incognitum
http://www.plantsystematics.org

Слайд 7Число образцов, хранящихся в
мировых ботанических коллекциях,
для 317 б.м. случайно выбранных
видов
Число образцов, хранящихся в
мировых ботанических коллекциях,
для 317 б.м. случайно выбранных
видов

Слайд 8Vaccinium (Ericaceae): около 450 видов, большое разнообразие в горных районах тропиков
узкий ареал
Vaccinium
Vaccinium (Ericaceae): около 450 видов, большое разнообразие в горных районах тропиков
узкий ареал
Vaccinium

Слайд 9Shortia rotata Gaddy & Nuraliev (Diapensiaceae). Вьетнам.
Новые для науки виды, подвиды и
Shortia rotata Gaddy & Nuraliev (Diapensiaceae). Вьетнам.
Новые для науки виды, подвиды и

Слайд 10Aspidistra elatior
Aspidistra elatior

Слайд 11Aspidistra
1998 – 11 видов
2000 – 49 видов
2005 – 80 видов
2015 – 130
Aspidistra
1998 – 11 видов
2000 – 49 видов
2005 – 80 видов
2015 – 130

Слайд 12Молекулярный баркодинг личинок и имаго опылителей Aspidistra xuansonensis
Диаграмма накопления числа известных науке
Молекулярный баркодинг личинок и имаго опылителей Aspidistra xuansonensis
Диаграмма накопления числа известных науке


Слайд 16Ginkgo ♀
Ginkgo ♀
Слайд 18Типы систем живых организмов
Искусственные – произвольный выбор небольшого числа признаков как основы
Типы систем живых организмов
Искусственные – произвольный выбор небольшого числа признаков как основы

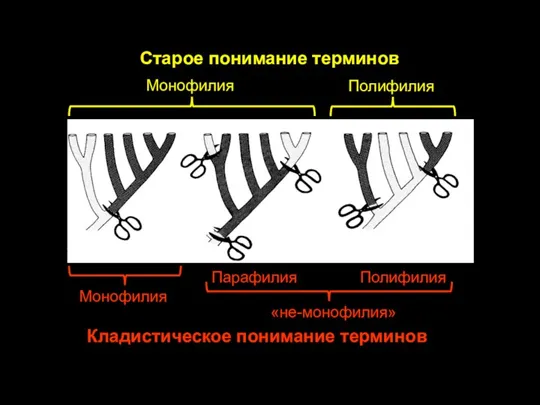
Слайд 19Монофилия
Полифилия
Старое понимание терминов
Монофилия
Парафилия
Полифилия
«не-монофилия»
Кладистическое понимание терминов
Монофилия
Полифилия
Старое понимание терминов
Монофилия
Парафилия
Полифилия
«не-монофилия»
Кладистическое понимание терминов
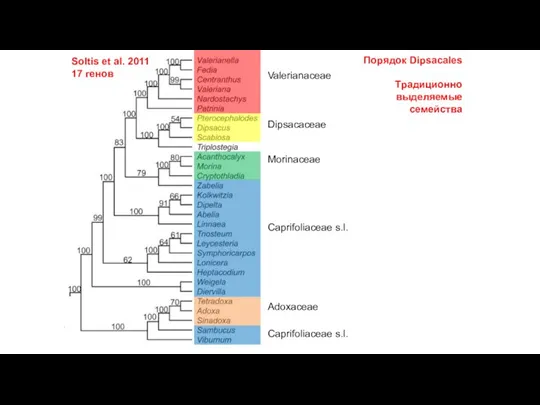
Слайд 20Soltis et al. 2011
17 генов
Valerianaceae
Dipsacaceae
Morinaceae
Caprifoliaceae s.l.
Caprifoliaceae s.l.
Adoxaceae
Порядок Dipsacales
Традиционно выделяемые семейства
Soltis et al. 2011
17 генов
Valerianaceae
Dipsacaceae
Morinaceae
Caprifoliaceae s.l.
Caprifoliaceae s.l.
Adoxaceae
Порядок Dipsacales
Традиционно выделяемые семейства
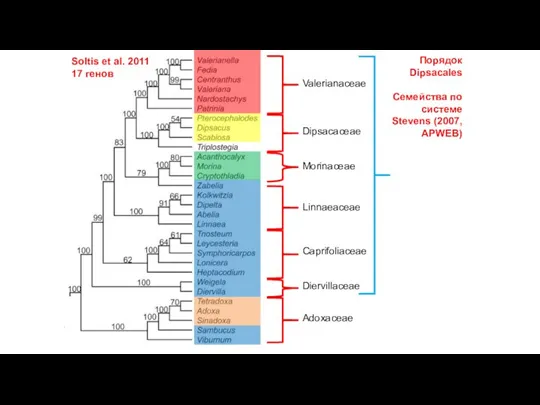
Слайд 21Soltis et al. 2011
17 генов
Caprifoliaceae
Adoxaceae
Порядок Dipsacales
Семейства по системе Stevens (2007, APWEB)
Diervillaceae
Linnaeaceae
Morinaceae
Dipsacaceae
Valerianaceae
Soltis et al. 2011
17 генов
Caprifoliaceae
Adoxaceae
Порядок Dipsacales
Семейства по системе Stevens (2007, APWEB)
Diervillaceae
Linnaeaceae
Morinaceae
Dipsacaceae
Valerianaceae
Слайд 22Кладизм,
или «филогенетическая систематика»
W. Hennig (1950, 1966) – основатель кладизма
монофилия – парафилия
Кладизм,
или «филогенетическая систематика»
W. Hennig (1950, 1966) – основатель кладизма
монофилия – парафилия

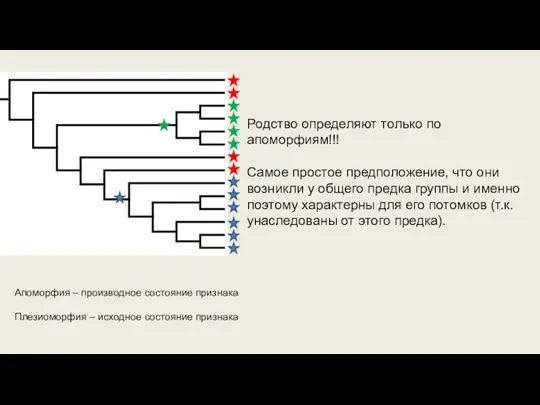
Слайд 23Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
Родство определяют только по
Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
Родство определяют только по
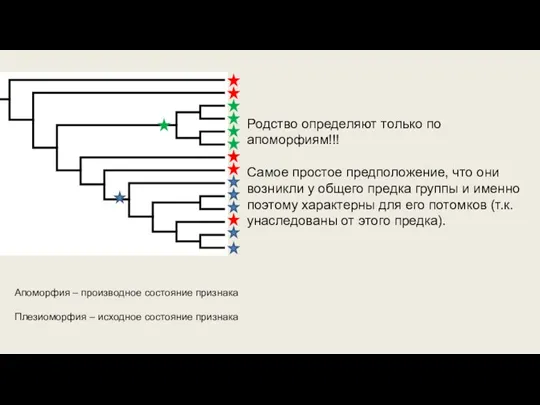
Слайд 24Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
Родство определяют только по
Апоморфия – производное состояние признака
Плезиоморфия – исходное состояние признака
Родство определяют только по
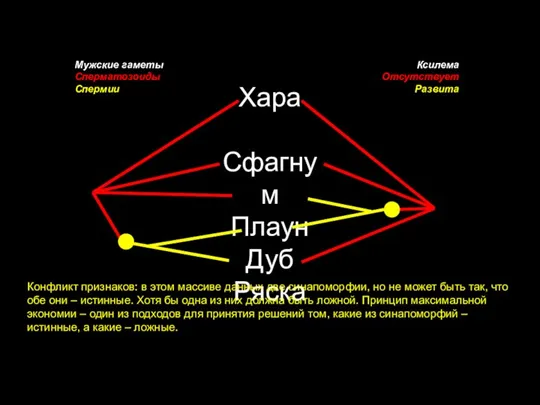
Слайд 25Хара
Сфагнум
Плаун
Дуб
Ряска
Мужские гаметы
Сперматозоиды
Спермии
Ксилема
Отсутствует
Развита
Конфликт признаков: в этом массиве данных две
Хара
Сфагнум
Плаун
Дуб
Ряска
Мужские гаметы
Сперматозоиды
Спермии
Ксилема
Отсутствует
Развита
Конфликт признаков: в этом массиве данных две
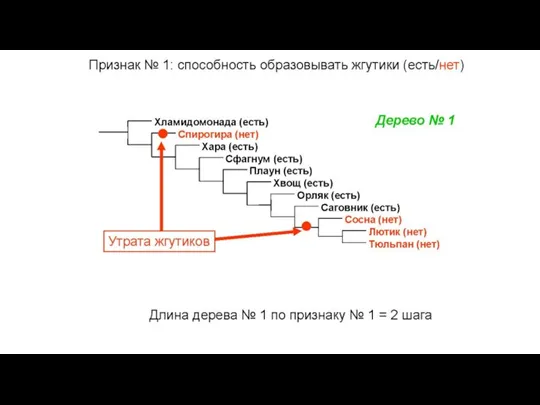
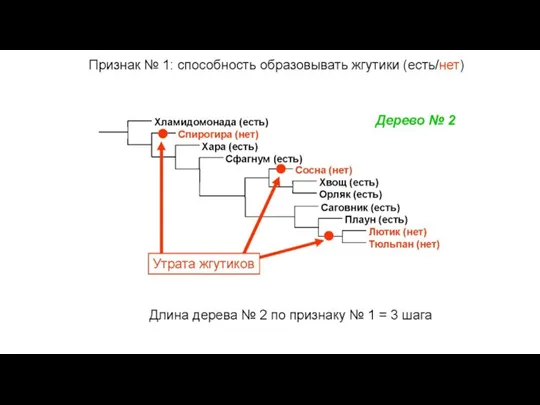

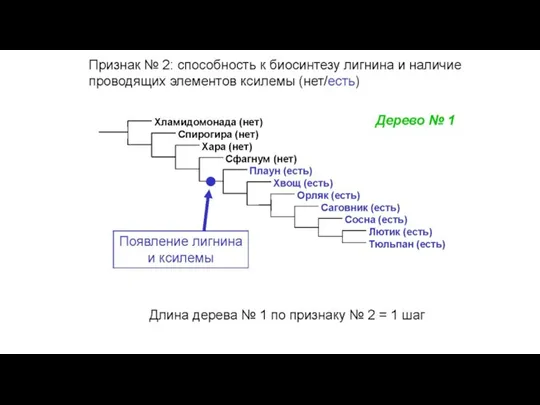
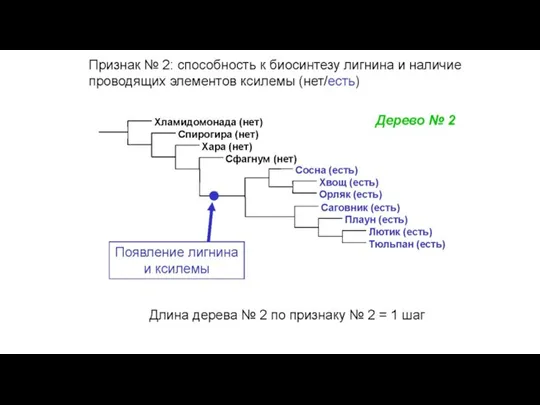
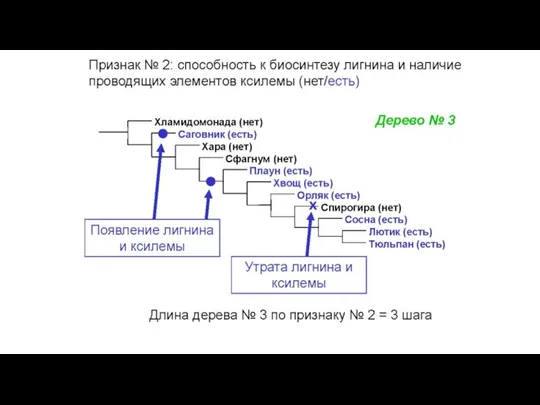

Слайд 34Общий вид первичных данных, используемых в молекулярно-филогенетическом анализе
Признак – позиция нуклеотида
Состояние признака
Общий вид первичных данных, используемых в молекулярно-филогенетическом анализе
Признак – позиция нуклеотида
Состояние признака

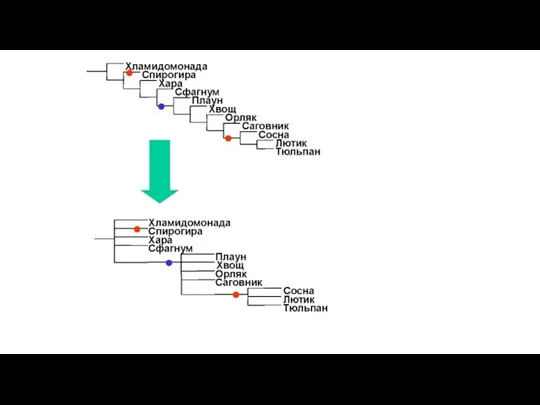
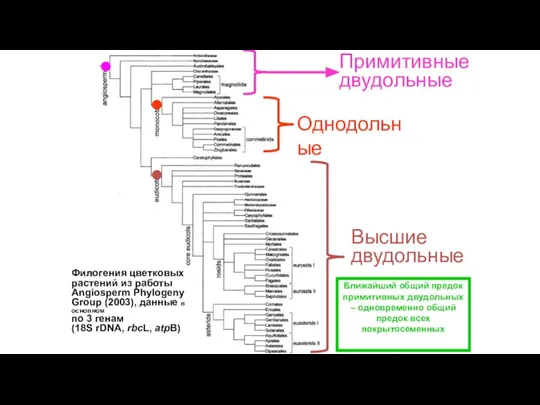
Слайд 35Однодольные
Примитивные двудольные
Высшие двудольные
Филогения цветковых растений из работы Angiosperm Phylogeny Group (2003), данные
Однодольные
Примитивные двудольные
Высшие двудольные
Филогения цветковых растений из работы Angiosperm Phylogeny Group (2003), данные
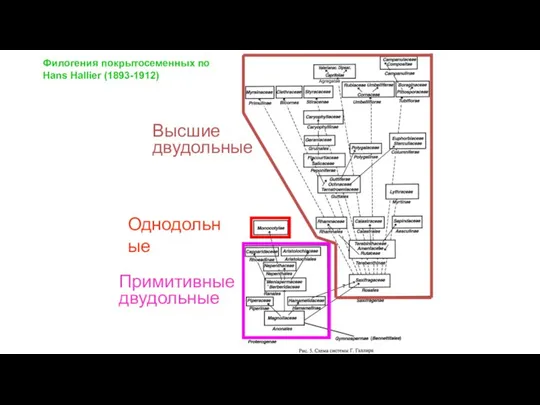
Слайд 36Однодольные
Примитивные двудольные
Высшие двудольные
Филогения покрытосеменных по
Hans Hallier (1893-1912)
Однодольные
Примитивные двудольные
Высшие двудольные
Филогения покрытосеменных по
Hans Hallier (1893-1912)
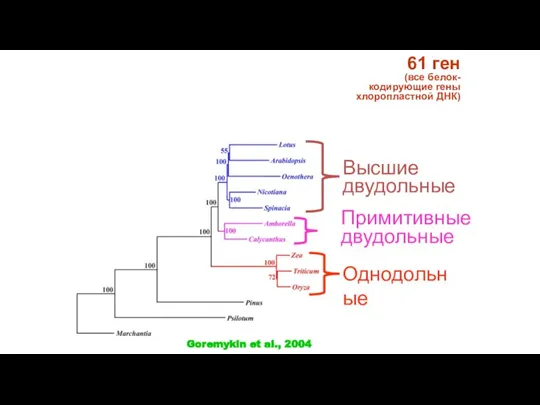
Слайд 3761 ген
(все белок-кодирующие гены хлоропластной ДНК)
Примитивные двудольные
Высшие двудольные
Однодольные
Goremykin et al., 2004
61 ген
(все белок-кодирующие гены хлоропластной ДНК)
Примитивные двудольные
Высшие двудольные
Однодольные
Goremykin et al., 2004
Слайд 383 гена
(18S rDNA, rbcL, atpB)
61 ген
(все белок-кодирующие гены хлоропластной ДНК)
Деревья
3 гена
(18S rDNA, rbcL, atpB)
61 ген
(все белок-кодирующие гены хлоропластной ДНК)
Деревья
Слайд 39Logacheva et al., 2008
Goremykin et al., 2003
Даже используя данные о полных хлоропластных
Logacheva et al., 2008
Goremykin et al., 2003
Даже используя данные о полных хлоропластных
Слайд 40Rudall, Bateman, 2010
морфология
ДНК
сагов-
ники
гинкго
хвойные
гнетовые
цветковые
Rudall, Bateman, 2010
морфология
ДНК
сагов-
ники
гинкго
хвойные
гнетовые
цветковые
Слайд 41Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений (А.Л. Тахтаджян, 1987)
Молекулярно-филогенетические данные
Early-divergent
Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений (А.Л. Тахтаджян, 1987)
Молекулярно-филогенетические данные
Early-divergent
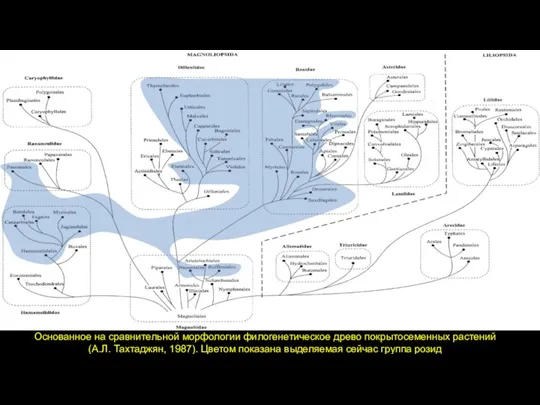
Слайд 42Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений
(А.Л. Тахтаджян, 1987). Цветом
Основанное на сравнительной морфологии филогенетическое древо покрытосеменных растений
(А.Л. Тахтаджян, 1987). Цветом
Слайд 43subkingdom Embryophyta OK
superdivision Tracheophyta OK
division Spermatophyta OK
subdivision Angiospermae ОК
class Dicotyledones almost ОК
subclass
subkingdom Embryophyta OK
superdivision Tracheophyta OK
division Spermatophyta OK
subdivision Angiospermae ОК
class Dicotyledones almost ОК
subclass

Слайд 44Platanus, Nelumbo, Proteaceae
Platanus, Nelumbo, Proteaceae

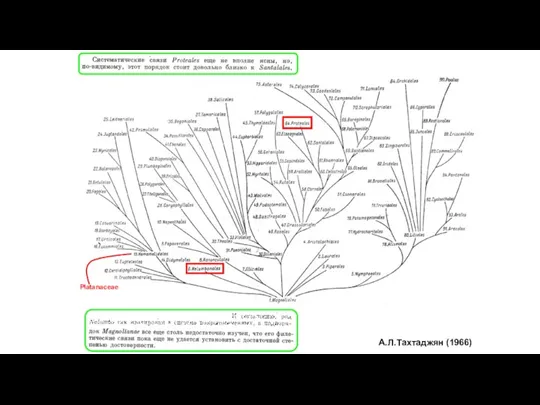
Слайд 45А.Л.Тахтаджян (1966)
Platanaceae
А.Л.Тахтаджян (1966)
Platanaceae
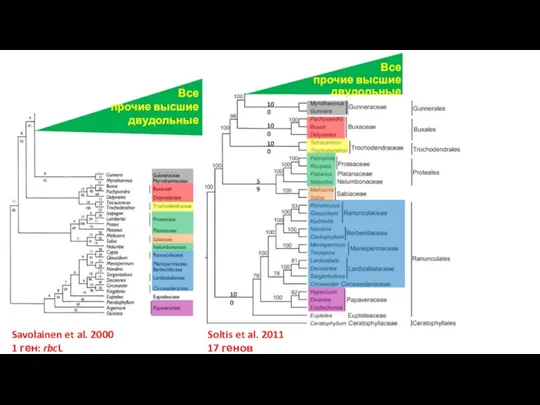
Слайд 46Все
прочие высшие двудольные
Все
прочие высшие двудольные
Soltis et al. 2011
17 генов
Savolainen et
Все
прочие высшие двудольные
Все
прочие высшие двудольные
Soltis et al. 2011
17 генов
Savolainen et
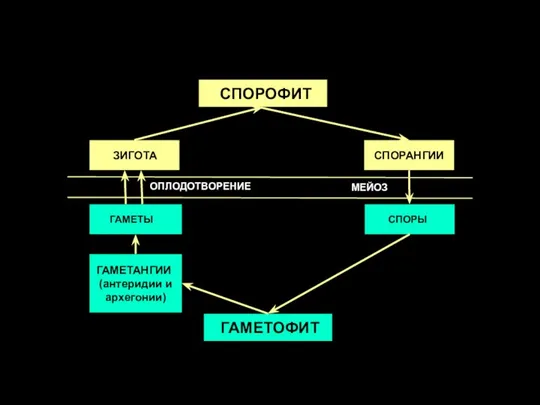
Слайд 47 СПОРОФИТ
СПОРАНГИИ
СПОРЫ
ГАМЕТОФИТ
ГАМЕТАНГИИ
(антеридии и архегонии)
ГАМЕТЫ
ЗИГОТА
МЕЙОЗ
ОПЛОДОТВОРЕНИЕ
СПОРОФИТ
СПОРАНГИИ
СПОРЫ
ГАМЕТОФИТ
ГАМЕТАНГИИ
(антеридии и архегонии)
ГАМЕТЫ
ЗИГОТА
МЕЙОЗ
ОПЛОДОТВОРЕНИЕ
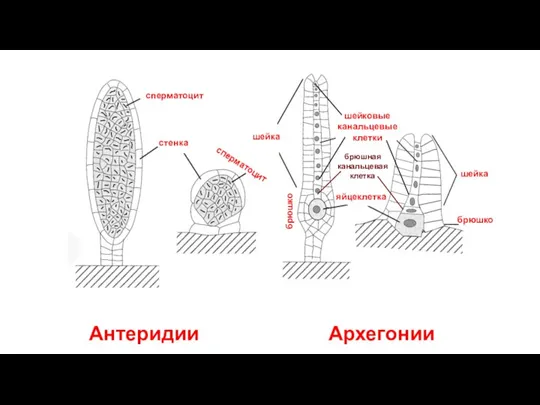
Слайд 48Антеридии Архегонии
яйцеклетка
шейковые
канальцевые
клетки
брюшная канальцевая клетка
шейка
шейка
брюшко
брюшко
стенка
сперматоцит
сперматоцит
Антеридии Архегонии
яйцеклетка
шейковые
канальцевые
клетки
брюшная канальцевая клетка
шейка
шейка
брюшко
брюшко
стенка
сперматоцит
сперматоцит
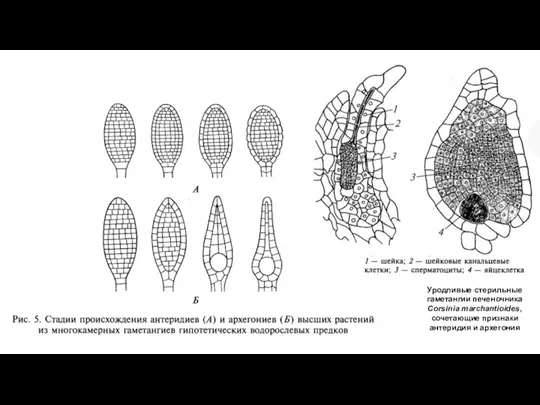
Слайд 49Уродливые стерильные гаметангии печеночника Corsinia marchantioides, сочетающие признаки антеридия и архегония
Уродливые стерильные гаметангии печеночника Corsinia marchantioides, сочетающие признаки антеридия и архегония
 Безусловные рефлексы
Безусловные рефлексы Презентация на тему Отряды млекопитающих. Грызуны. Зайцеобразные.
Презентация на тему Отряды млекопитающих. Грызуны. Зайцеобразные.  Презентация на тему Охрана растений
Презентация на тему Охрана растений  Презентация на тему Миграции животных
Презентация на тему Миграции животных  Жизнь растений осенью
Жизнь растений осенью Тип Кольчатые черви
Тип Кольчатые черви Презентация на тему АМИНОКИСЛОТЫ И БЕЛКИ. СТРОЕНИЕ И СВОЙСТВА
Презентация на тему АМИНОКИСЛОТЫ И БЕЛКИ. СТРОЕНИЕ И СВОЙСТВА  Губа. Нитевидные сосочки языка
Губа. Нитевидные сосочки языка Презентация на тему Растительный и животный мир Арктики
Презентация на тему Растительный и животный мир Арктики  Manchots royaux
Manchots royaux Атырау ОФ бойынша 2018 жылдың 9 айлық қызметінің қорытындысы
Атырау ОФ бойынша 2018 жылдың 9 айлық қызметінің қорытындысы Семязачаток и семя
Семязачаток и семя Презентация на тему Животные, истребленные человеком
Презентация на тему Животные, истребленные человеком  Клетки иммунной системы
Клетки иммунной системы Презентация на тему Палеозой. Ордовик
Презентация на тему Палеозой. Ордовик  Дыхательная система, кожа
Дыхательная система, кожа Животные Красной книги. Выхухоль
Животные Красной книги. Выхухоль Кожа человека
Кожа человека Лук репчатый. Выращивание лука в почве с использованием удобрений
Лук репчатый. Выращивание лука в почве с использованием удобрений Структура и роль кожи в процессе выделения. 8 класс
Структура и роль кожи в процессе выделения. 8 класс odnokletochnye-1
odnokletochnye-1 Клетки универсальны
Клетки универсальны Органы чувств. Анализаторы
Органы чувств. Анализаторы Понятие видообразование. Формы и механизмы видообразования
Понятие видообразование. Формы и механизмы видообразования Презентация на тему ОСОБЕННОСТИ РАЗМНОЖЕНИЯ РЫБ
Презентация на тему ОСОБЕННОСТИ РАЗМНОЖЕНИЯ РЫБ  Царство Растений
Царство Растений Цитология. Клетка - универсальная единица живой материи
Цитология. Клетка - универсальная единица живой материи Презентация12. Дыхательная система
Презентация12. Дыхательная система