- Происхождение трансляционных процессов и генетического кода

Содержание
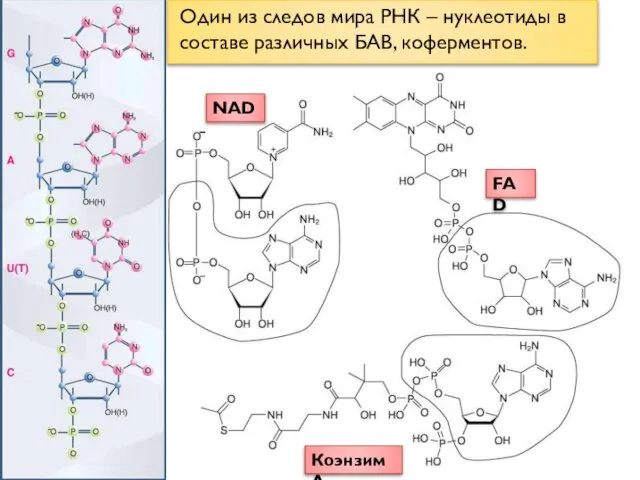
- 2. Один из следов мира РНК – нуклеотиды в составе различных БАВ, коферментов. NAD Коэнзим А FAD
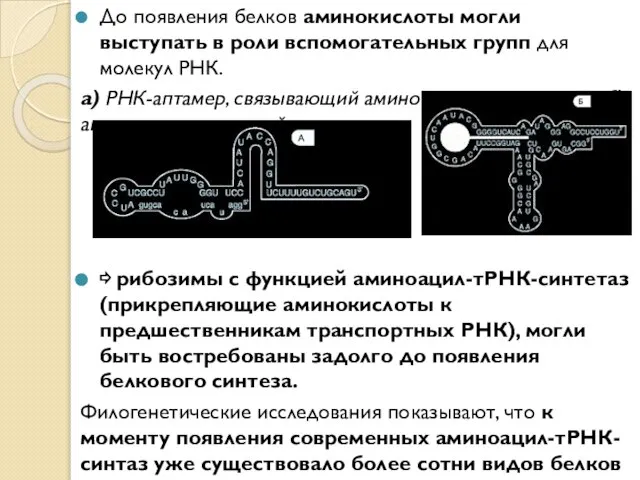
- 3. До появления белков аминокислоты могли выступать в роли вспомогательных групп для молекул РНК. а) РНК-аптамер, связывающий
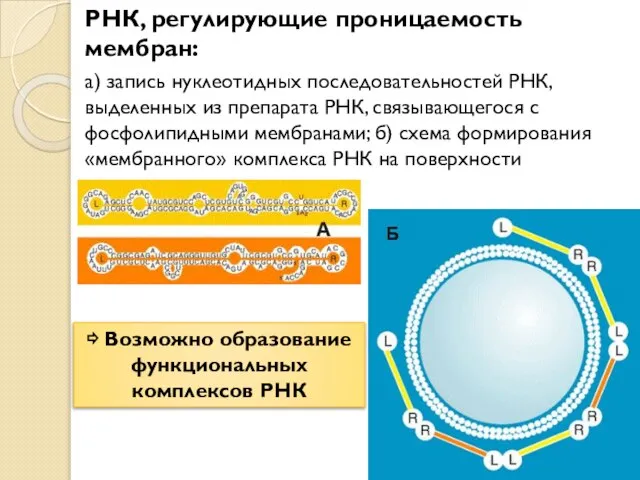
- 4. РНК, регулирующие проницаемость мембран: а) запись нуклеотидных последовательностей РНК, выделенных из препарата РНК, связывающегося с фосфолипидными
- 5. Транспортные РНК – важнейшая деталь системы синтеза белка Посредник между аминокислотами и нуклеотидами; Короче других типов
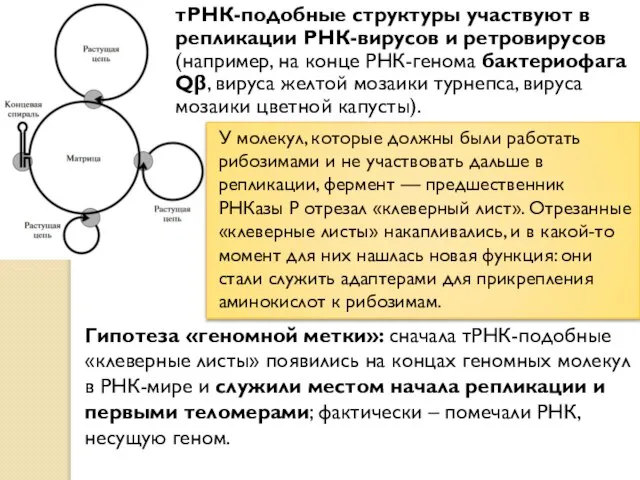
- 6. тРНК-подобные структуры участвуют в репликации РНК-вирусов и ретровирусов (например, на конце РНК-генома бактериофага Qβ, вируса желтой
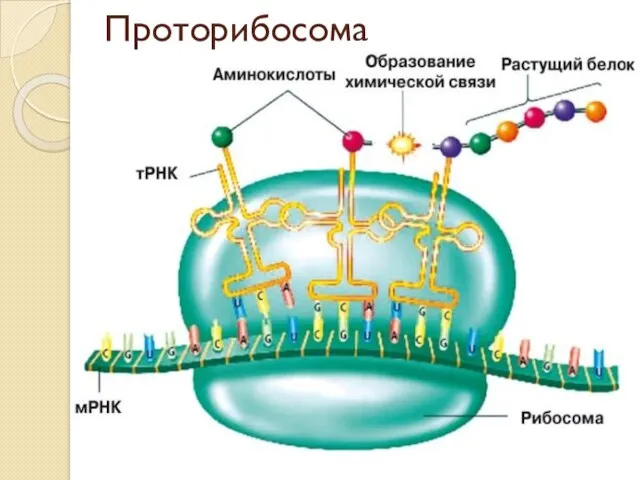
- 7. Проторибосома Рибосомы могли сформироваться в результате постепенной эволюции из очень простой маленькой молекулы РНК — «проторибосомы»,
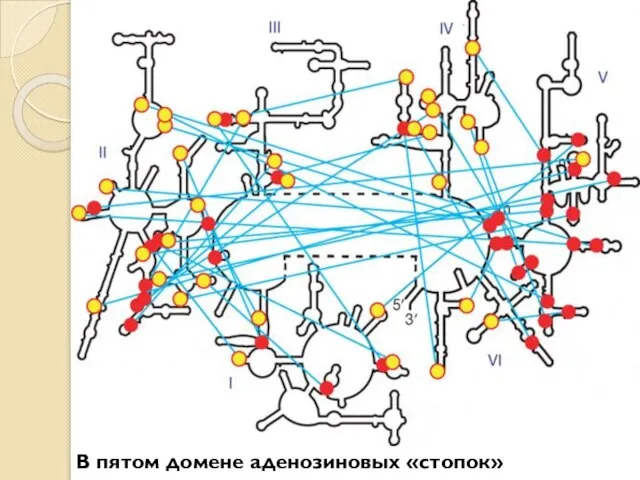
- 8. А-минорные взаимодействия необходимы для поддержания стабильной трехмерной структуры той части молекулы, к которой принадлежит аденозиновая «стопка»,
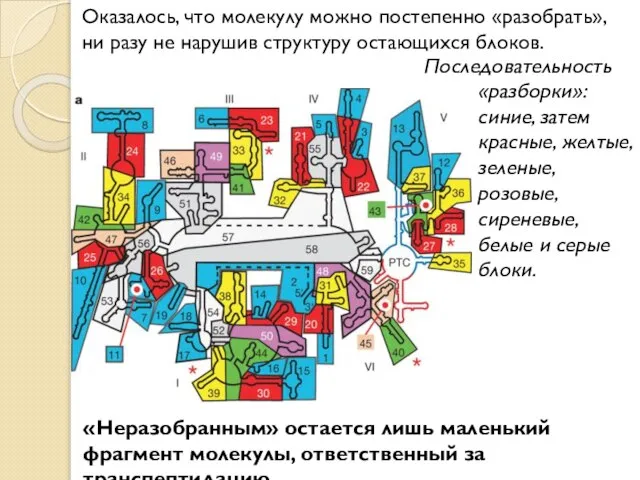
- 9. Оказалось, что молекулу можно постепенно «разобрать», ни разу не нарушив структуру остающихся блоков. «Неразобранным» остается лишь
- 10. Красным цветом выделена проторибосома, желтым — «выросты» (1–3; помогают рибосоме выбирать «правильную» тРНК), сиреневым — малая
- 11. Многие белки, образующие альфа-спираль, могут неспецифически связываться с двуспиральной РНК. При этом карбонильные группы пептидной цепи
- 12. Теории возникновения связи между аминокислотами и нуклеотидными триплетами: «Застывшая случайность»; Оптимизация на минимум ошибок белкового синтеза:
- 13. Теория оптимизации: Цель – чтобы при ошибках трансляции аминокислоты заменялись на химически похожие. При этом вероятность
- 14. Теория стереохимического соответствия Для восьми аминокислот с большими боковыми цепями были выделены аптамеры, значительно обогащенные триплетами
- 15. Три аминокислоты, синтезируемые в одну стадию из альфа-кетокислот — аланин, аспартат и глутамат, — имеют кодоны,
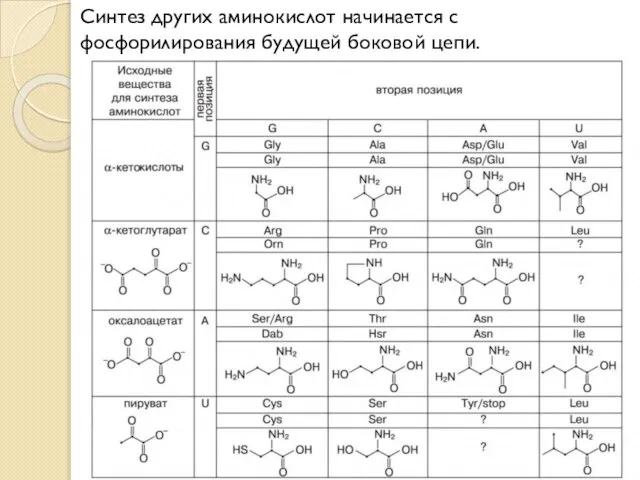
- 16. Синтез других аминокислот начинается с фосфорилирования будущей боковой цепи.
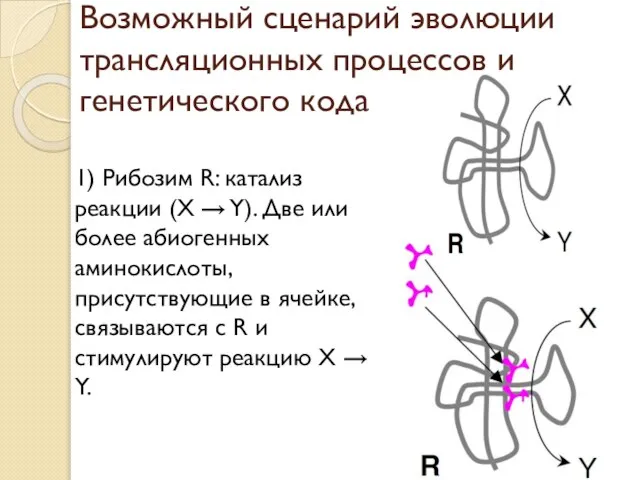
- 17. Возможный сценарий эволюции трансляционных процессов и генетического кода 1) Рибозим R: катализ реакции (X → Y).
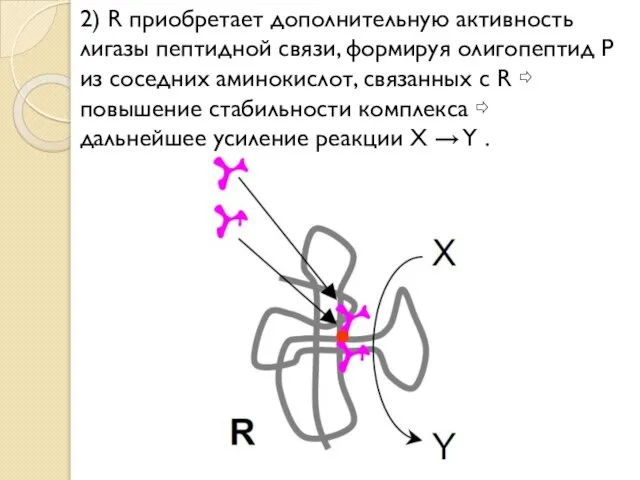
- 18. 2) R приобретает дополнительную активность лигазы пептидной связи, формируя олигопептид P из соседних аминокислот, связанных с
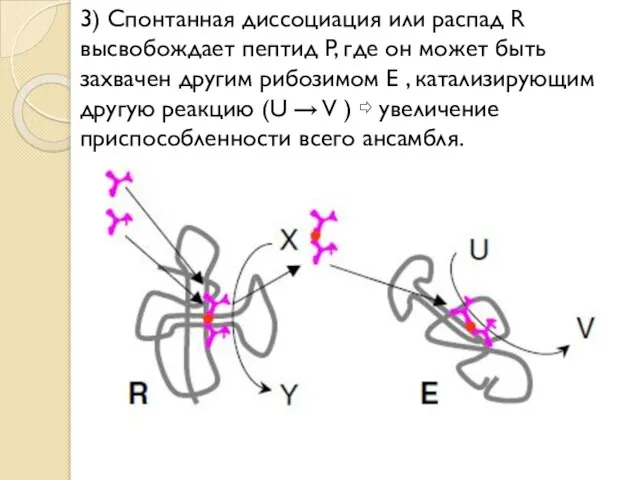
- 19. 3) Спонтанная диссоциация или распад R высвобождает пептид P, где он может быть захвачен другим рибозимом
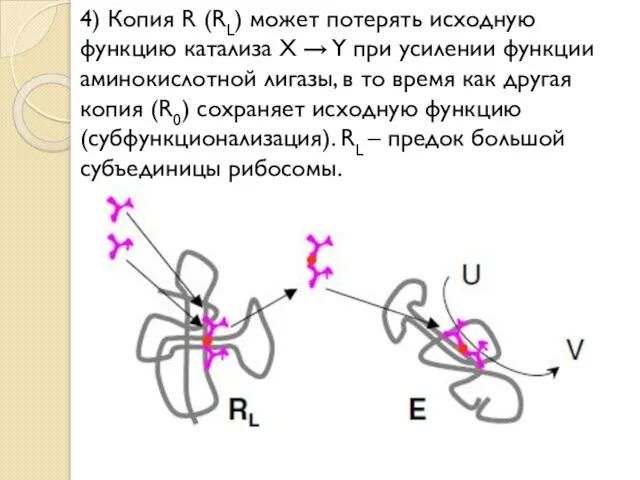
- 20. 4) Копия R (RL) может потерять исходную функцию катализа X → Y при усилении функции аминокислотной
- 21. 5) Катализ при помощи пептидов ⇨ аминокислоты становятся ценным ресурсом ⇨ развиваются малые РНК (T), связывающие
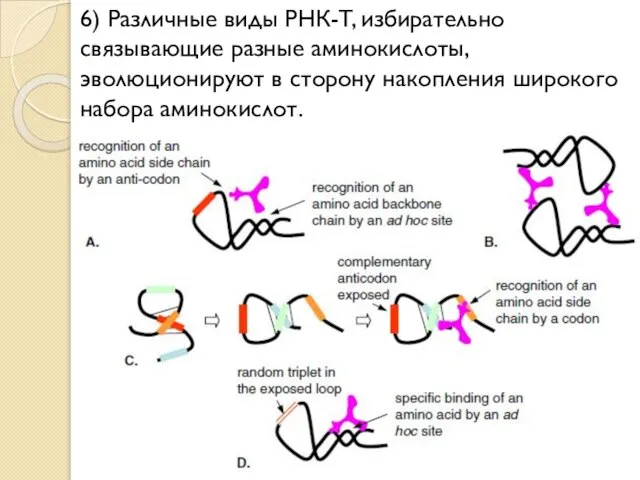
- 22. 6) Различные виды РНК-T, избирательно связывающие разные аминокислоты, эволюционируют в сторону накопления широкого набора аминокислот.
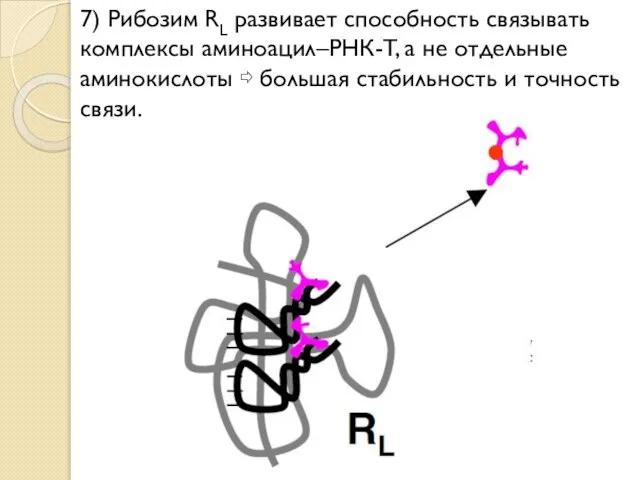
- 23. 7) Рибозим RL развивает способность связывать комплексы аминоацил–РНК-T, а не отдельные аминокислоты ⇨ большая стабильность и
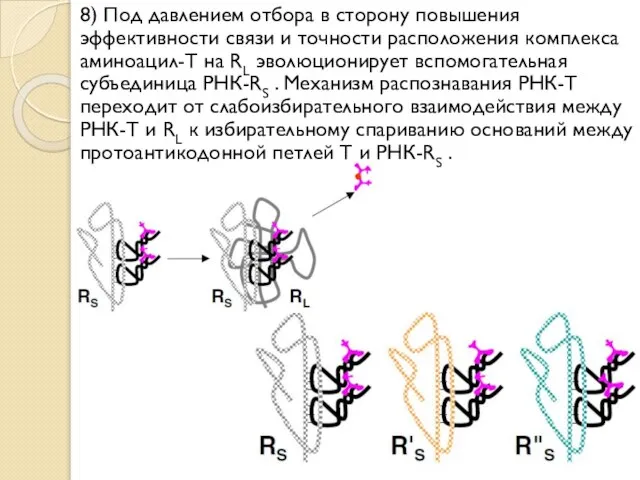
- 24. 8) Под давлением отбора в сторону повышения эффективности связи и точности расположения комплекса аминоацил-T на RL
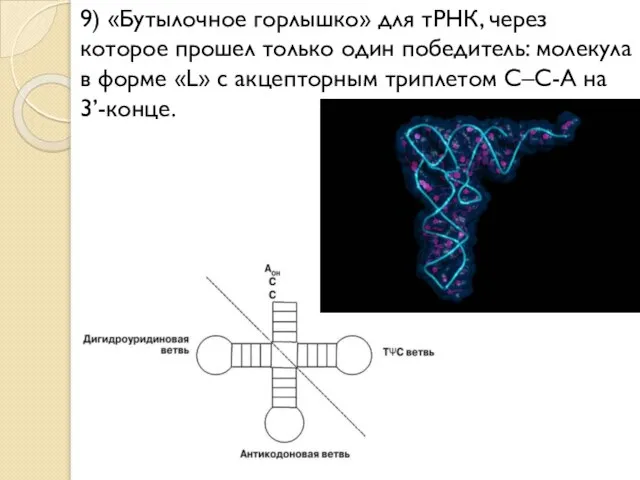
- 25. 9) «Бутылочное горлышко» для тРНК, через которое прошел только один победитель: молекула в форме «L» с
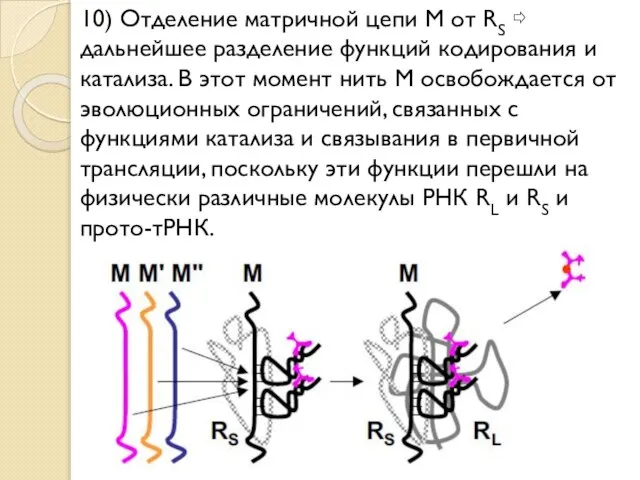
- 26. 10) Отделение матричной цепи M от RS ⇨ дальнейшее разделение функций кодирования и катализа. В этот
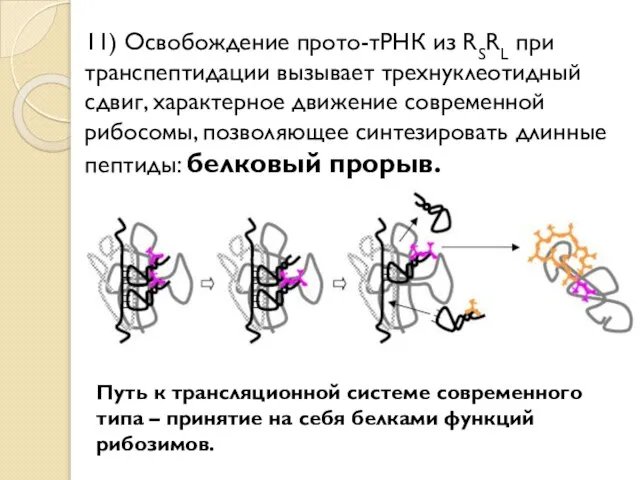
- 27. 11) Освобождение прото-тРНК из RSRL при транспептидации вызывает трехнуклеотидный сдвиг, характерное движение современной рибосомы, позволяющее синтезировать
- 29. Скачать презентацию
Слайд 3До появления белков аминокислоты могли выступать в роли вспомогательных групп для молекул
До появления белков аминокислоты могли выступать в роли вспомогательных групп для молекул
Слайд 4РНК, регулирующие проницаемость мембран:
а) запись нуклеотидных последовательностей РНК, выделенных из препарата РНК,
РНК, регулирующие проницаемость мембран:
а) запись нуклеотидных последовательностей РНК, выделенных из препарата РНК,
Слайд 5Транспортные РНК – важнейшая деталь системы синтеза белка
Посредник между аминокислотами и
Транспортные РНК – важнейшая деталь системы синтеза белка
Посредник между аминокислотами и
Слайд 6тРНК-подобные структуры участвуют в репликации РНК-вирусов и ретровирусов (например, на конце РНК-генома
тРНК-подобные структуры участвуют в репликации РНК-вирусов и ретровирусов (например, на конце РНК-генома
Слайд 7Проторибосома
Рибосомы могли сформироваться в результате постепенной эволюции из очень простой маленькой молекулы РНК —
Проторибосома
Рибосомы могли сформироваться в результате постепенной эволюции из очень простой маленькой молекулы РНК —
Слайд 8А-минорные взаимодействия необходимы для поддержания стабильной трехмерной структуры той части молекулы, к
А-минорные взаимодействия необходимы для поддержания стабильной трехмерной структуры той части молекулы, к
Слайд 9Оказалось, что молекулу можно постепенно «разобрать», ни разу не нарушив структуру остающихся
Оказалось, что молекулу можно постепенно «разобрать», ни разу не нарушив структуру остающихся
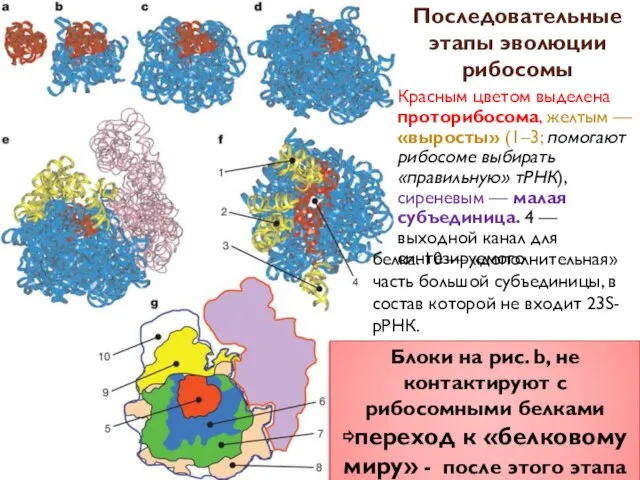
Слайд 10Красным цветом выделена проторибосома, желтым — «выросты» (1–3; помогают рибосоме выбирать «правильную»
Красным цветом выделена проторибосома, желтым — «выросты» (1–3; помогают рибосоме выбирать «правильную»
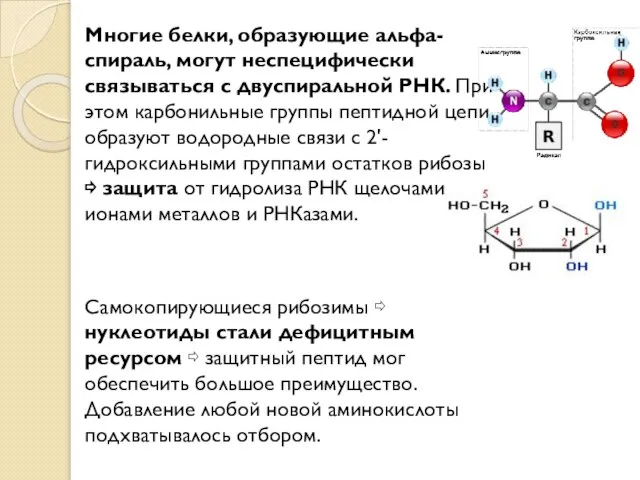
Слайд 11Многие белки, образующие альфа-спираль, могут неспецифически связываться с двуспиральной РНК. При этом
Многие белки, образующие альфа-спираль, могут неспецифически связываться с двуспиральной РНК. При этом
Слайд 12Теории возникновения связи между аминокислотами и нуклеотидными триплетами:
«Застывшая случайность»;
Оптимизация на минимум ошибок
Теории возникновения связи между аминокислотами и нуклеотидными триплетами:
«Застывшая случайность»;
Оптимизация на минимум ошибок

Слайд 13Теория оптимизации:
Цель – чтобы при ошибках трансляции аминокислоты заменялись на химически похожие.
Теория оптимизации:
Цель – чтобы при ошибках трансляции аминокислоты заменялись на химически похожие.

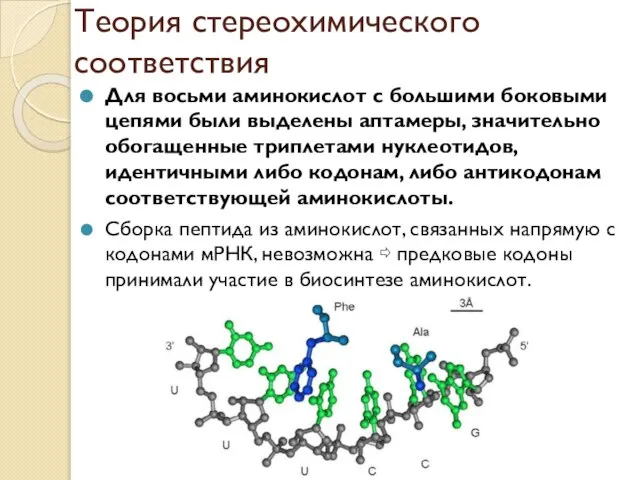
Слайд 14Теория стереохимического соответствия
Для восьми аминокислот с большими боковыми цепями были выделены
Теория стереохимического соответствия
Для восьми аминокислот с большими боковыми цепями были выделены
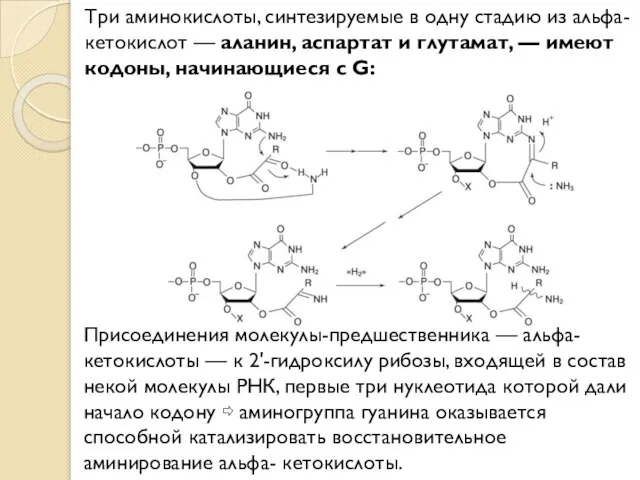
Слайд 15Три аминокислоты, синтезируемые в одну стадию из альфа-кетокислот — аланин, аспартат и
Три аминокислоты, синтезируемые в одну стадию из альфа-кетокислот — аланин, аспартат и
Слайд 16Синтез других аминокислот начинается с фосфорилирования будущей боковой цепи.
Синтез других аминокислот начинается с фосфорилирования будущей боковой цепи.
Слайд 17Возможный сценарий эволюции трансляционных процессов и генетического кода
1) Рибозим R: катализ реакции
Возможный сценарий эволюции трансляционных процессов и генетического кода
1) Рибозим R: катализ реакции
Слайд 182) R приобретает дополнительную активность лигазы пептидной связи, формируя олигопептид P из
2) R приобретает дополнительную активность лигазы пептидной связи, формируя олигопептид P из
Слайд 193) Спонтанная диссоциация или распад R высвобождает пептид P, где он может
3) Спонтанная диссоциация или распад R высвобождает пептид P, где он может
Слайд 204) Копия R (RL) может потерять исходную функцию катализа X → Y
4) Копия R (RL) может потерять исходную функцию катализа X → Y
Слайд 215) Катализ при помощи пептидов ⇨ аминокислоты становятся ценным ресурсом ⇨ развиваются
5) Катализ при помощи пептидов ⇨ аминокислоты становятся ценным ресурсом ⇨ развиваются

Слайд 226) Различные виды РНК-T, избирательно связывающие разные аминокислоты, эволюционируют в сторону накопления
6) Различные виды РНК-T, избирательно связывающие разные аминокислоты, эволюционируют в сторону накопления
Слайд 237) Рибозим RL развивает способность связывать комплексы аминоацил–РНК-T, а не отдельные аминокислоты
7) Рибозим RL развивает способность связывать комплексы аминоацил–РНК-T, а не отдельные аминокислоты
Слайд 248) Под давлением отбора в сторону повышения эффективности связи и точности расположения
8) Под давлением отбора в сторону повышения эффективности связи и точности расположения
Слайд 259) «Бутылочное горлышко» для тРНК, через которое прошел только один победитель: молекула
9) «Бутылочное горлышко» для тРНК, через которое прошел только один победитель: молекула
Слайд 2610) Отделение матричной цепи M от RS ⇨ дальнейшее разделение функций кодирования
10) Отделение матричной цепи M от RS ⇨ дальнейшее разделение функций кодирования
Слайд 2711) Освобождение прото-тРНК из RSRL при транспептидации вызывает трехнуклеотидный сдвиг, характерное движение
11) Освобождение прото-тРНК из RSRL при транспептидации вызывает трехнуклеотидный сдвиг, характерное движение
 Белки. Функции белков
Белки. Функции белков Деление клеток
Деление клеток Контрольная работа. Генетика
Контрольная работа. Генетика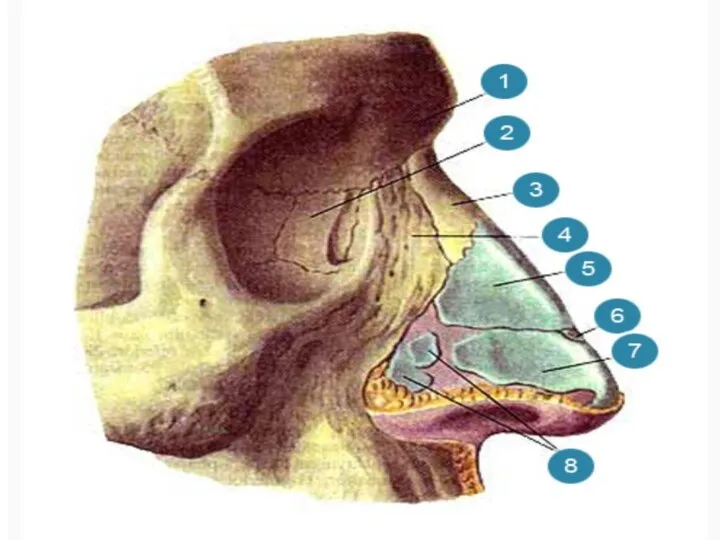 Органы дыхания
Органы дыхания Зрительный анализатор. Тест
Зрительный анализатор. Тест Австралопитек. Антропогенез. Эволюция человека. Часть 3
Австралопитек. Антропогенез. Эволюция человека. Часть 3 Растения луга
Растения луга Мхи. Папоротники. Голосеменные
Мхи. Папоротники. Голосеменные Презентация на тему Класс Двудольные, семейство Паслёновые
Презентация на тему Класс Двудольные, семейство Паслёновые  Триасово-Юрское вымирание динозавров
Триасово-Юрское вымирание динозавров Психофизиология. Память
Психофизиология. Память Строение органов человека
Строение органов человека Дыхание растений
Дыхание растений Презентация на тему "Лимфатическая система" - презентации по Биологии
Презентация на тему "Лимфатическая система" - презентации по Биологии Всё о чае
Всё о чае Презентация на тему Критерии живых систем
Презентация на тему Критерии живых систем  Сцепленное наследование. Закон Томаса Ханта Моргана
Сцепленное наследование. Закон Томаса Ханта Моргана Змеи. Систематика
Змеи. Систематика Митоз – способ деления эукариотических клеток
Митоз – способ деления эукариотических клеток Рефрактогенез
Рефрактогенез Первоцеты в природе
Первоцеты в природе Обитатели тайги
Обитатели тайги Физиология сосудов
Физиология сосудов Chromalveolata. (Часть 2)
Chromalveolata. (Часть 2) Аквариумные рыбки
Аквариумные рыбки Тест. Рыбы. 7 класс
Тест. Рыбы. 7 класс Презентация на тему Кровь и кровообращение
Презентация на тему Кровь и кровообращение  Окружающий мир. Тест
Окружающий мир. Тест