- Кроветворение гемопоэз

Содержание
- 2. Кроветворение (гемопоэз) – процесс образования крови. Выделяют эмбриональный и постэмбриональный гемопоэз. Эмбриональный гемопоэз – это процесс
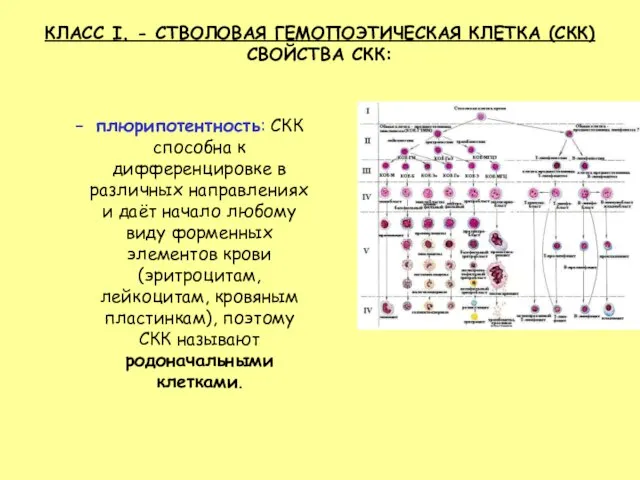
- 3. Согласно унитарной теории кроветворения, все клетки крови развиваются из одной родоначальной стволовой кроветворной клетки (СКК).
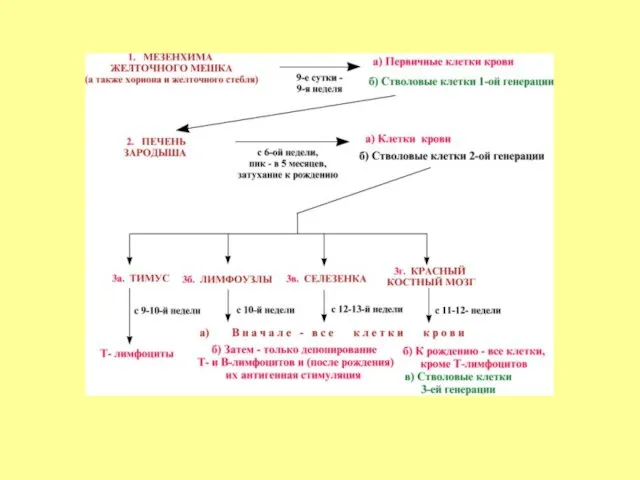
- 4. Эмбриональный гемопоэз делится на три периода в зависимости от времени и места протекания. Это периоды в
- 5. Мегалобластический период начинается со 2-3 недели внутриутробной жизни в мезенхиме желточного мешка. В результате интенсивного деления
- 6. Большая часть бластов делится и превращается в первичные эритробласты больших размеров – мегалобласты. Мегалобласты активно делятся
- 7. Одновременно с эритропоэзом в желточном мешке экстраваскулярно – вне просвета сосудов - идёт гранулоцитопоэз – образуются
- 8. В печени кроветворение начинается на 5-6 неделях развития. Здесь образуются эритроциты, гранулоциты и тромбоциты. К концу
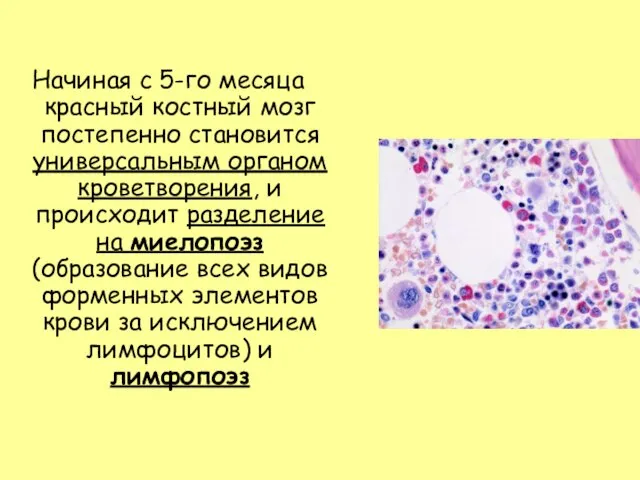
- 9. Начиная с 5-го месяца красный костный мозг постепенно становится универсальным органом кроветворения, и происходит разделение на
- 10. Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической и репаративной регенерации после рождения.
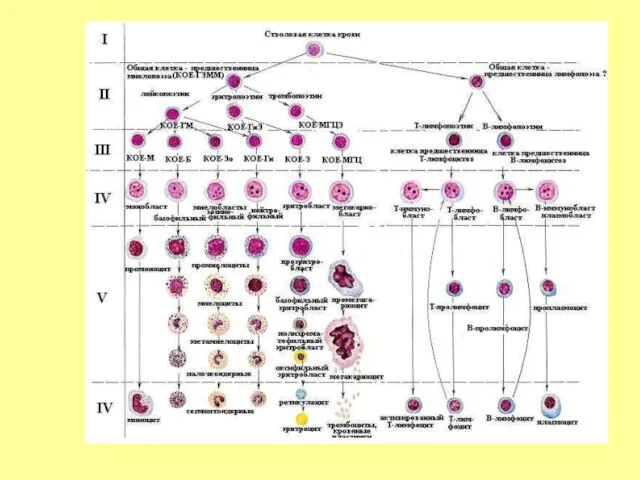
- 11. Современные представления о кроветворении основаны на признании унитарной теории кроветворения. Согласно этой теории, развитие всех клеток
- 13. КЛАСС I. - СТВОЛОВАЯ ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА (СКК) СВОЙСТВА СКК: плюрипотентность: СКК способна к дифференцировке в различных
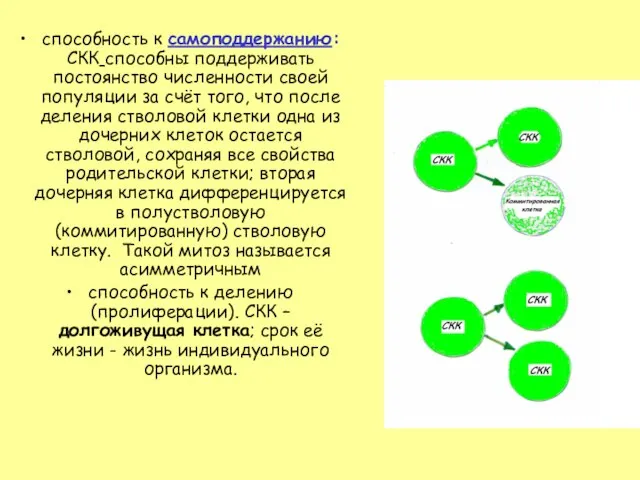
- 14. способность к самоподдержанию: СКК способны поддерживать постоянство численности своей популяции за счёт того, что после деления
- 15. устойчивость к действию повреждающих факторов, вероятно вследствие того, что СКК делятся редко; большую часть своей жизни
- 16. морфологически СКК не идентифицируются: то есть их нельзя различить обычными методами под световым или электронным микроскопом,
- 17. основное место локализации СКК – красный костный мозг, хотя численность СКК невелика (1 СКК на 2000
- 18. II. КЛАСС - МУЛЬТИПОТЕНТНЫЕ КОММИТИРОВАННЫЕ, частично детерминированные (ПОЛУСТВОЛОВЫЕ) КЛЕТКИ Мультипотентные коммитированные клетки дают начало форменным элементам
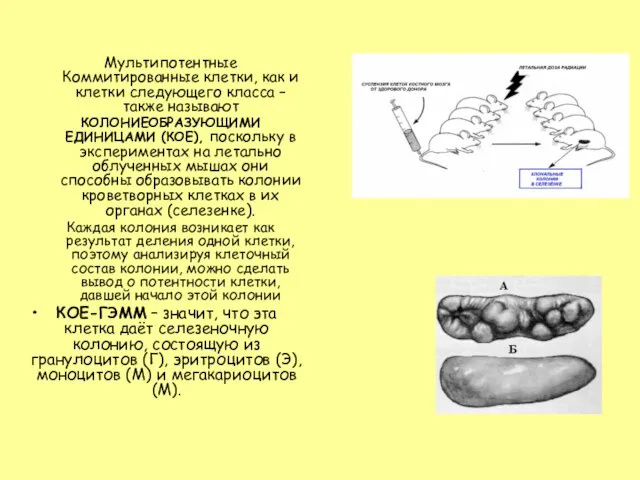
- 19. Мультипотентные Коммитированные клетки, как и клетки следующего класса – также называют КОЛОНИЕОБРАЗУЮЩИМИ ЕДИНИЦАМИ (КОЕ), поскольку в
- 20. КЛАСС III. УНИПОТЕНТНЫЕ (КОММИТИРОВАННЫЕ) РОДОНАЧАЛЬНЫЕ КЛЕТКИ (ПРОГЕНИТОРНЫЕ, PROGENITORS) унипотентны - детерминированы в направлении развития только одного
- 21. IY. КЛЕТКИ-ПРЕДШЕСТВЕННИКИ (БЛАСТЫ, PRECURSORS) представляют отдельные линии развития форменных элементов; пролиферативная активность ограничена, но выше, чем
- 22. Y.Созревающие клетки Подвергаются структурной и функциональной дифференцировке, в ходе которой утрачивают способность к делению (за исключением
- 23. ДИФФЕРОН - совокупность всех клеток, составляющих ту или иную линию дифференцировки от стволовых (наименее дифференцированных) клеток
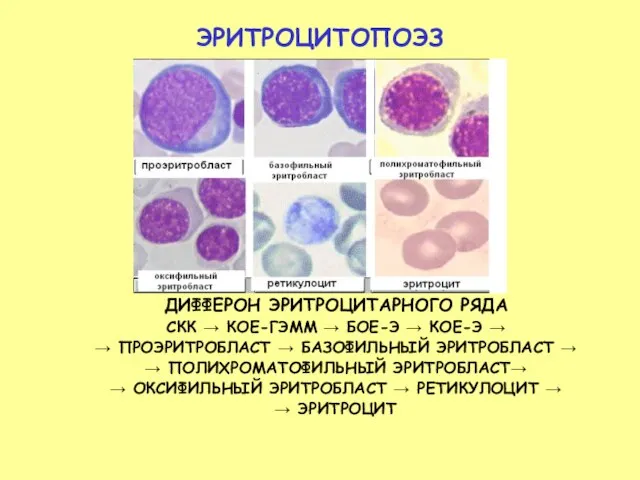
- 24. ЭРИТРОЦИТОПОЭЗ ДИФФЕРОН ЭРИТРОЦИТАРНОГО РЯДА СКК → КОЕ-ГЭММ → БОЕ-Э → КОЕ-Э → → ПРОЭРИТРОБЛАСТ → БАЗОФИЛЬНЫЙ
- 25. Начало эритроидного ряда – взрывообразующая единица эритропоэза – BFU-E. При активации и делении BFU-E образуется множество
- 26. Из проэритробласта последовательно образуются: базофильный эритробласт (накопление рибосом и начало синтеза Hb); полихроматофильный эритробласт (накопление Hb);
- 27. При дифференцировке предшественников эритроцитов в зрелые эритроциты происходят следующие процессы: уменьшение размеров клетки; выработка и накопление
- 28. Гранулоцитопоэз СКК → КОЕ-ГЭММ → КОЕ-ГнМ → КОЕ-Гн → МИЕЛОБЛАСТ → ПРОМИЕЛОЦИТ → МИЕЛОЦИТ → →
- 29. Гранулоциты при развитии проходят следующие стадии: миелобласт (не имеет гранул) промиелоцит (первичные, азурофильные гранулы) миелоцит (появление
- 30. По мере созревания гранулоцитов в зрелые клетки происходит: уменьшение размеров клетки; изменение формы их ядер от
- 31. Тромбоцитопоэз – процесс образования и созревания тромбоцитов происходит в миелоидной ткани. Тромбоциты (кровяные пластинки) образуются в
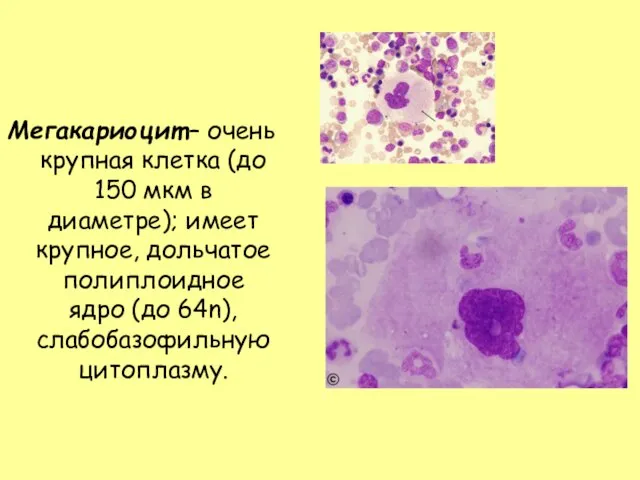
- 32. Мегакариоцит – очень крупная клетка (до 150 мкм в диаметре); имеет крупное, дольчатое полиплоидное ядро (до
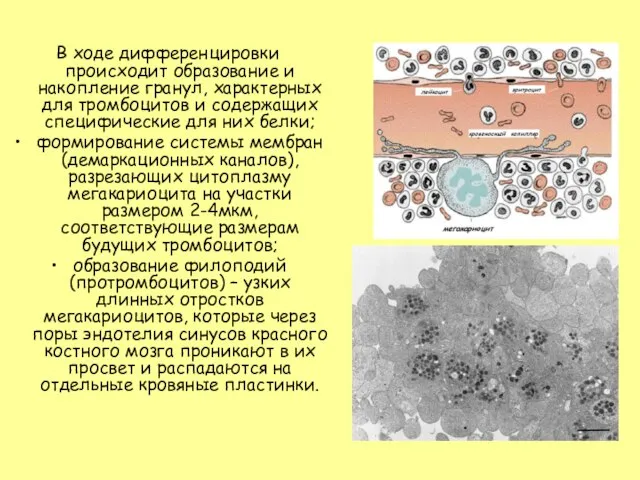
- 33. В ходе дифференцировки происходит образование и накопление гранул, характерных для тромбоцитов и содержащих специфические для них
- 36. Скачать презентацию
Слайд 2Кроветворение (гемопоэз) – процесс образования крови.
Выделяют эмбриональный и постэмбриональный гемопоэз.
Эмбриональный
Кроветворение (гемопоэз) – процесс образования крови.
Выделяют эмбриональный и постэмбриональный гемопоэз.
Эмбриональный

Слайд 3
Согласно унитарной теории кроветворения, все клетки крови развиваются из одной родоначальной стволовой
Согласно унитарной теории кроветворения, все клетки крови развиваются из одной родоначальной стволовой

Слайд 4Эмбриональный гемопоэз делится на три периода в зависимости от времени и места
Эмбриональный гемопоэз делится на три периода в зависимости от времени и места
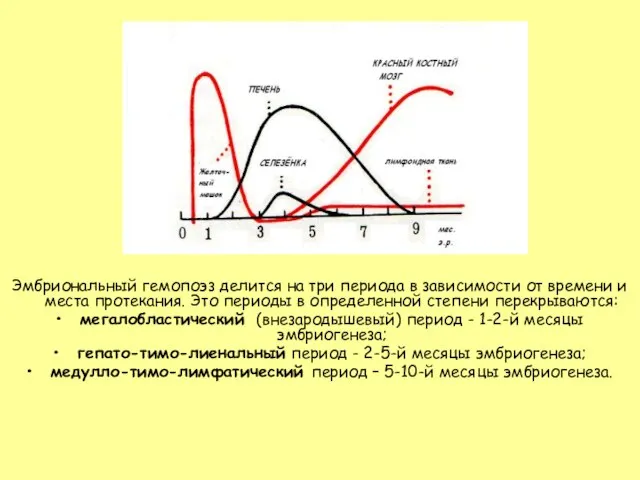
Слайд 5Мегалобластический период начинается со 2-3 недели внутриутробной жизни в мезенхиме желточного мешка.
Мегалобластический период начинается со 2-3 недели внутриутробной жизни в мезенхиме желточного мешка.
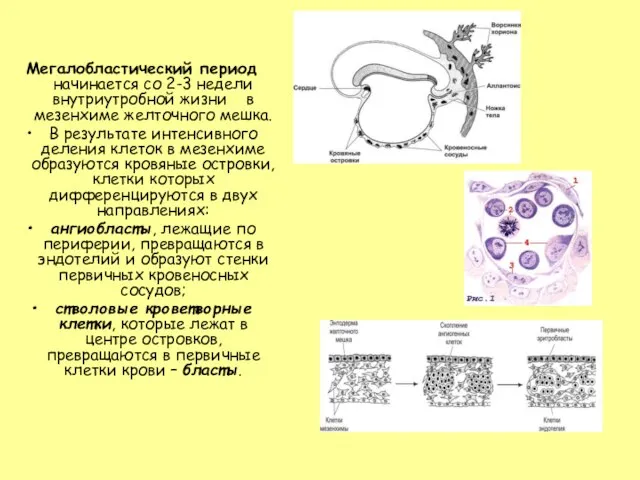
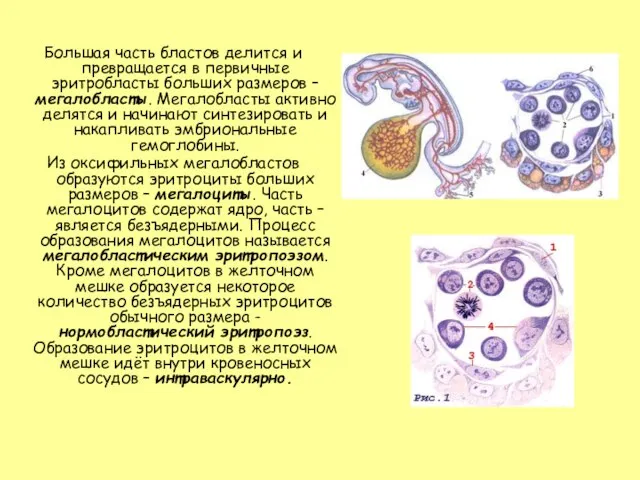
Слайд 6Большая часть бластов делится и превращается в первичные эритробласты больших размеров –
Большая часть бластов делится и превращается в первичные эритробласты больших размеров –
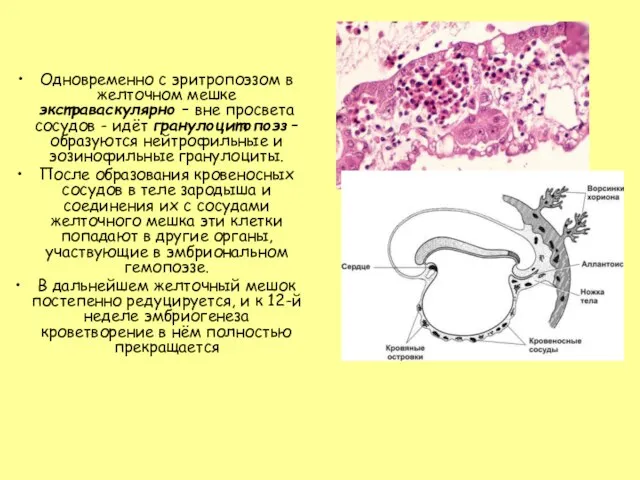
Слайд 7Одновременно с эритропоэзом в желточном мешке экстраваскулярно – вне просвета сосудов -
Одновременно с эритропоэзом в желточном мешке экстраваскулярно – вне просвета сосудов -
Слайд 8В печени кроветворение начинается на 5-6 неделях развития. Здесь образуются эритроциты, гранулоциты
В печени кроветворение начинается на 5-6 неделях развития. Здесь образуются эритроциты, гранулоциты

Слайд 9Начиная с 5-го месяца красный костный мозг постепенно становится универсальным органом кроветворения,
Начиная с 5-го месяца красный костный мозг постепенно становится универсальным органом кроветворения,
Слайд 10Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической и
Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической и

Слайд 11Современные представления о кроветворении основаны на признании унитарной теории кроветворения. Согласно этой
Современные представления о кроветворении основаны на признании унитарной теории кроветворения. Согласно этой

Слайд 13КЛАСС I. - СТВОЛОВАЯ ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА (СКК)
СВОЙСТВА СКК:
плюрипотентность: СКК способна к дифференцировке
КЛАСС I. - СТВОЛОВАЯ ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА (СКК)
СВОЙСТВА СКК:
плюрипотентность: СКК способна к дифференцировке
Слайд 14способность к самоподдержанию: СКК способны поддерживать постоянство численности своей популяции за счёт
способность к самоподдержанию: СКК способны поддерживать постоянство численности своей популяции за счёт
Слайд 15устойчивость к действию повреждающих факторов, вероятно вследствие того, что СКК делятся редко;
устойчивость к действию повреждающих факторов, вероятно вследствие того, что СКК делятся редко;

Слайд 16морфологически СКК не идентифицируются: то есть их нельзя различить обычными методами под
морфологически СКК не идентифицируются: то есть их нельзя различить обычными методами под

Слайд 17основное место локализации СКК – красный костный мозг, хотя численность СКК невелика
основное место локализации СКК – красный костный мозг, хотя численность СКК невелика

Слайд 18II. КЛАСС - МУЛЬТИПОТЕНТНЫЕ КОММИТИРОВАННЫЕ, частично детерминированные (ПОЛУСТВОЛОВЫЕ) КЛЕТКИ
Мультипотентные коммитированные клетки дают
II. КЛАСС - МУЛЬТИПОТЕНТНЫЕ КОММИТИРОВАННЫЕ, частично детерминированные (ПОЛУСТВОЛОВЫЕ) КЛЕТКИ
Мультипотентные коммитированные клетки дают

Слайд 19Мультипотентные Коммитированные клетки, как и клетки следующего класса – также называют
КОЛОНИЕОБРАЗУЮЩИМИ ЕДИНИЦАМИ
Мультипотентные Коммитированные клетки, как и клетки следующего класса – также называют
КОЛОНИЕОБРАЗУЮЩИМИ ЕДИНИЦАМИ
Слайд 20КЛАСС III. УНИПОТЕНТНЫЕ (КОММИТИРОВАННЫЕ) РОДОНАЧАЛЬНЫЕ КЛЕТКИ (ПРОГЕНИТОРНЫЕ, PROGENITORS)
унипотентны - детерминированы в направлении
КЛАСС III. УНИПОТЕНТНЫЕ (КОММИТИРОВАННЫЕ) РОДОНАЧАЛЬНЫЕ КЛЕТКИ (ПРОГЕНИТОРНЫЕ, PROGENITORS)
унипотентны - детерминированы в направлении
Слайд 21IY. КЛЕТКИ-ПРЕДШЕСТВЕННИКИ (БЛАСТЫ, PRECURSORS)
представляют отдельные линии развития форменных элементов;
пролиферативная активность ограничена, но
IY. КЛЕТКИ-ПРЕДШЕСТВЕННИКИ (БЛАСТЫ, PRECURSORS)
представляют отдельные линии развития форменных элементов;
пролиферативная активность ограничена, но
Слайд 22Y.Созревающие клетки
Подвергаются структурной и функциональной дифференцировке, в ходе которой утрачивают способность к
Y.Созревающие клетки
Подвергаются структурной и функциональной дифференцировке, в ходе которой утрачивают способность к
Слайд 23ДИФФЕРОН - совокупность всех клеток, составляющих ту или иную линию дифференцировки от
ДИФФЕРОН - совокупность всех клеток, составляющих ту или иную линию дифференцировки от

Слайд 24ЭРИТРОЦИТОПОЭЗ
ДИФФЕРОН ЭРИТРОЦИТАРНОГО РЯДА
СКК → КОЕ-ГЭММ → БОЕ-Э → КОЕ-Э →
→ ПРОЭРИТРОБЛАСТ
ЭРИТРОЦИТОПОЭЗ
ДИФФЕРОН ЭРИТРОЦИТАРНОГО РЯДА
СКК → КОЕ-ГЭММ → БОЕ-Э → КОЕ-Э →
→ ПРОЭРИТРОБЛАСТ
Слайд 25Начало эритроидного ряда – взрывообразующая единица эритропоэза – BFU-E.
При активации и
Начало эритроидного ряда – взрывообразующая единица эритропоэза – BFU-E.
При активации и

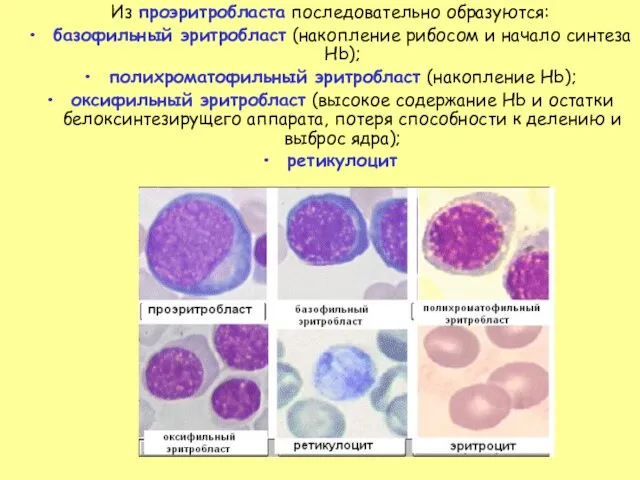
Слайд 26Из проэритробласта последовательно образуются:
базофильный эритробласт (накопление рибосом и начало синтеза Hb);
полихроматофильный эритробласт
Из проэритробласта последовательно образуются:
базофильный эритробласт (накопление рибосом и начало синтеза Hb);
полихроматофильный эритробласт
Слайд 27При дифференцировке предшественников эритроцитов в зрелые эритроциты происходят следующие процессы:
уменьшение размеров клетки;
выработка
При дифференцировке предшественников эритроцитов в зрелые эритроциты происходят следующие процессы:
уменьшение размеров клетки;
выработка

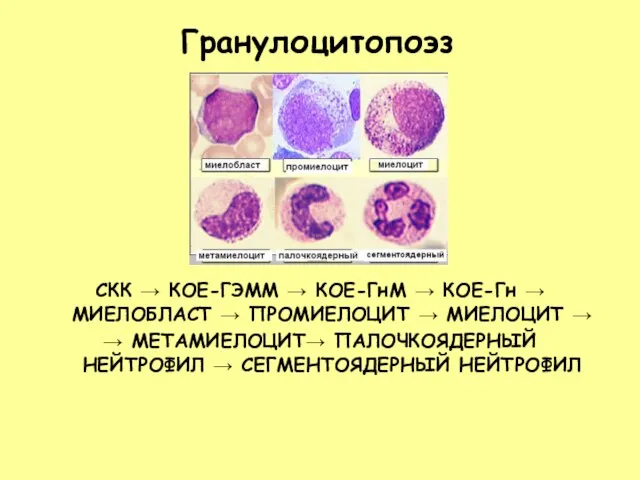
Слайд 28Гранулоцитопоэз
СКК → КОЕ-ГЭММ → КОЕ-ГнМ → КОЕ-Гн → МИЕЛОБЛАСТ → ПРОМИЕЛОЦИТ →
Гранулоцитопоэз
СКК → КОЕ-ГЭММ → КОЕ-ГнМ → КОЕ-Гн → МИЕЛОБЛАСТ → ПРОМИЕЛОЦИТ →
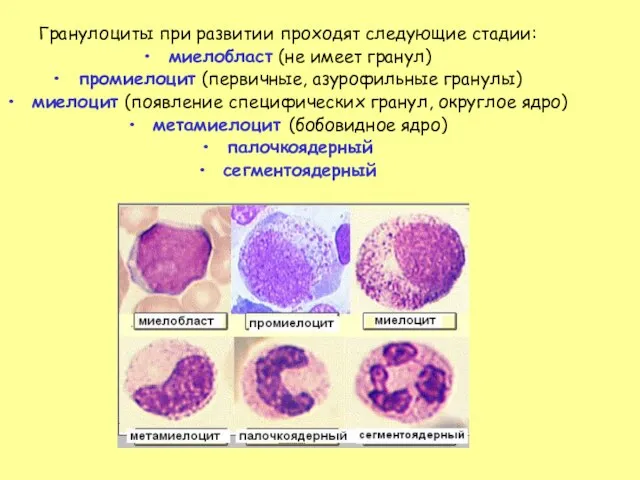
Слайд 29Гранулоциты при развитии проходят следующие стадии:
миелобласт (не имеет гранул)
промиелоцит (первичные, азурофильные гранулы)
миелоцит
Гранулоциты при развитии проходят следующие стадии:
миелобласт (не имеет гранул)
промиелоцит (первичные, азурофильные гранулы)
миелоцит
Слайд 30По мере созревания гранулоцитов в зрелые клетки происходит:
уменьшение размеров клетки;
изменение формы их
По мере созревания гранулоцитов в зрелые клетки происходит:
уменьшение размеров клетки;
изменение формы их

Слайд 31Тромбоцитопоэз – процесс образования и созревания тромбоцитов происходит в миелоидной ткани. Тромбоциты
Тромбоцитопоэз – процесс образования и созревания тромбоцитов происходит в миелоидной ткани. Тромбоциты

Слайд 32Мегакариоцит – очень крупная клетка (до 150 мкм в диаметре); имеет крупное,
Мегакариоцит – очень крупная клетка (до 150 мкм в диаметре); имеет крупное,
Слайд 33В ходе дифференцировки происходит образование и накопление гранул, характерных для тромбоцитов и
В ходе дифференцировки происходит образование и накопление гранул, характерных для тромбоцитов и
 Подпись/расшифровка АБИЭ
Подпись/расшифровка АБИЭ Презентация на тему жанр рецензии
Презентация на тему жанр рецензии  Презентация на тему Авиационная промышленность. Рынок гражданской авиации
Презентация на тему Авиационная промышленность. Рынок гражданской авиации  Результаты маркетингового исследования
Результаты маркетингового исследования Результаты опроса молодежи во время концерта, посвященного проблемам ВИЧ/СПИД в Украине
Результаты опроса молодежи во время концерта, посвященного проблемам ВИЧ/СПИД в Украине Lektsia_po_stipendialnomu_obespecheniyu (1)
Lektsia_po_stipendialnomu_obespecheniyu (1) ЗАКОНЫ И ДРУГИЕ НОРМАТИВНО-ПРАВОВЫЕ АКТЫ РОССИЙСКОЙ ФЕДЕРАЦИИ ПО ОБЕСПЕЧЕНИЮ БЕЗОПАСНОСТИ
ЗАКОНЫ И ДРУГИЕ НОРМАТИВНО-ПРАВОВЫЕ АКТЫ РОССИЙСКОЙ ФЕДЕРАЦИИ ПО ОБЕСПЕЧЕНИЮ БЕЗОПАСНОСТИ ЕЛКИ - 2
ЕЛКИ - 2 Служба по контракту
Служба по контракту Рекомендации по оформлению и защите научноисследовательской работы. Оформление сопроводительных документов
Рекомендации по оформлению и защите научноисследовательской работы. Оформление сопроводительных документов Прошлое и настоящее цифровой фотограмметрии
Прошлое и настоящее цифровой фотограмметрии Народная глиняная игрушка
Народная глиняная игрушка Рекомендации по применению принципов usability в разработке веб-систем и управлении веб-системами
Рекомендации по применению принципов usability в разработке веб-систем и управлении веб-системами Основные концепции происхождения и сущности права
Основные концепции происхождения и сущности права Правила поведения в школе и на улице
Правила поведения в школе и на улице Шахматная нотация. Занятие 2
Шахматная нотация. Занятие 2 Презентация на тему Правовое воспитание дошкольников
Презентация на тему Правовое воспитание дошкольников Мировой опыт развития посылочной торговли
Мировой опыт развития посылочной торговли ВетераныВеликой Отечественной войнычасть 1
ВетераныВеликой Отечественной войнычасть 1 Правки. Лого полноценные
Правки. Лого полноценные 2019 декабрь ООО Жилкомсервис Кронштадтского района
2019 декабрь ООО Жилкомсервис Кронштадтского района Коренной перелом в ходе войны
Коренной перелом в ходе войны Работа в Excel 2007
Работа в Excel 2007 Choosing a career
Choosing a career Участие базальных ганглиев и коры больших полушарий
Участие базальных ганглиев и коры больших полушарий  Разнообразие животных Беспозвоночные
Разнообразие животных Беспозвоночные «Название проекта» ФИО докладчика
«Название проекта» ФИО докладчика Презентация на тему Государство на берегах Нила 5 класс
Презентация на тему Государство на берегах Нила 5 класс