- Презентация на тему Физиология микроорганизмов

Содержание
- 2. Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий реакции, связанные с получением
- 3. При использовании в качестве источника углерода и энергии глюкозы или других гексоз начальные этапы окисления глюкозы
- 4. Пути расщепления глюкозы. Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных продуктов обмена веществ, у
- 5. Пути расщепления глюкозы 1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или, по имени изучавших его
- 6. Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента гексокиназы до метаболически активной формы
- 7. ФДФ-путь. Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во фруктозо-1,6-дифосфат, который в дальнейшем через
- 8. ПФ-путь. В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается в рибулезо-5-фосфат (Ри-5-Ф), который находится
- 9. Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат, замыкая реакции в цикл, и
- 10. КДФГ-путь (путь Этнера—Дудорова) Этот путь расщепления глюкозы специфичен только для бактерий. Встречается у бактерий, потерявших фермент
- 11. Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под действием дегидрогеназы отщепляется вода и
- 12. На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного НАД и 1 молекула восстановленного
- 13. Окислительный метаболизм у бактерий (дыхание)
- 14. Окислительный метаболизм Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания. Дыхание— процесс получения энергии в реакциях
- 15. В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные анаэробы и облигатные анаэробы.
- 16. Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются только в присутствии кислорода.
- 17. Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии. Тип метаболизма у них бродильный.
- 18. Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться как в присутствии кислорода, так
- 19. В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода Н2О2 и закисный радикал
- 20. У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является молекулярный кислород. В этом случае
- 21. Цикл трикарбоновых кислот (цикл Кребса)
- 22. Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов, так и атомов водорода, который
- 23. Типичная цепь выглядит следующим образом: ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
- 24. Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным этапом переноса электронов (протонов) по
- 25. Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков в ЦПМ приводит к тому,
- 26. У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит непосредственный перенос водорода на кислород
- 27. Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки, в качестве источника энергии, окисляя
- 29. Скачать презентацию
Слайд 2Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий
Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий

Слайд 3При использовании в качестве источника углерода и энергии глюкозы или других гексоз
При использовании в качестве источника углерода и энергии глюкозы или других гексоз

Слайд 4Пути расщепления глюкозы.
Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных
Пути расщепления глюкозы.
Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных
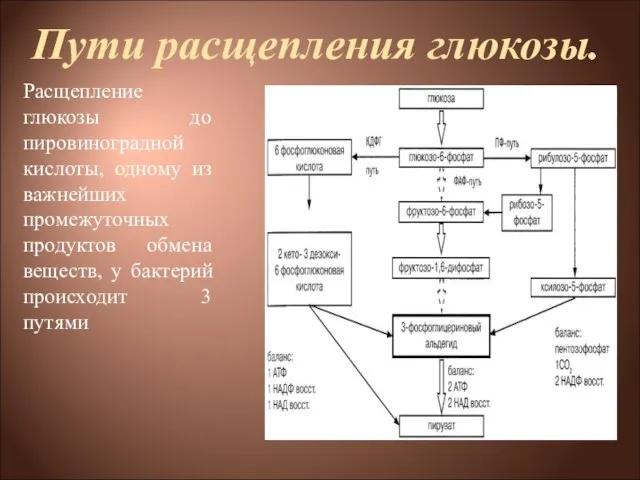
Слайд 5Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или, по
Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или, по

Слайд 6Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента гексокиназы
Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента гексокиназы

Слайд 7ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во фруктозо-1,6-дифосфат,
ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во фруктозо-1,6-дифосфат,

Слайд 8ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается в
ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается в

Слайд 9Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат, замыкая
Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат, замыкая

Слайд 10КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий. Встречается
КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий. Встречается

Слайд 11Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под действием
Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под действием

Слайд 12На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного НАД
На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного НАД

Слайд 13Окислительный метаболизм у бактерий (дыхание)
Окислительный метаболизм у бактерий (дыхание)
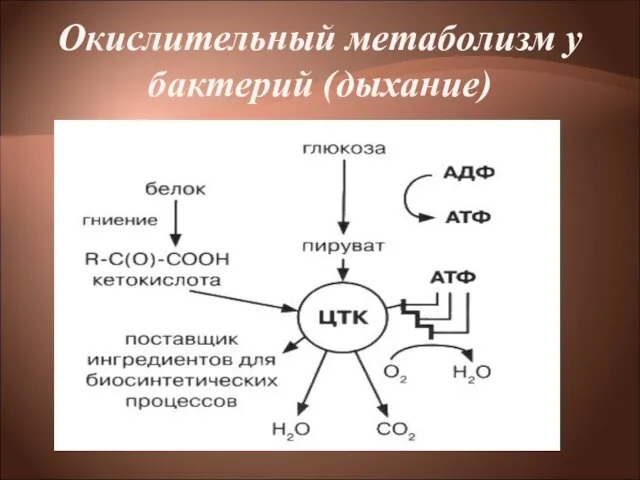
Слайд 14Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание— процесс получения энергии
Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание— процесс получения энергии

Слайд 15В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные
В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные

Слайд 16Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются
Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются

Слайд 17Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии. Тип
Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии. Тип

Слайд 18Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться как
Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться как

Слайд 19В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода
В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода

Слайд 20У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является молекулярный
У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является молекулярный

Слайд 21Цикл трикарбоновых кислот (цикл Кребса)
Цикл трикарбоновых кислот (цикл Кребса)
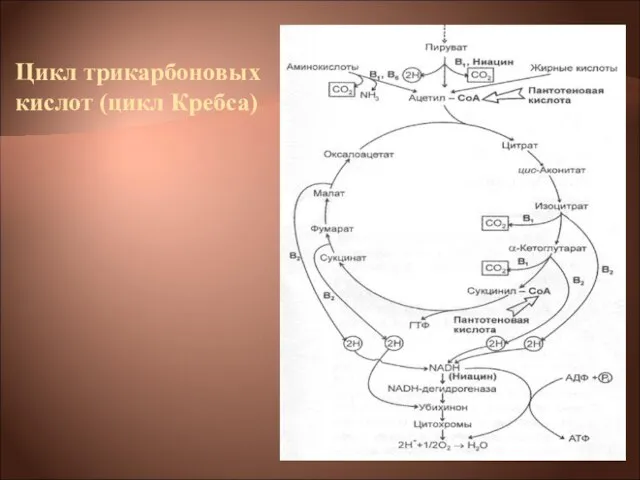
Слайд 22Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов, так
Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов, так

Слайд 23Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2

Слайд 24Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным этапом
Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным этапом

Слайд 25Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков в
Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков в

Слайд 26У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит непосредственный
У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит непосредственный

Слайд 27Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки, в
Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки, в

 Урок презентация на тему Мировое хозяйство (4 класс)
Урок презентация на тему Мировое хозяйство (4 класс) Источники информации о населении и демографических процессах
Источники информации о населении и демографических процессах АЛЕРГІЯ
АЛЕРГІЯ Народное творчество – плетение лаптей
Народное творчество – плетение лаптей История национального костюма
История национального костюма Урок - игра Мальвина и цветик-семицветик. 2 класс
Урок - игра Мальвина и цветик-семицветик. 2 класс Проект Тыводитель
Проект Тыводитель Организация методической работыв МОУДОД«Центр внешкольной работы»
Организация методической работыв МОУДОД«Центр внешкольной работы» Динамическое программирование
Динамическое программирование Техника дыхания
Техника дыхания Налог на доходы физических лиц
Налог на доходы физических лиц Банковская карта
Банковская карта Орловский народный костюм
Орловский народный костюм Электрофизические свойства водородосодержащих доноров в субмикронных слоях кремния Магистерская диссертация Выполнил – м
Электрофизические свойства водородосодержащих доноров в субмикронных слоях кремния Магистерская диссертация Выполнил – м Первичные документы, составляемые по итогам инвентаризации отдельных видов имущества
Первичные документы, составляемые по итогам инвентаризации отдельных видов имущества Мёртвые души
Мёртвые души Марко Вовчок. Життєвий та творчишлях
Марко Вовчок. Життєвий та творчишлях ОБЩЕСТВЕННЫЙ ПРОЕКТ: РОЖДЕНИЕ, ПРОДВИЖЕНИЕ, ЭФФЕКТИВНАЯ РЕАЛИЗАЦИЯ
ОБЩЕСТВЕННЫЙ ПРОЕКТ: РОЖДЕНИЕ, ПРОДВИЖЕНИЕ, ЭФФЕКТИВНАЯ РЕАЛИЗАЦИЯ Learner Profile
Learner Profile Медведева Ольга Евгеньевна, д.э.н., профессор Государственного университета управления
Медведева Ольга Евгеньевна, д.э.н., профессор Государственного университета управления  Буква закона
Буква закона Чеканка (тиснение по фольге)
Чеканка (тиснение по фольге) Музыка моего народа
Музыка моего народа «Развитие транспортной инфраструктуры флота река-море РФ. (2012-2015 гг.)»
«Развитие транспортной инфраструктуры флота река-море РФ. (2012-2015 гг.)» Работа с гипотезами
Работа с гипотезами Vistula University
Vistula University Модернизированная автоматизированная информационная система управления государственными и муниципальными закупками Республик
Модернизированная автоматизированная информационная система управления государственными и муниципальными закупками Республик Презентация на тему Координация и регуляция
Презентация на тему Координация и регуляция