- МОЛЕКУЛЯРНА БІОФІЗИКА

Содержание
- 2. Біофізика білків Білки – молекулярні “машини”. Структурна організація білків. Ферменти – як представники групи білків. Механізм
- 3. Білки – молекулярні “машини”
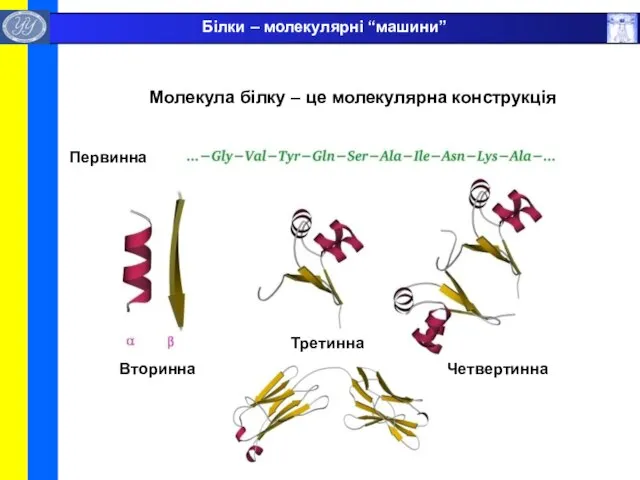
- 4. Молекула білку – це молекулярна конструкція Первинна Вторинна Третинна Четвертинна Білки – молекулярні “машини”
- 5. Атомарна та скелетна моделі молекули білка Білки – молекулярні “машини”
- 6. α-L-амінокислоти Структурна організація білків. Амінокислоти Первинна структура— послідовність амінокислот у ланцюзі полімеру, які пов'язані міцними ковалентними
- 7. Структурна організація білків. Амінокислоти
- 8. Структурна організація білків. Амінокислоти
- 9. Структурна організація білків. Амінокислоти
- 10. Структурна організація білків. Амінокислоти
- 11. Структурна організація білків. Амінокислоти
- 12. Структурна організація білків. Амінокислоти
- 13. Бокові радикали головних L-амінокислот, які складають природні білки. Структурна організація білків. Амінокислоти
- 14. Білки – молекулярні конструкції Амінокислоти Тирозин
- 15. Білки – молекулярні конструкції Амінокислоти Тирозин
- 16. Білки – молекулярні конструкції Амінокислоти Фенілаланін
- 17. Білки – молекулярні конструкції Амінокислоти Гістидин
- 18. Білки – молекулярні конструкції Амінокислоти Валін
- 19. Білки – молекулярні конструкції Амінокислоти Аспарагінова кислота
- 20. Білки – молекулярні конструкції Амінокислоти Глутамін
- 21. Методи дослідження амінокислот Хроматографічні. Електрофоретичні. Оптичні (поглинання і флуоресценція). Маспектрометричні. Структурна організація білків. Амінокислоти
- 22. Зв'язки, які забезпечують стабілізацію просторової структури біомакромолекул: ковалентні зв'язки; електростатичні взаємодії; водневі зв'язки; Ван-дер-Ваальсові взаємодії; гідрофобні
- 23. Рівні структурної організації білків Первинна структура (1º) - амінокислотна послідовність білків; Вторинна структура (2º) – сегменти,
- 24. Взаємодії в пептидному зв'язку Структурна організація білків
- 25. “Конструкція” пептидного зв'язку Обертання навколо одинарного зв'язку Обертання навколо пептидного зв'язку практично неможливо Структурна організація білків
- 26. Білки – молекулярні конструкції
- 27. Вторинна структура характеризує просторову форму білкової молекули, яка найчастіше повністю або частково закручується у спіраль. Амінокислотні
- 28. α-спіралі — щільні витки навколо довгої осі структури, один виток становлять 4 амінокислотних залишки, спіраль стабілізована
- 29. β-листи (складчасті шари) — декілька зигзагоподібних поліпептідних ланцюжків, в яких водневі зв'язки утворюються між відносно віддаленними
- 30. Лист β-структури Структурна організація білків
- 31. Приклади розташування β-тяжів
- 32. Білки – молекулярні конструкції β-структурні “мотиви” у білках
- 33. π-спіралі Невпорядкована структура Структурна організація білків
- 34. Розташування амінокислотних радикалів в α- спіралях та β-структурах та їх тенденції амінокислотних залишків до формування певних
- 35. «Надвторинна» структура Більшість білків організовані на кількох проміжних рівнях між основними елементами вторинної структури та повною
- 36. Елемент β-α-β Структурна організація білків
- 37. Типові варіанти (мотиви) просторової укладки αβ-білків Структурна організація білків
- 38. “Мистецьке” походження терміну “мотиви” згортання молекул білків Структурна організація білків
- 39. Нейрамінідаза Структурна організація білків
- 40. Ацетілтрансфераза Структурна організація білків
- 41. Ретінол-зв’язуючий білок Структурна організація білків
- 42. Утворення суперспіралі у α-спіральних білках Взаємодія спіралей колагену Структурна організація білків
- 43. Третинна структура Формується за рахунок далекодіючих взаємодій між амінокислотними залишками в різних ділянках поліпептидного ланцюга. Поліпептид
- 44. Спрощені моделі молекул білків Третинна і четвертинна структура білків Структурна організація білків
- 45. Динаміка білкової структури Структурна організація білків
- 46. Мимовільне згортання поліпептидного ланцюга Структурна організація білків
- 47. Ієрархія структурної організації білка та послідовність його згортання Структурна організація білків
- 48. Ферменти – як представники групи білків Ферменти (ензими) — органічні каталізатори білкової природи, які утворюються в
- 49. Механізм дії ферментів Найбільші рівні структури білків третинна та четвертинна структури — руйнуються при нагріванні або
- 50. Механізм дії ферментів Специфічність Ферменти зазвичай проявляють високу специфічність по відношенню до своїх субстратів. Це досягається
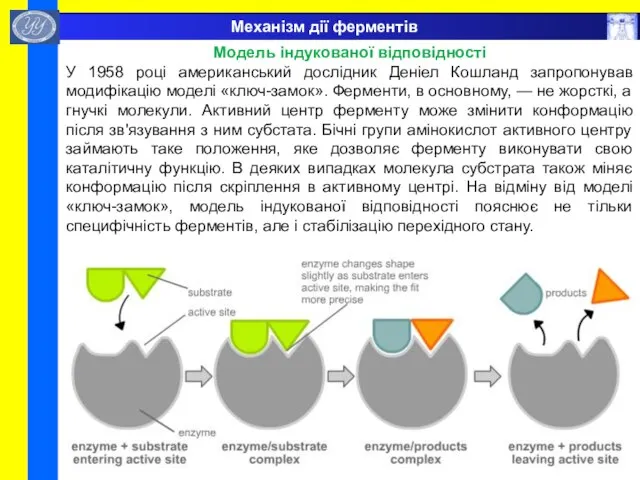
- 51. Механізм дії ферментів Модель індукованої відповідності У 1958 році американський дослідник Деніел Кошланд запропонував модифікацію моделі
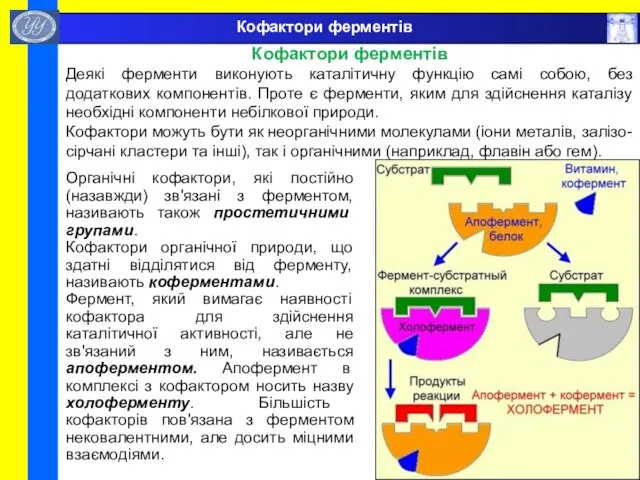
- 52. Кофактори ферментів Кофактори ферментів Деякі ферменти виконують каталітичну функцію самі собою, без додаткових компонентів. Проте є
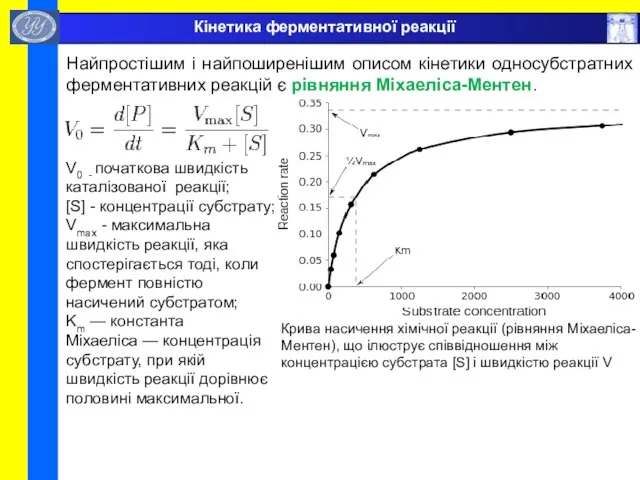
- 53. Кінетика ферментативної реакції Найпростішим і найпоширенішим описом кінетики односубстратних ферментативних реакцій є рівняння Міхаеліса-Ментен. Крива насичення
- 55. Скачать презентацию
Слайд 2Біофізика білків
Білки – молекулярні “машини”.
Структурна організація білків.
Ферменти – як представники групи білків.
Механізм
Біофізика білків
Білки – молекулярні “машини”.
Структурна організація білків.
Ферменти – як представники групи білків.
Механізм

Слайд 3Білки – молекулярні “машини”
Білки – молекулярні “машини”

Слайд 4Молекула білку – це молекулярна конструкція
Первинна
Вторинна
Третинна
Четвертинна
Білки – молекулярні “машини”
Молекула білку – це молекулярна конструкція
Первинна
Вторинна
Третинна
Четвертинна
Білки – молекулярні “машини”
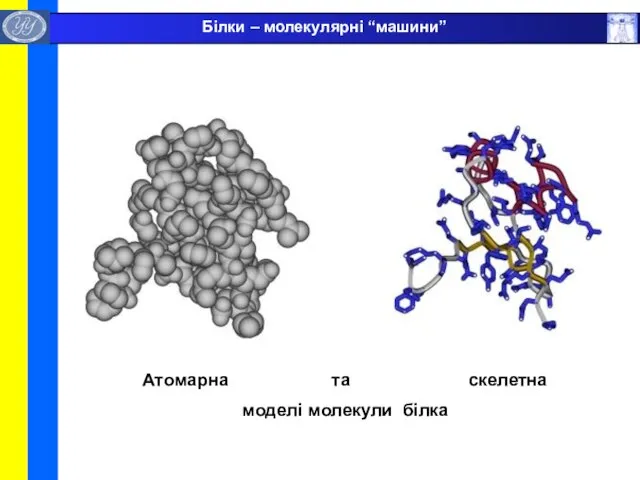
Слайд 5Атомарна та скелетна
моделі молекули білка
Білки – молекулярні “машини”
Атомарна та скелетна
моделі молекули білка
Білки – молекулярні “машини”
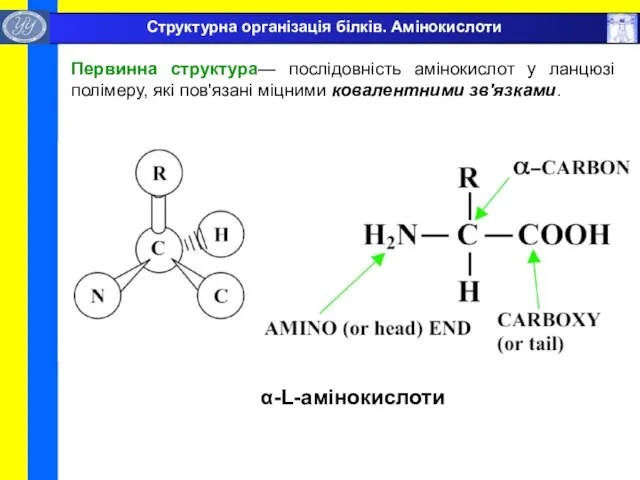
Слайд 6α-L-амінокислоти
Структурна організація білків. Амінокислоти
Первинна структура— послідовність амінокислот у ланцюзі полімеру, які пов'язані
α-L-амінокислоти
Структурна організація білків. Амінокислоти
Первинна структура— послідовність амінокислот у ланцюзі полімеру, які пов'язані
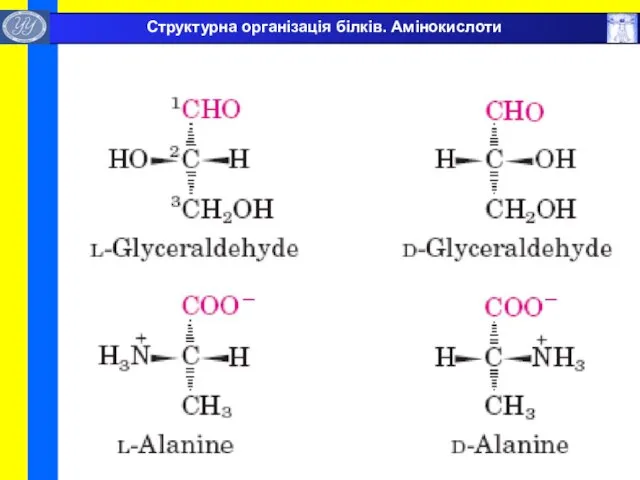
Слайд 7Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти
Слайд 8Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти

Слайд 9Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти
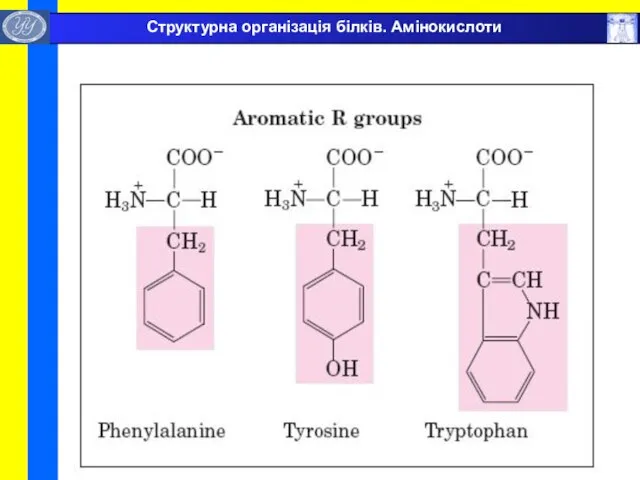
Слайд 10Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти
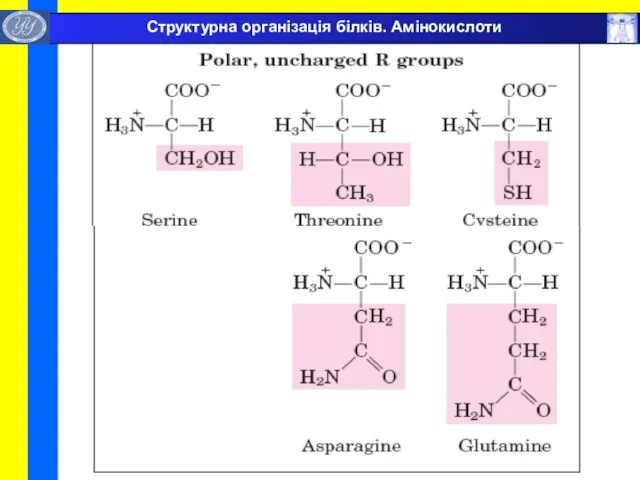
Слайд 11Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти
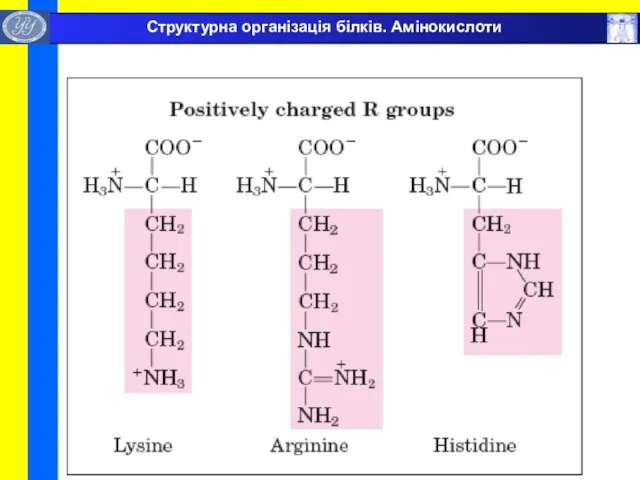
Слайд 12Структурна організація білків. Амінокислоти
Структурна організація білків. Амінокислоти

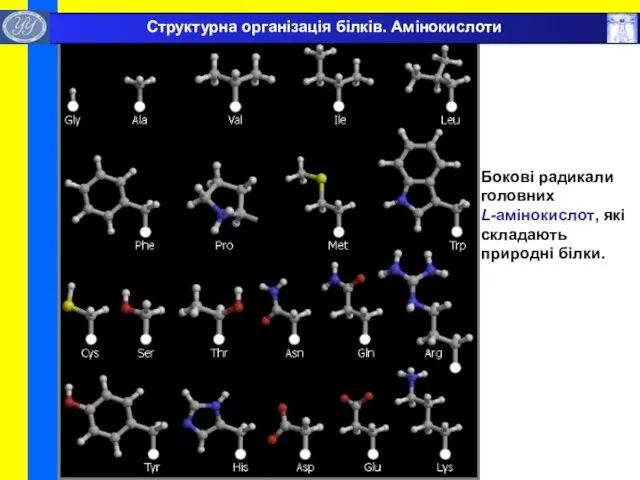
Слайд 13Бокові радикали головних
L-амінокислот, які складають природні білки.
Структурна організація білків. Амінокислоти
Бокові радикали головних
L-амінокислот, які складають природні білки.
Структурна організація білків. Амінокислоти
Слайд 14Білки – молекулярні конструкції
Амінокислоти
Тирозин
Білки – молекулярні конструкції
Амінокислоти
Тирозин
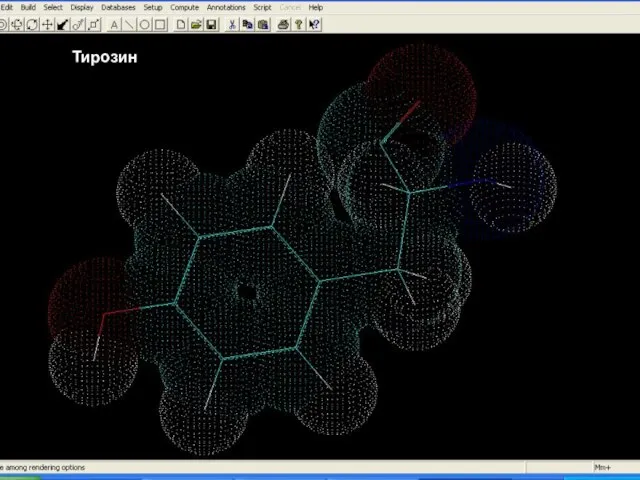
Слайд 15Білки – молекулярні конструкції
Амінокислоти
Тирозин
Білки – молекулярні конструкції
Амінокислоти
Тирозин
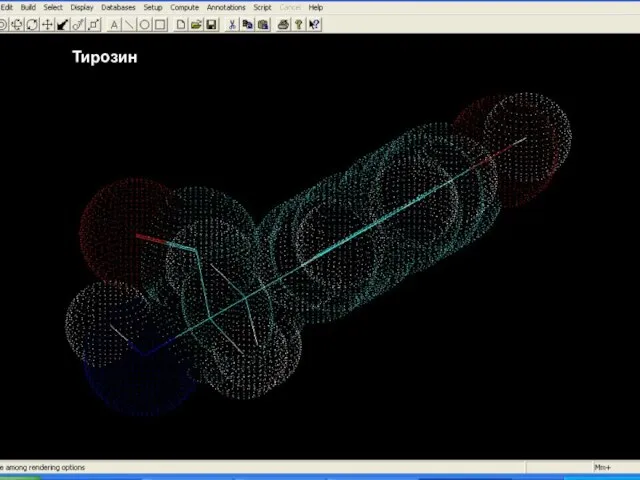
Слайд 16Білки – молекулярні конструкції
Амінокислоти
Фенілаланін
Білки – молекулярні конструкції
Амінокислоти
Фенілаланін
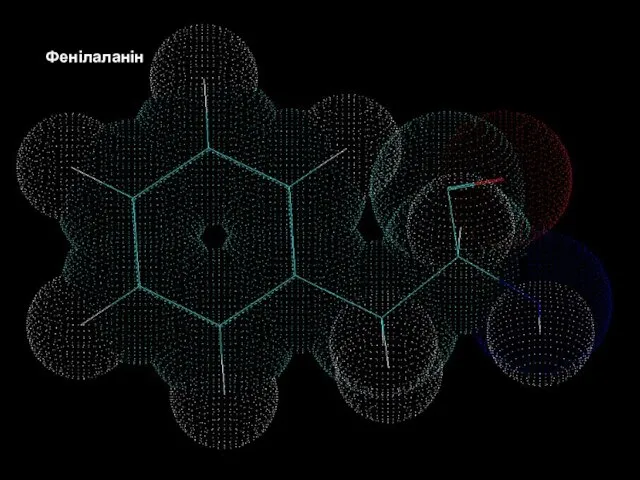
Слайд 17Білки – молекулярні конструкції
Амінокислоти
Гістидин
Білки – молекулярні конструкції
Амінокислоти
Гістидин
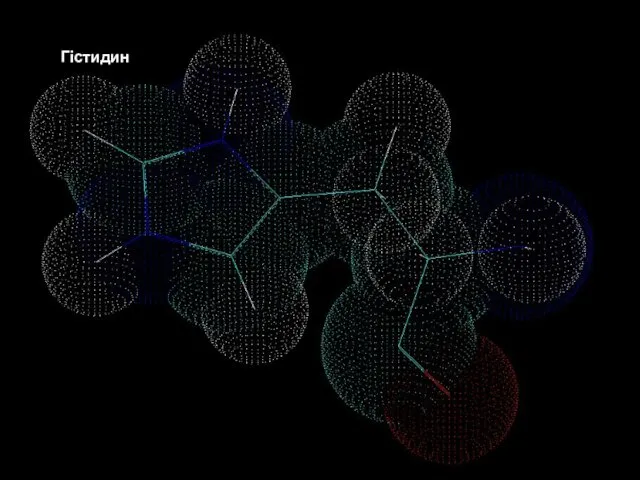
Слайд 18Білки – молекулярні конструкції
Амінокислоти
Валін
Білки – молекулярні конструкції
Амінокислоти
Валін

Слайд 19Білки – молекулярні конструкції
Амінокислоти
Аспарагінова кислота
Білки – молекулярні конструкції
Амінокислоти
Аспарагінова кислота

Слайд 20Білки – молекулярні конструкції
Амінокислоти
Глутамін
Білки – молекулярні конструкції
Амінокислоти
Глутамін

Слайд 21Методи дослідження амінокислот
Хроматографічні.
Електрофоретичні.
Оптичні (поглинання і флуоресценція).
Маспектрометричні.
Структурна організація білків. Амінокислоти
Методи дослідження амінокислот
Хроматографічні.
Електрофоретичні.
Оптичні (поглинання і флуоресценція).
Маспектрометричні.
Структурна організація білків. Амінокислоти

Слайд 22 Зв'язки, які забезпечують стабілізацію просторової структури біомакромолекул:
ковалентні зв'язки;
електростатичні взаємодії;
водневі зв'язки;
Ван-дер-Ваальсові
Зв'язки, які забезпечують стабілізацію просторової структури біомакромолекул:
ковалентні зв'язки;
електростатичні взаємодії;
водневі зв'язки;
Ван-дер-Ваальсові

Слайд 23Рівні структурної організації білків
Первинна структура (1º) - амінокислотна послідовність білків;
Вторинна структура (2º)
Рівні структурної організації білків
Первинна структура (1º) - амінокислотна послідовність білків;
Вторинна структура (2º)

Слайд 24Взаємодії в пептидному зв'язку
Структурна організація білків
Взаємодії в пептидному зв'язку
Структурна організація білків
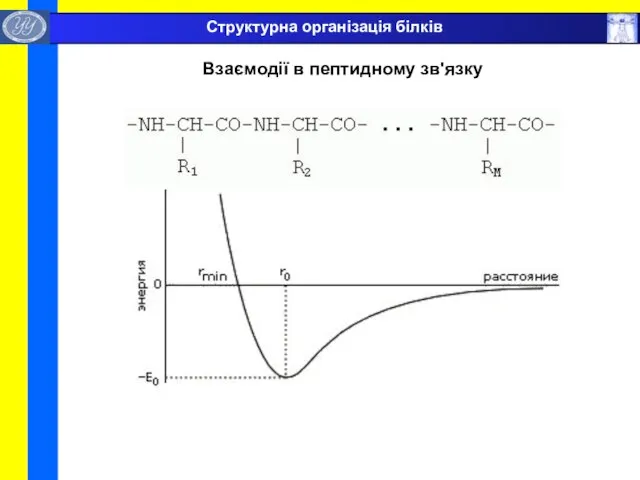
Слайд 25“Конструкція” пептидного зв'язку
Обертання навколо одинарного зв'язку
Обертання навколо пептидного зв'язку
практично неможливо
Структурна організація
“Конструкція” пептидного зв'язку
Обертання навколо одинарного зв'язку
Обертання навколо пептидного зв'язку
практично неможливо
Структурна організація
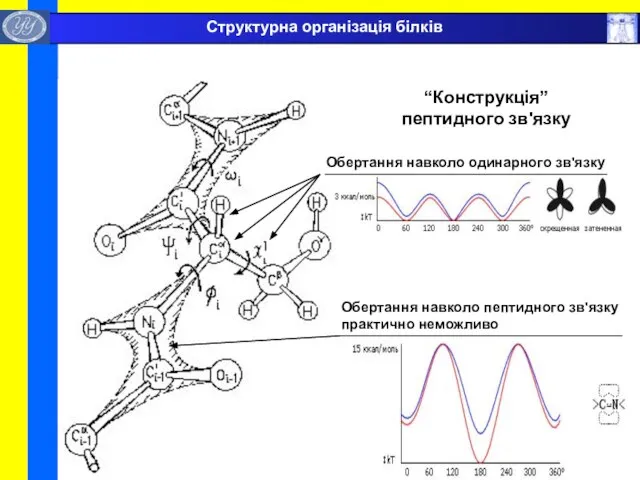
Слайд 26Білки – молекулярні конструкції
Білки – молекулярні конструкції
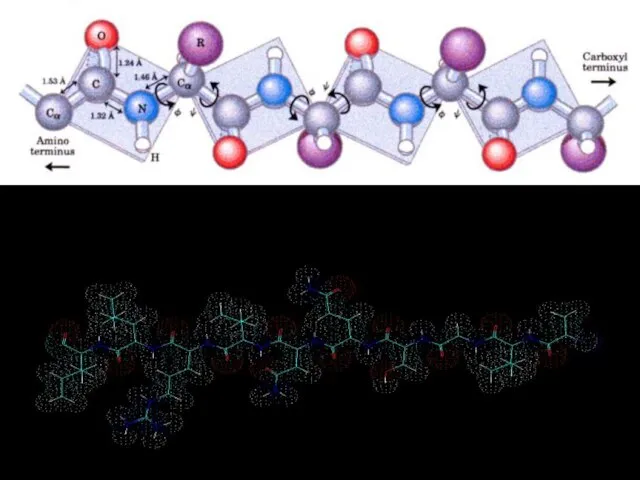
Слайд 27Вторинна структура характеризує просторову форму білкової молекули, яка найчастіше повністю або частково
Вторинна структура характеризує просторову форму білкової молекули, яка найчастіше повністю або частково

Слайд 28α-спіралі — щільні витки навколо довгої осі структури, один виток становлять 4
α-спіралі — щільні витки навколо довгої осі структури, один виток становлять 4
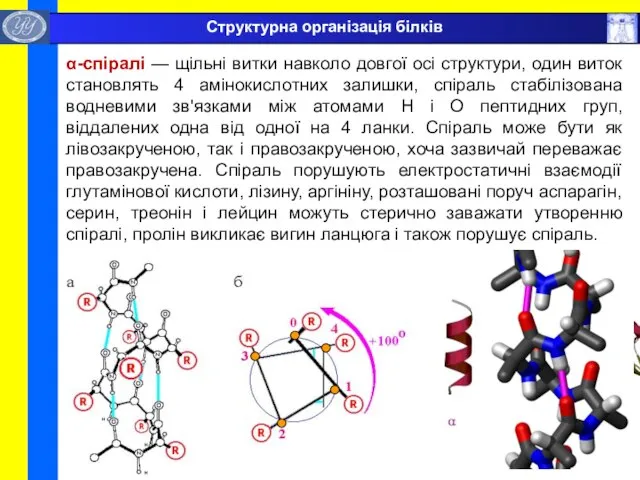
Слайд 29β-листи (складчасті шари) — декілька зигзагоподібних поліпептідних ланцюжків, в яких водневі зв'язки
β-листи (складчасті шари) — декілька зигзагоподібних поліпептідних ланцюжків, в яких водневі зв'язки

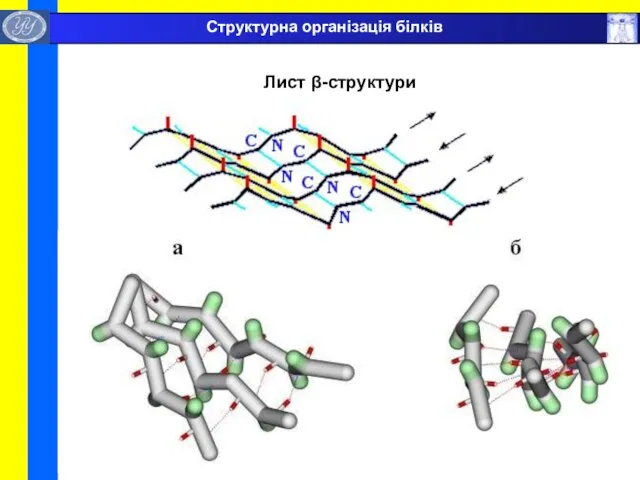
Слайд 30Лист β-структури
Структурна організація білків
Лист β-структури
Структурна організація білків
Слайд 31Приклади розташування β-тяжів
Приклади розташування β-тяжів

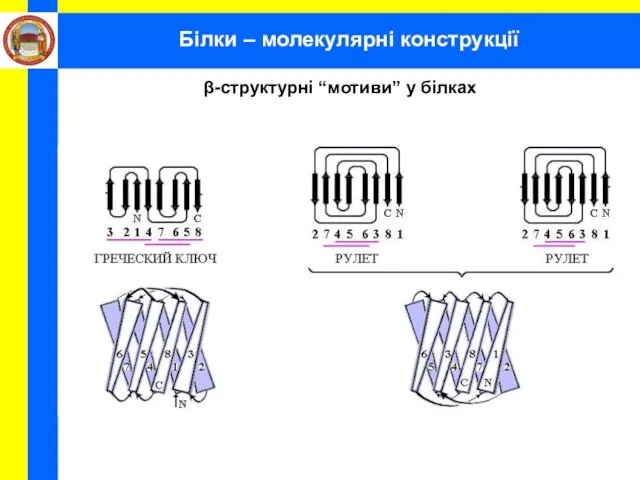
Слайд 32Білки – молекулярні конструкції
β-структурні “мотиви” у білках
Білки – молекулярні конструкції
β-структурні “мотиви” у білках
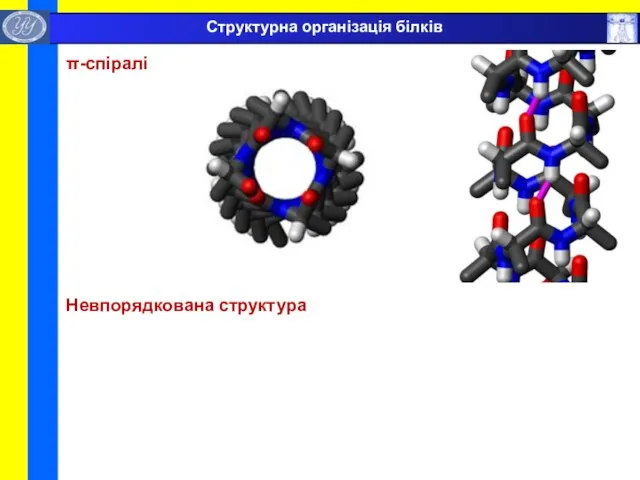
Слайд 33π-спіралі
Невпорядкована структура
Структурна організація білків
π-спіралі
Невпорядкована структура
Структурна організація білків
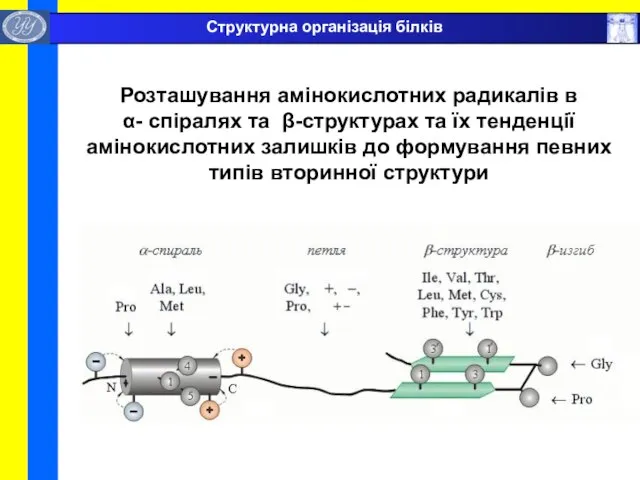
Слайд 34Розташування амінокислотних радикалів в
α- спіралях та β-структурах та їх тенденції амінокислотних
Розташування амінокислотних радикалів в α- спіралях та β-структурах та їх тенденції амінокислотних
Слайд 35«Надвторинна» структура
Більшість білків організовані на кількох проміжних рівнях між основними елементами вторинної
«Надвторинна» структура
Більшість білків організовані на кількох проміжних рівнях між основними елементами вторинної

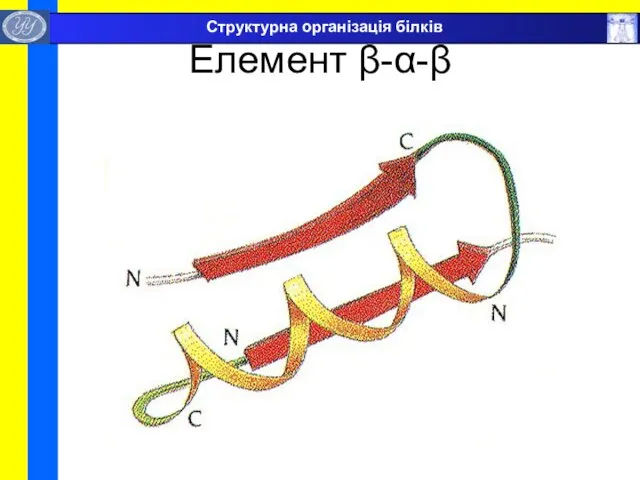
Слайд 36Елемент β-α-β
Структурна організація білків
Елемент β-α-β
Структурна організація білків
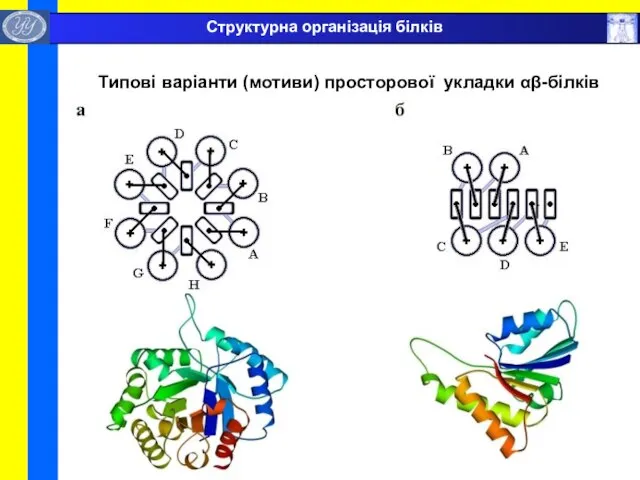
Слайд 37Типові варіанти (мотиви) просторової укладки αβ-білків
Структурна організація білків
Типові варіанти (мотиви) просторової укладки αβ-білків
Структурна організація білків
Слайд 38“Мистецьке” походження терміну
“мотиви” згортання молекул білків
Структурна організація білків
“Мистецьке” походження терміну
“мотиви” згортання молекул білків
Структурна організація білків
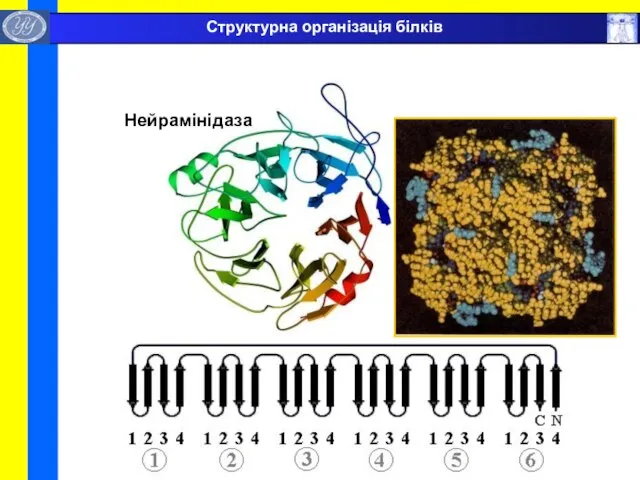
Слайд 39Нейрамінідаза
Структурна організація білків
Нейрамінідаза
Структурна організація білків
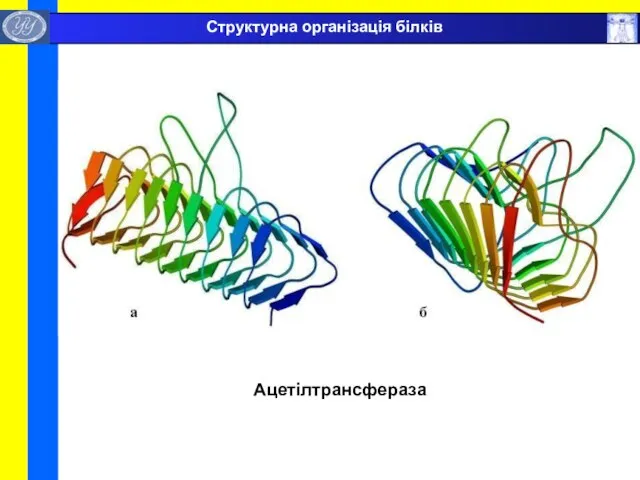
Слайд 40Ацетілтрансфераза
Структурна організація білків
Ацетілтрансфераза
Структурна організація білків
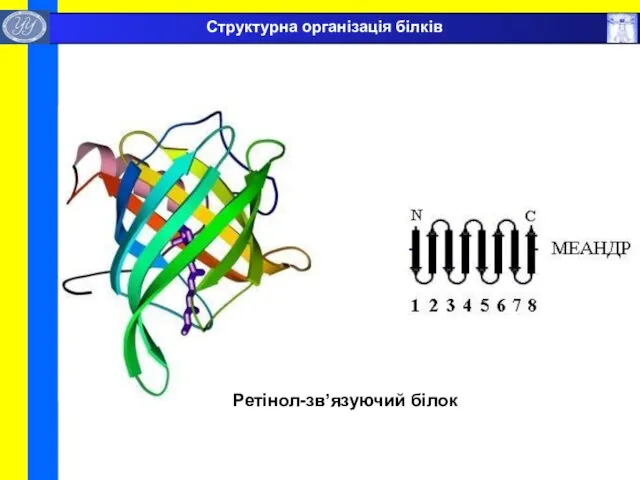
Слайд 41Ретінол-зв’язуючий білок
Структурна організація білків
Ретінол-зв’язуючий білок
Структурна організація білків
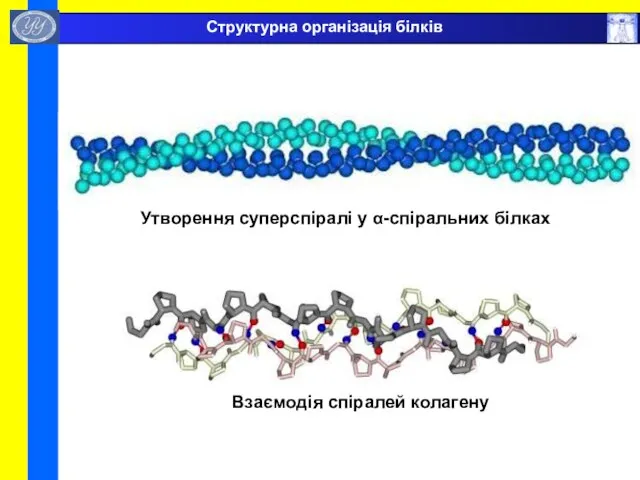
Слайд 42Утворення суперспіралі у α-спіральних білках
Взаємодія спіралей колагену
Структурна організація білків
Утворення суперспіралі у α-спіральних білках
Взаємодія спіралей колагену
Структурна організація білків
Слайд 43Третинна структура
Формується за рахунок далекодіючих взаємодій між амінокислотними залишками в різних ділянках
Третинна структура
Формується за рахунок далекодіючих взаємодій між амінокислотними залишками в різних ділянках

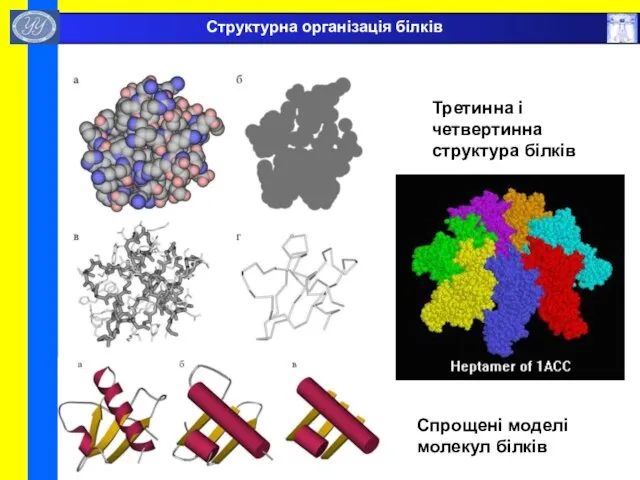
Слайд 44Спрощені моделі молекул білків
Третинна і четвертинна
структура білків
Структурна організація білків
Спрощені моделі молекул білків
Третинна і четвертинна
структура білків
Структурна організація білків
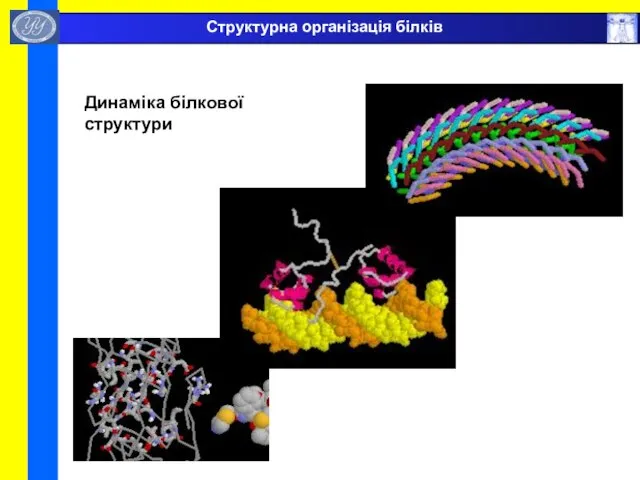
Слайд 45Динаміка білкової структури
Структурна організація білків
Динаміка білкової структури
Структурна організація білків
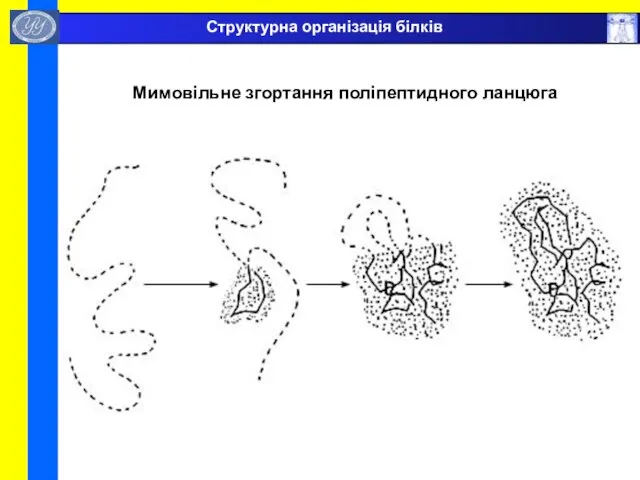
Слайд 46Мимовільне згортання поліпептидного ланцюга
Структурна організація білків
Мимовільне згортання поліпептидного ланцюга
Структурна організація білків
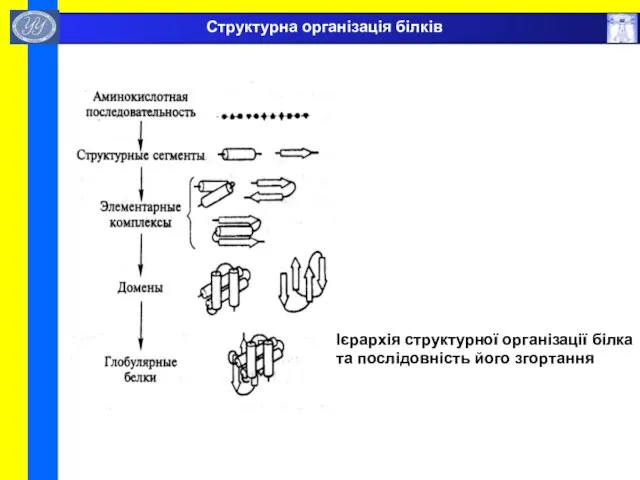
Слайд 47Ієрархія структурної організації білка
та послідовність його згортання
Структурна організація білків
Ієрархія структурної організації білка
та послідовність його згортання
Структурна організація білків
Слайд 48Ферменти – як представники групи білків
Ферменти (ензими) — органічні каталізатори білкової природи, які утворюються в живих організмах,
Ферменти – як представники групи білків
Ферменти (ензими) — органічні каталізатори білкової природи, які утворюються в живих організмах,

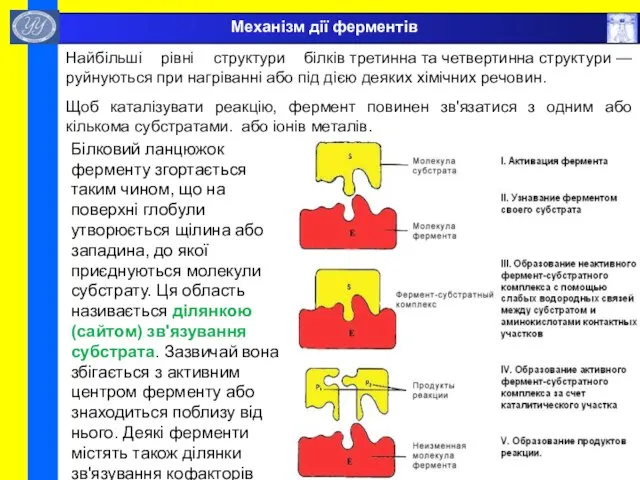
Слайд 49Механізм дії ферментів
Найбільші рівні структури білків третинна та четвертинна структури — руйнуються при нагріванні або під дією
Механізм дії ферментів
Найбільші рівні структури білків третинна та четвертинна структури — руйнуються при нагріванні або під дією
Слайд 50Механізм дії ферментів
Специфічність
Ферменти зазвичай проявляють високу специфічність по відношенню до своїх субстратів.
Механізм дії ферментів
Специфічність
Ферменти зазвичай проявляють високу специфічність по відношенню до своїх субстратів.

Слайд 51Механізм дії ферментів
Модель індукованої відповідності
У 1958 році американський дослідник Деніел Кошланд запропонував
Механізм дії ферментів
Модель індукованої відповідності
У 1958 році американський дослідник Деніел Кошланд запропонував
Слайд 52Кофактори ферментів
Кофактори ферментів
Деякі ферменти виконують каталітичну функцію самі собою, без додаткових компонентів.
Кофактори ферментів
Кофактори ферментів
Деякі ферменти виконують каталітичну функцію самі собою, без додаткових компонентів.
Слайд 53Кінетика ферментативної реакції
Найпростішим і найпоширенішим описом кінетики односубстратних ферментативних реакцій є рівняння Міхаеліса-Ментен.
Крива
Кінетика ферментативної реакції
Найпростішим і найпоширенішим описом кінетики односубстратних ферментативних реакцій є рівняння Міхаеліса-Ментен.
Крива
 Тандырная это – приготовление шашлыков в тандыре
Тандырная это – приготовление шашлыков в тандыре Лист. Внешнее и внутреннее строение
Лист. Внешнее и внутреннее строение Архитектура и дизайн
Архитектура и дизайн Международное гуманитарное право.
Международное гуманитарное право. Метод фундаментального проектирования Мэтчетта
Метод фундаментального проектирования Мэтчетта Контроль, согласование и утверждение КТД
Контроль, согласование и утверждение КТД Москва (2 класс)
Москва (2 класс) Подготовка дела к судебному разбирательству как обязательная стадия гражданского судопроизводства
Подготовка дела к судебному разбирательству как обязательная стадия гражданского судопроизводства Презентация на тему Вспоминаем Юрия Гагарина 55 лет со дня первого полета человека в Космос
Презентация на тему Вспоминаем Юрия Гагарина 55 лет со дня первого полета человека в Космос  История учения о личности преступника. Тема 2
История учения о личности преступника. Тема 2 Sonder для рассылки
Sonder для рассылки Сертификация
Сертификация Бизнес-партнерство с Rama Yoga
Бизнес-партнерство с Rama Yoga Презентация на тему Движущие силы антропогенеза
Презентация на тему Движущие силы антропогенеза  Геометрия вокруг нас
Геометрия вокруг нас БЮДЖЕТИРОВАНИЕ В 1С
БЮДЖЕТИРОВАНИЕ В 1С Мошенничества с инвестициями
Мошенничества с инвестициями Природное и общественное. Отличия человека от животного
Природное и общественное. Отличия человека от животного Жарқын болашаққа жол
Жарқын болашаққа жол Раскадровка. Для чего нужна раскадровка
Раскадровка. Для чего нужна раскадровка Культурное наследие регионов России
Культурное наследие регионов России Презентация на тему Дисграфия
Презентация на тему Дисграфия Профессиональные функции и умения старшего воспитателя ДОУ
Профессиональные функции и умения старшего воспитателя ДОУ Иллюзии зрения
Иллюзии зрения Презентация на тему Развитие метапредметных компетенций учащихся
Презентация на тему Развитие метапредметных компетенций учащихся Образец презентации к отчету ИР БкЮ-100
Образец презентации к отчету ИР БкЮ-100 Презентация на тему Роль права в жизни государства (9 класс)
Презентация на тему Роль права в жизни государства (9 класс) Классификация элементарных частиц
Классификация элементарных частиц