- ФИЗИОЛОГИЯ МЫШЦ

Содержание
- 2. ФИЗИОЛОГИЯ СКЕЛЕТНЫХ МЫШЦ
- 3. ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ Двигательная. Обменная – в мышцах происходят биохимические процессы, приводящие к изменению уровня мономеров
- 4. ТИПЫ СКЕЛЕТНЫХ МЫШЦ Параллельно- волокнистый Перистые Веретено-образный
- 5. СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ Физические: Эластичность – способность изменять длину под действием деформирующей силы Упругость – способность
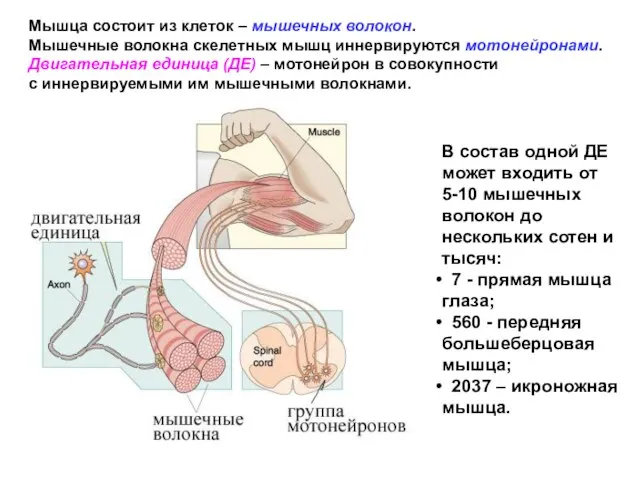
- 6. Мышца состоит из клеток – мышечных волокон. Мышечные волокна скелетных мышц иннервируются мотонейронами. Двигательная единица (ДЕ)
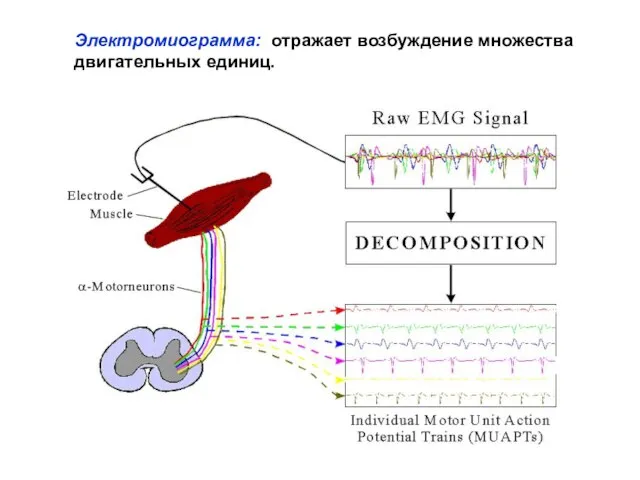
- 7. Электромиограмма: отражает возбуждение множества двигательных единиц.
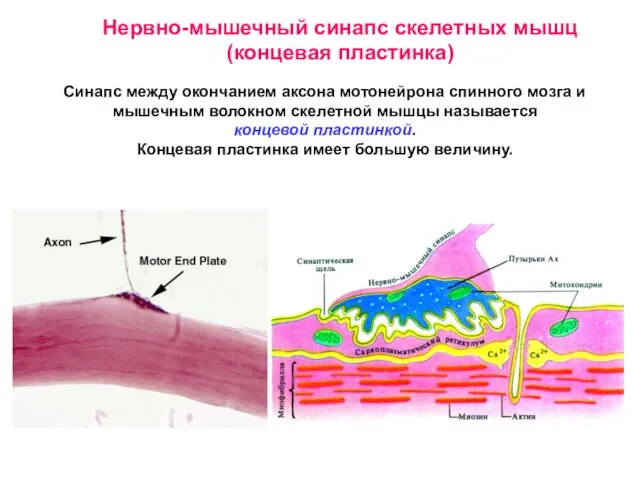
- 8. Нервно-мышечный синапс скелетных мышц (концевая пластинка) Синапс между окончанием аксона мотонейрона спинного мозга и мышечным волокном
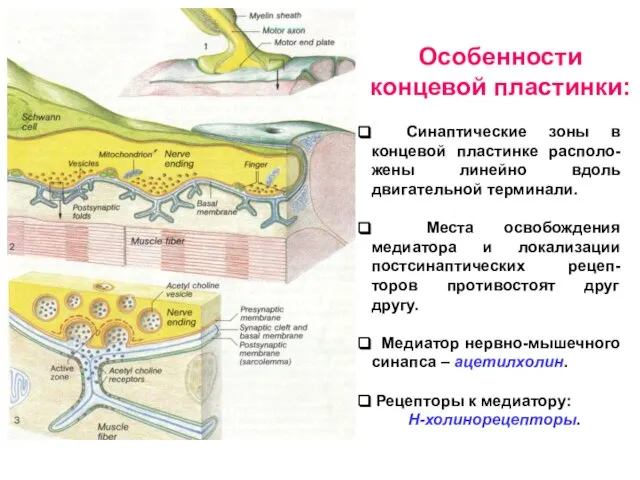
- 9. Синаптические зоны в концевой пластинке располо-жены линейно вдоль двигательной терминали. Места освобождения медиатора и локализации постсинаптических
- 10. ПОТЕНЦИАЛ КОНЦЕВОЙ ПЛАСТИНКИ В нервно-мышечном синапсе на постсинаптической мембране возникает потенциал концевой пластинки (ПКП): ПКП имеет
- 11. Квант – содержимое 1 везикулы. Выделение отдельных квантов в покое: на постсинаптической мембране возникают слабые потенциалы
- 12. ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ Одиночное сокращение Одиночное сокращение мышцы возникает при ее однократном раздражении. Фазы: А. Латентный
- 13. Тетаническое сокращение (тетанус) – суммация сокращений мышцы при действии ритмических раздражителей. Виды тетануса: Зубчатый тетанус: наступает,
- 14. Следовая деполяризация Вторичная экзальтация Реполяризация Рефрактерность
- 15. Режимы мышечного сокращения Изометрический – длина мышцы постоянна, но развивается внутреннее напряжение за счет силы растяжения
- 16. Типы мышечных волокон
- 17. Мышца состоит из мышечных волокон. СТРОЕНИЕ СКЕЛЕТНЫХ МЫШЦ
- 18. СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА миофибриллы, состоящие из саркомеров; саркоплазматический ретикулум - депо Са++; митохондрии; плазматическая мембрана мышечного
- 19. Миофибриллы содержат саркомеры – структурно-функциональные единицы мышцы.
- 20. СТРОЕНИЕ САРКОМЕРА МЫШЕЧНОГО ВОЛОКНА
- 21. Саркомер состоит из сократительных белков и Z-мембран. Z-мембраны располагаются в области Т-трубочек. К Z-мембранам крепятся тонкие
- 22. Строение актина и миозина Миозиновые нити имеют выступы – поперечные мостики. На их концах расположены утолщения
- 23. Механизм скольжения нитей актина и миозина. Электромеханическое сопряжение. Ион Са++ – посредник между возбуждением и сокращением.
- 24. Механизм скольжения нитей: укорочение саркомера 6. Гидролиз АТФ головки миозина, отсоединение АДФ и Ф – высвобождение
- 25. Электромеханическое сопряжение
- 26. Поперечный мостик распрямляется, головка миозина присоединяется к следующему активному центру актина. Механизм скольжения: разобщение нитей Отсоединение
- 28. Расслабление мышцы происходит в результате снижения уровня Са++ в цитоплазме. Са++ закачивается обратно в саркоплазматический ретикулум
- 29. ЗАВИСИМОСТЬ СИЛЫ СОКРАЩЕНИЯ ОТ ИСХОДНОЙ ДЛИНЫ МЫШЦЫ
- 30. ФИЗИОЛОГИЯ ГЛАДКИХ МЫШЦ
- 31. Гладкие мышцы располагаются в стенках внутренних полых органов: Сосудов Верхних дыхательных путей Желудочно-кишечного тракта Мочевого пузыря
- 32. 1) Миофибриллы с саркомерами располагаются нерегулярно. 2) Другой механизм сокращения (участвует внешний Са++). 3) Сокращение развивается
- 33. Способность к автоматии. Низкий мембранный потенциал покоя 3. Наличие потенциалзависимых медленных Са-каналов на плазматической мембране. 4.
- 34. Сравнительная характеристика скелетных и гладких мышц
- 36. Скачать презентацию

Слайд 3ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ
Двигательная.
Обменная – в мышцах происходят биохимические процессы, приводящие к изменению
ФУНКЦИИ СКЕЛЕТНЫХ МЫШЦ
Двигательная.
Обменная – в мышцах происходят биохимические процессы, приводящие к изменению

Слайд 4ТИПЫ СКЕЛЕТНЫХ МЫШЦ
Параллельно-
волокнистый
Перистые
Веретено-образный
ТИПЫ СКЕЛЕТНЫХ МЫШЦ
Параллельно-
волокнистый
Перистые
Веретено-образный

Слайд 5СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
Физические:
Эластичность – способность изменять длину под действием деформирующей силы
СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
Физические:
Эластичность – способность изменять длину под действием деформирующей силы

Слайд 6Мышца состоит из клеток – мышечных волокон.
Мышечные волокна скелетных мышц иннервируются мотонейронами.
Двигательная
Мышца состоит из клеток – мышечных волокон.
Мышечные волокна скелетных мышц иннервируются мотонейронами.
Двигательная
Слайд 7Электромиограмма: отражает возбуждение множества двигательных единиц.
Электромиограмма: отражает возбуждение множества двигательных единиц.
Слайд 8Нервно-мышечный синапс скелетных мышц (концевая пластинка)
Синапс между окончанием аксона мотонейрона спинного мозга
Нервно-мышечный синапс скелетных мышц (концевая пластинка)
Синапс между окончанием аксона мотонейрона спинного мозга
Слайд 9 Синаптические зоны в концевой пластинке располо-жены линейно вдоль двигательной терминали.
Синаптические зоны в концевой пластинке располо-жены линейно вдоль двигательной терминали.
Слайд 10ПОТЕНЦИАЛ КОНЦЕВОЙ ПЛАСТИНКИ
В нервно-мышечном синапсе на постсинаптической мембране возникает потенциал концевой пластинки
ПОТЕНЦИАЛ КОНЦЕВОЙ ПЛАСТИНКИ
В нервно-мышечном синапсе на постсинаптической мембране возникает потенциал концевой пластинки
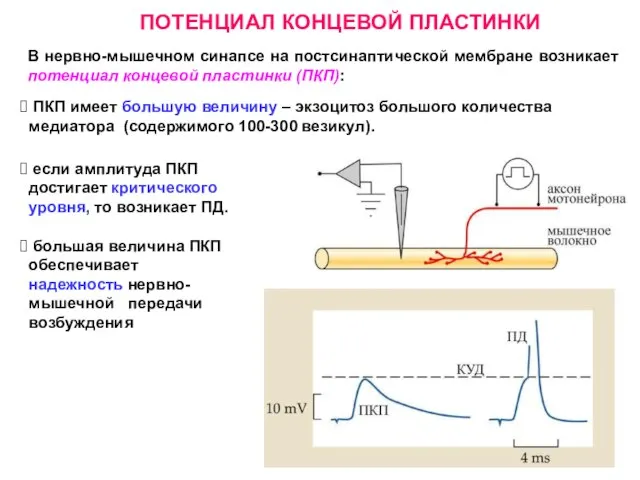
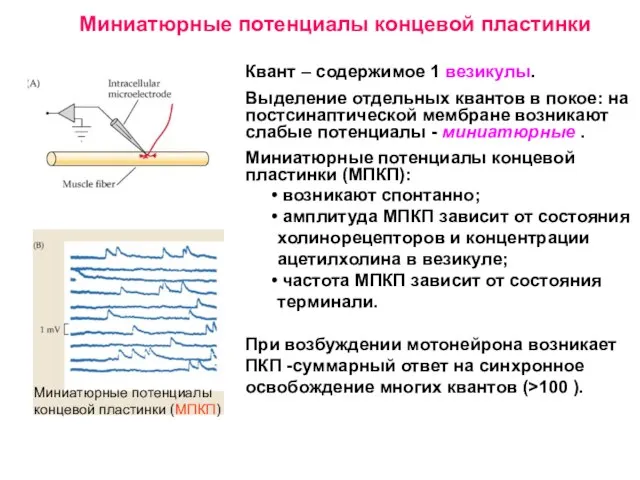
Слайд 11Квант – содержимое 1 везикулы.
Выделение отдельных квантов в покое: на постсинаптической мембране
Квант – содержимое 1 везикулы.
Выделение отдельных квантов в покое: на постсинаптической мембране
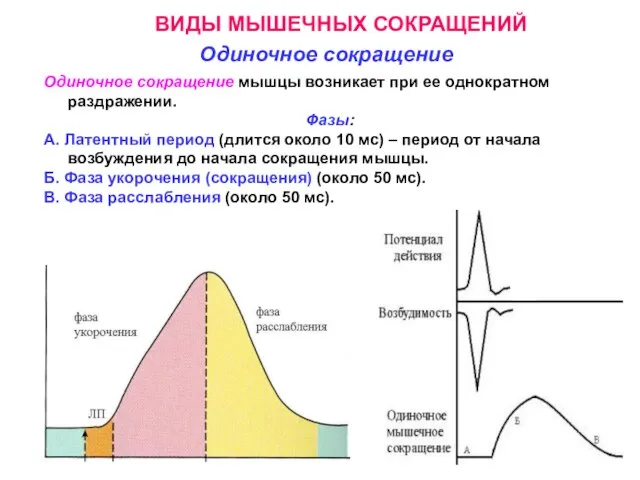
Слайд 12ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
Одиночное сокращение
Одиночное сокращение мышцы возникает при ее однократном раздражении.
ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
Одиночное сокращение
Одиночное сокращение мышцы возникает при ее однократном раздражении.
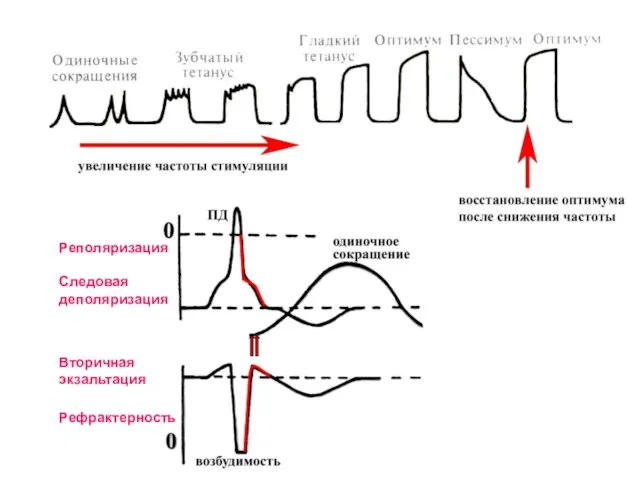
Слайд 13Тетаническое сокращение (тетанус) – суммация сокращений мышцы при действии ритмических раздражителей.
Виды тетануса:
Тетаническое сокращение (тетанус) – суммация сокращений мышцы при действии ритмических раздражителей.
Виды тетануса:
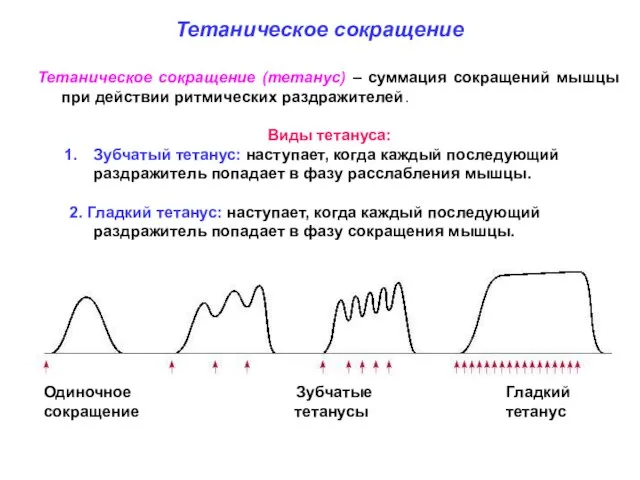
Слайд 14Следовая
деполяризация
Вторичная
экзальтация
Реполяризация
Рефрактерность
Следовая
деполяризация
Вторичная
экзальтация
Реполяризация
Рефрактерность
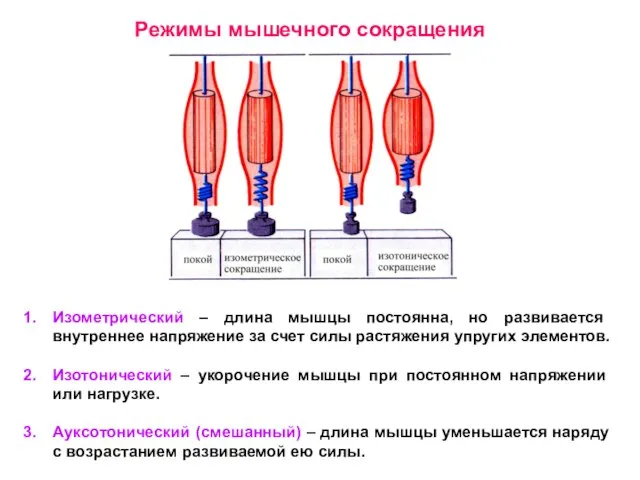
Слайд 15Режимы мышечного сокращения
Изометрический – длина мышцы постоянна, но развивается внутреннее напряжение
Режимы мышечного сокращения
Изометрический – длина мышцы постоянна, но развивается внутреннее напряжение
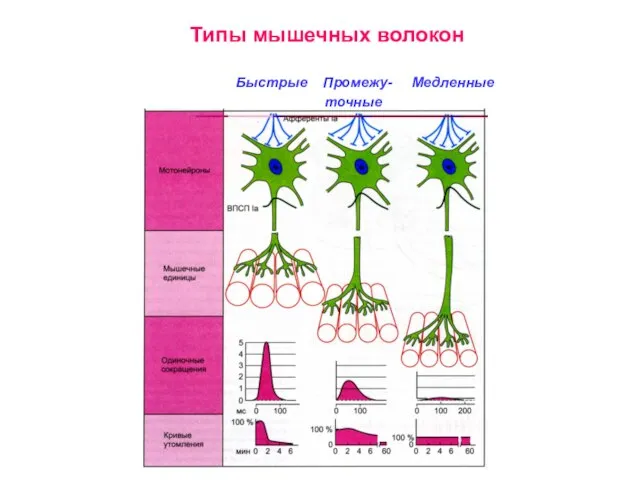
Слайд 16Типы мышечных волокон
Типы мышечных волокон
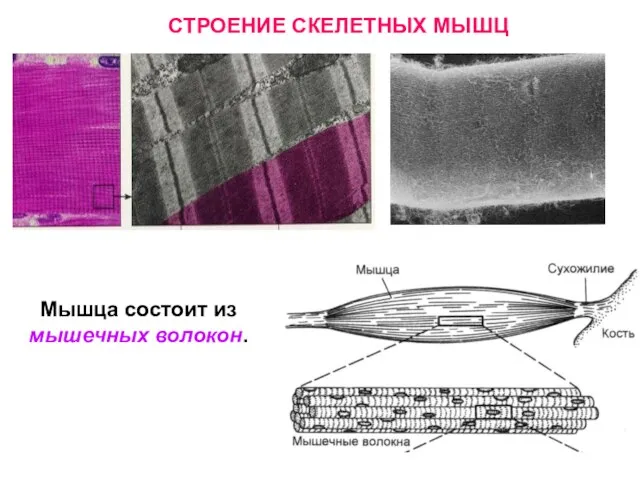
Слайд 17Мышца состоит из мышечных волокон.
СТРОЕНИЕ СКЕЛЕТНЫХ МЫШЦ
Мышца состоит из мышечных волокон.
СТРОЕНИЕ СКЕЛЕТНЫХ МЫШЦ
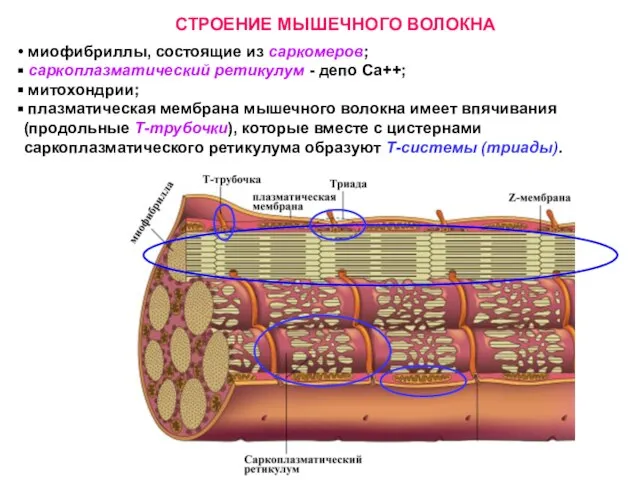
Слайд 18СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА
миофибриллы, состоящие из саркомеров;
саркоплазматический ретикулум - депо
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА
миофибриллы, состоящие из саркомеров;
саркоплазматический ретикулум - депо
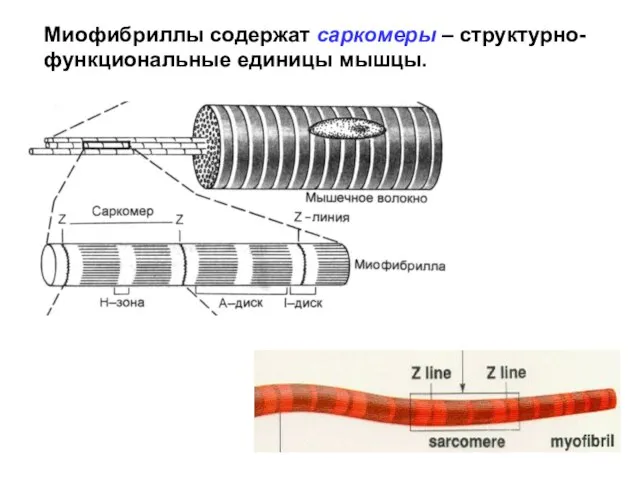
Слайд 19Миофибриллы содержат саркомеры – структурно-функциональные единицы мышцы.
Миофибриллы содержат саркомеры – структурно-функциональные единицы мышцы.
Слайд 20СТРОЕНИЕ САРКОМЕРА МЫШЕЧНОГО ВОЛОКНА
СТРОЕНИЕ САРКОМЕРА МЫШЕЧНОГО ВОЛОКНА
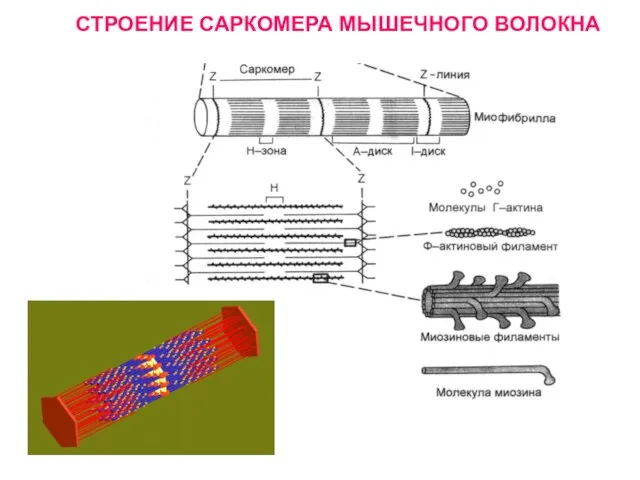
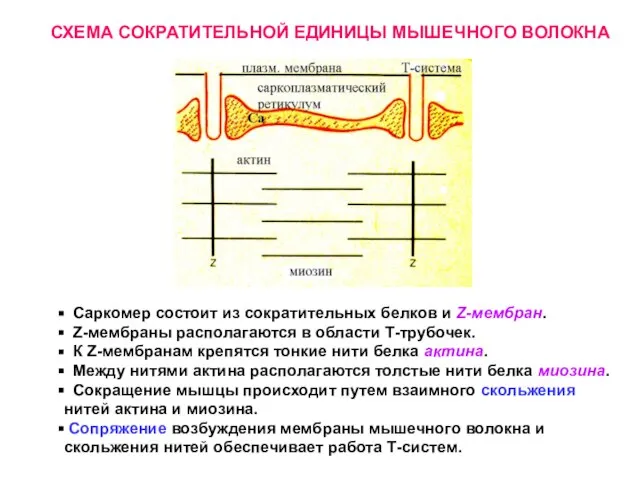
Слайд 21 Саркомер состоит из сократительных белков и Z-мембран.
Z-мембраны располагаются в
Саркомер состоит из сократительных белков и Z-мембран.
Z-мембраны располагаются в
Слайд 22Строение актина и миозина
Миозиновые нити имеют выступы – поперечные мостики. На их
Строение актина и миозина
Миозиновые нити имеют выступы – поперечные мостики. На их
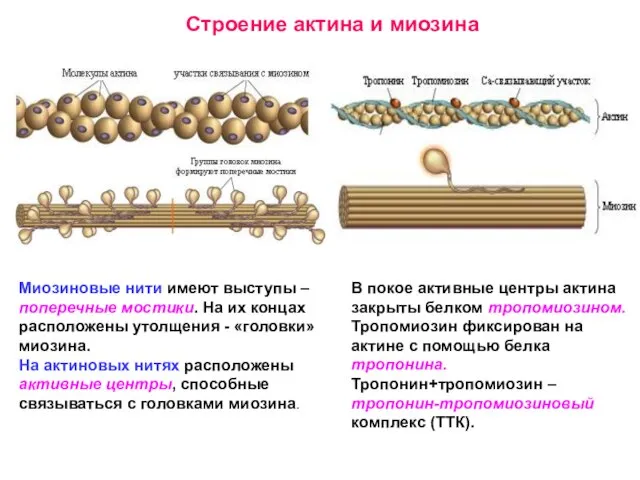
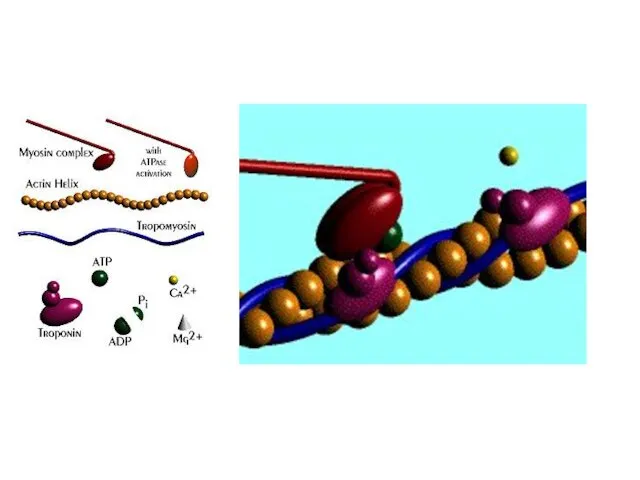
Слайд 23Механизм скольжения нитей актина и миозина.
Электромеханическое сопряжение.
Ион Са++ – посредник между
Механизм скольжения нитей актина и миозина.
Электромеханическое сопряжение.
Ион Са++ – посредник между
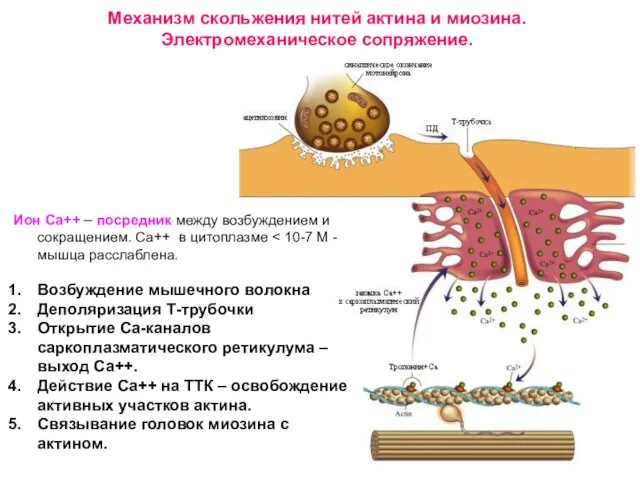
Слайд 24Механизм скольжения нитей: укорочение саркомера
6. Гидролиз АТФ головки миозина, отсоединение АДФ и
Механизм скольжения нитей: укорочение саркомера
6. Гидролиз АТФ головки миозина, отсоединение АДФ и
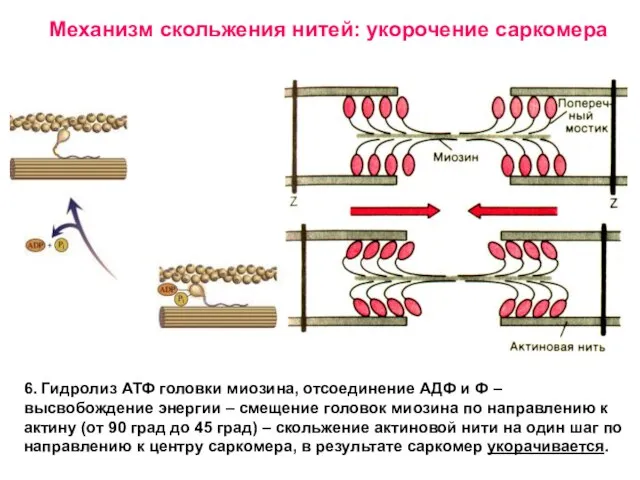
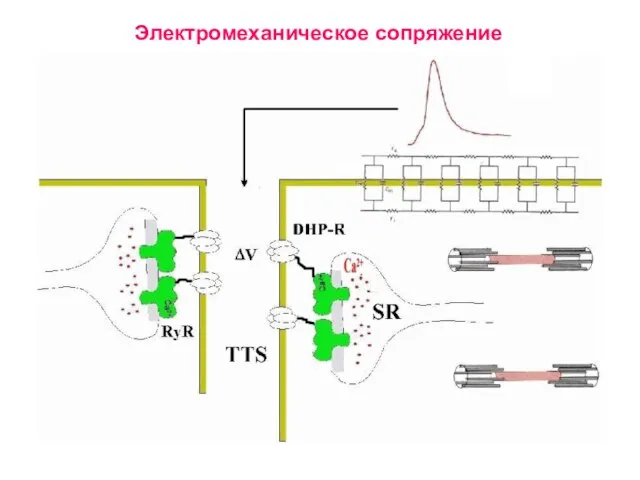
Слайд 25Электромеханическое сопряжение
Электромеханическое сопряжение
Слайд 26Поперечный мостик распрямляется, головка миозина присоединяется к следующему активному центру актина.
Механизм
Поперечный мостик распрямляется, головка миозина присоединяется к следующему активному центру актина.
Механизм
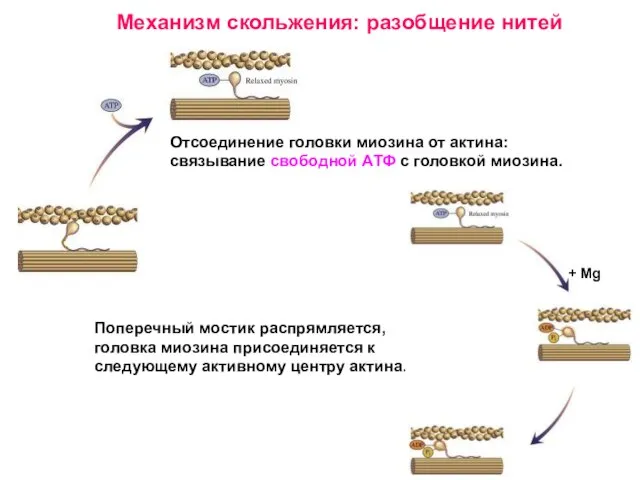
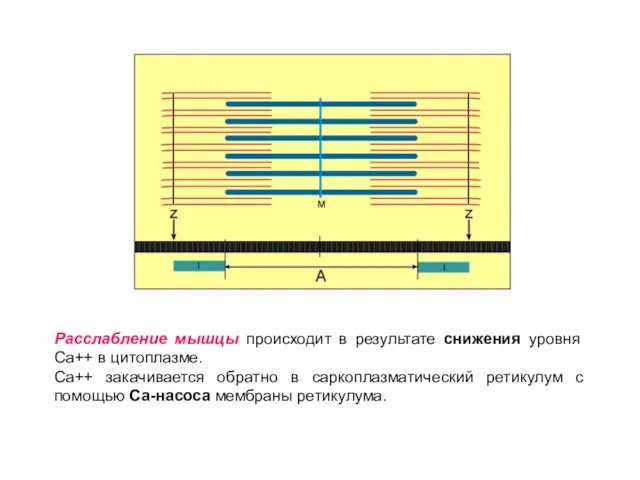
Слайд 28Расслабление мышцы происходит в результате снижения уровня Са++ в цитоплазме.
Са++ закачивается
Расслабление мышцы происходит в результате снижения уровня Са++ в цитоплазме.
Са++ закачивается
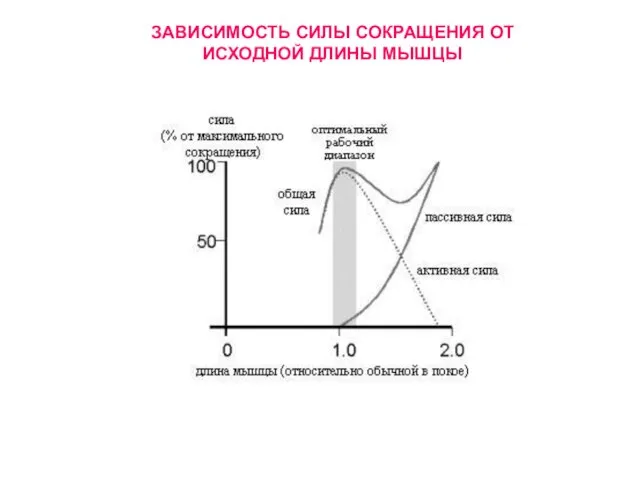
Слайд 29ЗАВИСИМОСТЬ СИЛЫ СОКРАЩЕНИЯ ОТ ИСХОДНОЙ ДЛИНЫ МЫШЦЫ
ЗАВИСИМОСТЬ СИЛЫ СОКРАЩЕНИЯ ОТ ИСХОДНОЙ ДЛИНЫ МЫШЦЫ
Слайд 30ФИЗИОЛОГИЯ
ГЛАДКИХ
МЫШЦ
ФИЗИОЛОГИЯ
ГЛАДКИХ
МЫШЦ

Слайд 31Гладкие мышцы располагаются в стенках внутренних полых органов:
Сосудов
Верхних дыхательных путей
Гладкие мышцы располагаются в стенках внутренних полых органов:
Сосудов
Верхних дыхательных путей
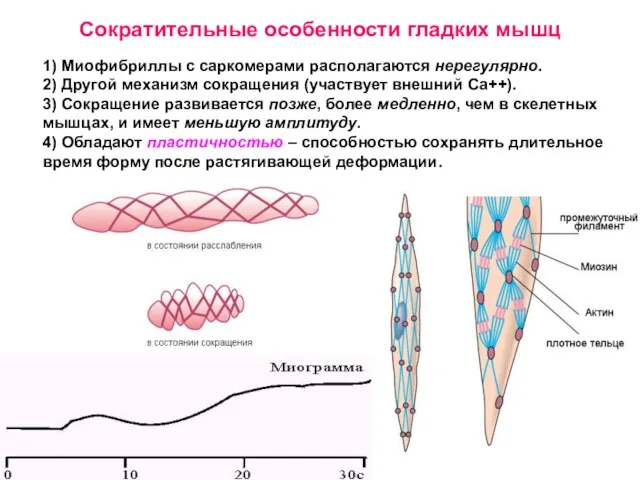
Слайд 321) Миофибриллы с саркомерами располагаются нерегулярно.
2) Другой механизм сокращения (участвует внешний Са++).
3)
1) Миофибриллы с саркомерами располагаются нерегулярно.
2) Другой механизм сокращения (участвует внешний Са++).
3)
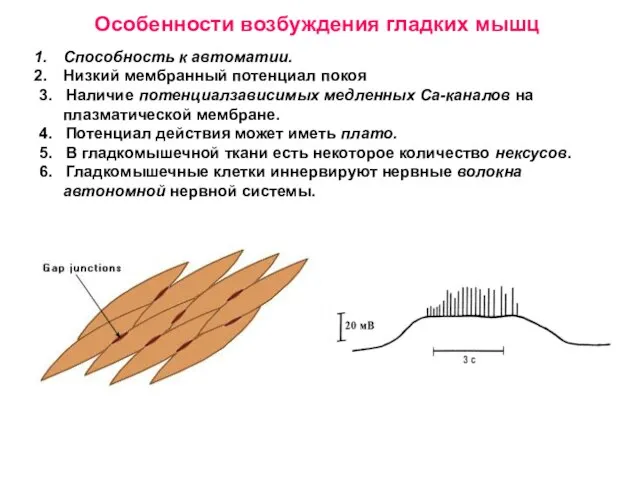
Слайд 33Способность к автоматии.
Низкий мембранный потенциал покоя
3. Наличие потенциалзависимых медленных Са-каналов на
Способность к автоматии.
Низкий мембранный потенциал покоя
3. Наличие потенциалзависимых медленных Са-каналов на
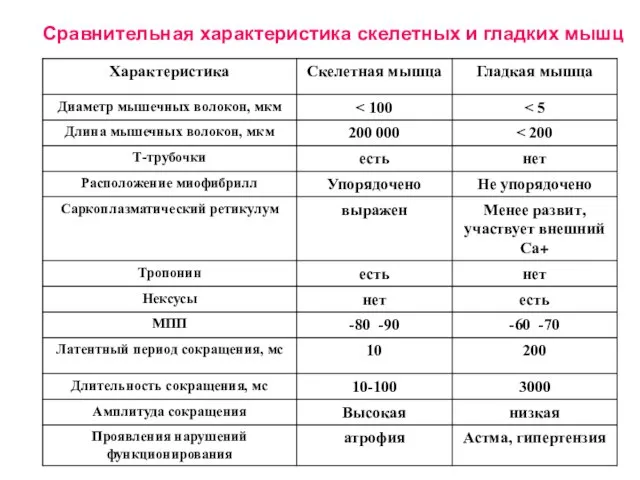
Слайд 34Сравнительная характеристика скелетных и гладких мышц
Сравнительная характеристика скелетных и гладких мышц
 Свобода и ответственность (4 класс)
Свобода и ответственность (4 класс) Презентация на тему Новый год. История праздника
Презентация на тему Новый год. История праздника  Загрязнение автотранспортом
Загрязнение автотранспортом Муниципальное автономное дошкольное образовательное учреждение «Центр развития ребенка – детский сад №50 «Родничок»б
Муниципальное автономное дошкольное образовательное учреждение «Центр развития ребенка – детский сад №50 «Родничок»б Энергетическое использование биомассы в Украине: правовые аспекты
Энергетическое использование биомассы в Украине: правовые аспекты Приоритетными направлениями деятельности общественного движения «Добрые дети мира» в 2010 году будут являться:
Приоритетными направлениями деятельности общественного движения «Добрые дети мира» в 2010 году будут являться: Воркшоп по Мастерской будущего Содержание
Воркшоп по Мастерской будущего Содержание Учителя – участники ВОВ
Учителя – участники ВОВ Цитомегаловирусы
Цитомегаловирусы Политическое лидерство: Дональд Трамп
Политическое лидерство: Дональд Трамп Всегда великая Россия
Всегда великая Россия Подготовка к ГИА по русскому языку часть 2 (тестовые задания и задания с кратким ответом)
Подготовка к ГИА по русскому языку часть 2 (тестовые задания и задания с кратким ответом) Биологическая и психологическая подструктура личности
Биологическая и психологическая подструктура личности Старая печатная русскоязычная реклама парфюмерии
Старая печатная русскоязычная реклама парфюмерии Тема 12.Правонарушения
Тема 12.Правонарушения Творения мира
Творения мира Коллективное управление смежными правами Глава III. Смежные права (Закон РК «Об авторском праве и смежных правах» от 10 июня 1996 года N
Коллективное управление смежными правами Глава III. Смежные права (Закон РК «Об авторском праве и смежных правах» от 10 июня 1996 года N OOP, metaprogramming, blocks, iterators
OOP, metaprogramming, blocks, iterators Независимый депозитарий с сегрегированными счетами и арбитражем сделок для трейдеров и брокеров
Независимый депозитарий с сегрегированными счетами и арбитражем сделок для трейдеров и брокеров Мат одинокому королю
Мат одинокому королю Панно средствами народного примитивного орнаментирования
Панно средствами народного примитивного орнаментирования Имя урока
Имя урока Установка локальной сети в школе
Установка локальной сети в школе Образовательный проект Промсвязьбанка
Образовательный проект Промсвязьбанка Презентация на тему Тюменская область история, география, экономика
Презентация на тему Тюменская область история, география, экономика Санаторий Лесники. Медицинские услуги
Санаторий Лесники. Медицинские услуги Опыт взаимодействия Церкви и государства в трезвенном просвещении
Опыт взаимодействия Церкви и государства в трезвенном просвещении Чытачам Брылеўскай бібліятэкі Красавік 2019 г
Чытачам Брылеўскай бібліятэкі Красавік 2019 г