- Развиће

Содержание
- 2. Онтогенетско (индивидуално) развиће (грч. онтос = биће; генесис = постанак, развиће) процеси прображаја оплођеног јајета или
- 3. Полно и бесполно размножавање
- 4. Основне фазе онтогенетског развића животиња Гаметогенеза Оплођење (фертилизација) Браздање Гаструлација Органогенеза Раст и хистолошка диференцијација Метаморфоза
- 5. Гаметогенеза Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета: јајних ћелија - оогенеза (овогенеза)
- 6. Оплођење (фертилизација) Оплођење које представља спајање женског и мушког гамета. Спајањем њихових једара настаје диплоидно једро
- 7. Браздање Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско тело, најчешће лоптастог облика
- 8. Гаструлација У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим и трослојно тело -
- 9. Органогенеза У фази органогенезе, петој фази развића, долази до формирања органа. У сваком од клициних листова
- 10. Раст и хистолошка диференцијација Развиће различитих типова ћелија Образовани зачеци органа расту, па тако животиња ступњевито
- 11. Метаморфоза и регенерација Последња, седма фаза обухвата све процесе који се јављају у каснијем животу јединке,
- 12. Диференцијација У процесу развића ћелије се диференцирају у различита ткива и органе При томе добијају различите
- 14. Диференцијација – на нивоу гена У основи диференцијације стоји смењивање активности различитих група гена које делују
- 15. Гаметогенеза Основни генетски потенцијал новог организма тј. нове генерације потиче из гамета које стварају родитељи Гамети
- 16. Мејоза У полним органима животиња и биљака настају полне ћелије – гамети. Гамети имају половину укупног
- 17. Профаза I
- 18. Лептонема (лептотен) постепена кондензација хромозома
- 19. Зигонема (зиготен) међусобно приближавање хомологих хромозома. Формирање хромозомских парова – БИВАЛЕНАТА. Хромозоми се пиљубљују у процесу
- 20. Пахинема (пахитен) оба приљубљена хромозома се деле уздужно на по две хроматиде везане заједничком центромером. Тако
- 21. Диплонема (диплотен) Хроматиде хомологих хромозома се међусобно раздвајају (одмичу) Остају везане преко хијазми које су у
- 22. Дијакинеза (дијакинезис) Спирализација хромозома и терминализација хијазми достиже највећи степен Тетраде добијају циклични облик Једарце нестаје
- 23. Метафаза I Тетраде се крећу према екваторијалној равни и постављају у њеној оси Свака тетрада има
- 24. Анфаза I Раздвајање хомологих хромозома, кидање терминалних хијазми Хомологи хромозоми полазе ка супротним половима (дијаде, половине
- 25. Број хромозома се тако своди на половину, а како се биваленти раздвајају независно, мајчински и очински
- 26. Телофаза I Два хаплоидна сета хромозома се групишу на супротним половима ћелије Појављује се једарце Ствара
- 27. Друга мејотичка деоба – Мејоза II Слична је митози Број хромозома који улази у митозу II
- 28. Гамети воде порекло од примарних герминативних ћелија које се јављају у ембриону човека крајем треће недеље.
- 29. Оогенеза Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском деобом примарних герминативних ћелија при
- 31. Оогенеза У време рођења, у оба јајника се налази око 2 милиона примарних фоликула, а затим
- 32. Оогенеза - мејоза Примарне герминативне ћелије оваријума митозама се деле и дају ООГОНИЈЕ Оогоније расту једно
- 33. Оогенеза - мејоза I ДЕОБА: ооцита I реда дели се на две неједнаке ћелије: ооциту II
- 34. Оогенеза - мејоза
- 35. Вителогенеза Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ Општи назив за резервни материјал јајета
- 36. Типови јајних ћелија Зависно од количине жуманцета и његове распрострањености у јајној ћелији могу се разликовати
- 37. Сперматогенеза Сперматогенеза је процес настанка сперматозоида. Почиње у пубертету Она се континуирано одвија у семеним каналићима
- 38. Улога сперматозоида Активација јајне ћелије да отпочне са развићем Да своју хаплоидну гарнитуру хромозома преда јајној
- 39. Сперматогенеза се одвија у 4 фазе: СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом и настају примарни сперматоцити,
- 41. Сперматозоиди Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm. Акрозом – прекрива предњи део
- 42. Сертолијеве ћелије Исхрана исходних клициних ћелија Ослобађање сперматозоида у лумен семених каналића
- 44. ОПЛОЂЕЊЕ - Фертилизација - Три главне функције фертилизације су: Трансмисија гена са оба родитеља на потомке
- 45. TIPOVI OPLOĐENJA I RAZVIĆA U životinjskom svetu se , prema mestu odvijanja, razlikuju dva osnovna tipa
- 46. Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće embriona vrše u spoljašnjoj sredini
- 47. Фертилизација пролази кроз следеће фазе: Контакт између сперматозоида и јајне ћелије (акрозомална реакција) Улазак сперматозоида у
- 48. Aktivacija spermatozoida počinje kada on dodirne omotač jajne ćelije što dovodi do tzv. akrozomske reakcije. Akrozomska
- 49. ФУЗИЈА НУКЛЕУСА Након продора сперматозоида у јајну ћелију, долази до значајних промена у јајној ћелији Прво
- 50. Ране деобе (браздање)
- 51. Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу је, у односу на јајну
- 52. Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома брзо ћелијском деобом. Ћелијске деобе
- 53. Типови браздања Радијално браздање - на ступњу од 8 бластомера свака бластомера горњег (анималног) слоја лежи
- 54. Жуманце има велики утицај на браздање. Вителус смањује браздање на вегетативном полу ембриона. Браздање увек отпочиње
- 55. Бластулација Бластула настаје митотичким деобама зигота Има лоптаст облик Један слој ћелија БЛАСТОМЕРА формира БЛАСТОДЕРМ Унутрашњост
- 56. БЛАСТУЛА
- 57. Типови бластула Постоје четири типа бластула: целобластула – то је лопта на чијој поврчини се налази
- 58. Сисари Од изолецитног оплођеног јајета образује се морула, а потом бластодиск у коме се образује дупља
- 59. Гаструлација Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где ће оне формирати три примарна
- 60. Начини формирања гаструле су: инвагинација – гаструла настаје из целобластуле, која се на једном делу угиба
- 61. Гаструлација Три примарна ћелијска слоја су: ЕКТОДЕРМ – спољашњи слој МЕЗОДЕРМ – средишњи слој ЕНДОДЕРМ -
- 62. Морфогенеза – нижи хордати Морфогенеза је прогресивно стицање телесног облика и развој одређених структура током ембр.
- 63. Развиће амфиоксуса Браздањем настаје бластула Гаструлација почиње на ступњу од око 800 ћел Долази до заравњивања
- 64. Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев која има отвор неуропор Архентерон
- 65. Неурулација Крајем гаструлације, образује се НЕРВНА ПЛОЧА Нервна плоча Почиње неурулација, образовање НЕРВНЕ ЦЕВИ (6.)
- 66. Развиће водоземаца Крајем гаструлације код водоземаца се формира се нервна плоча – то је једна равна
- 67. Неурулација и ембрионална индукција На формирање организма битан утицај имају околна ткива, тј. средина у којој
- 68. Околна ткива индукују настанак одређеног ткива и органа. Тако настанак нервне цеви (неурулацију) могу да индукују
- 69. Органогенеза Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом Сва ткива и органи настају
- 70. Сматра се да су за то одговорни градијенти информационих молекула унутар ембриона који обавештавају ћелије о
- 71. Овде су дати кључни моменти процеса развића код животиња Најдетаљније су проучени код бодљокожацa, нижих хордата,
- 72. Развиће код различитих група
- 73. Ембриогенеза код човека Уместо бластоцела, ингресијом се развија БЛАСТОДИСК Као и код гмизаваца и птица, развијају
- 74. Екстраембрионалне опне Код гмизаваца и птица Жуманцетна кеса – садржи жуманце, има крвне судове који је
- 75. ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја ОВОВИВИПАРИТЕТ – доношење на свет живих младунаца
- 76. ПЛАЦЕНТАЦИЈА ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива ПО ГРАЂИ се деле на: ХОРИО-ВИТЕЛИНСКУ
- 77. Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива ПЛАЦЕНТАЛНА БАРИЈЕРА (слој ткива који омогућава транспорт)
- 78. Повећањем површине контакта Развитак наставака на површини хориона, тзв. ресица (villi) ДИФУЗНА ПЛАЦЕНТА – вили по
- 79. Смањење дебљине плаценталне баријере Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари и копитари) Ендотелио-хоријална плацента:
- 80. Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из ткива материце и не долази
- 82. Скачать презентацию
Слайд 2Онтогенетско (индивидуално) развиће
(грч. онтос = биће; генесис = постанак, развиће)
процеси прображаја
Онтогенетско (индивидуално) развиће
(грч. онтос = биће; генесис = постанак, развиће)
процеси прображаја

Слайд 3Полно и бесполно размножавање
Полно и бесполно размножавање

Слайд 4Основне фазе онтогенетског развића животиња
Гаметогенеза
Оплођење (фертилизација)
Браздање
Гаструлација
Органогенеза
Раст и хистолошка диференцијација
Метаморфоза и регенерација
Основне фазе онтогенетског развића животиња
Гаметогенеза
Оплођење (фертилизација)
Браздање
Гаструлација
Органогенеза
Раст и хистолошка диференцијација
Метаморфоза и регенерација

Слайд 5Гаметогенеза
Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета:
јајних ћелија
Гаметогенеза
Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета:
јајних ћелија

Слайд 6Оплођење (фертилизација)
Оплођење које представља спајање женског и мушког гамета.
Спајањем њихових једара
Оплођење (фертилизација)
Оплођење које представља спајање женског и мушког гамета.
Спајањем њихових једара

Слайд 7Браздање
Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско тело,
Браздање
Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско тело,

Слайд 8Гаструлација
У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим и
Гаструлација
У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим и

Слайд 9Органогенеза
У фази органогенезе, петој фази развића, долази до формирања органа.
У сваком
Органогенеза
У фази органогенезе, петој фази развића, долази до формирања органа.
У сваком

Слайд 10Раст и хистолошка диференцијација
Развиће различитих типова ћелија
Образовани зачеци органа расту, па тако
Раст и хистолошка диференцијација
Развиће различитих типова ћелија
Образовани зачеци органа расту, па тако

Слайд 11Метаморфоза и регенерација
Последња, седма фаза обухвата све процесе који се јављају
Метаморфоза и регенерација
Последња, седма фаза обухвата све процесе који се јављају

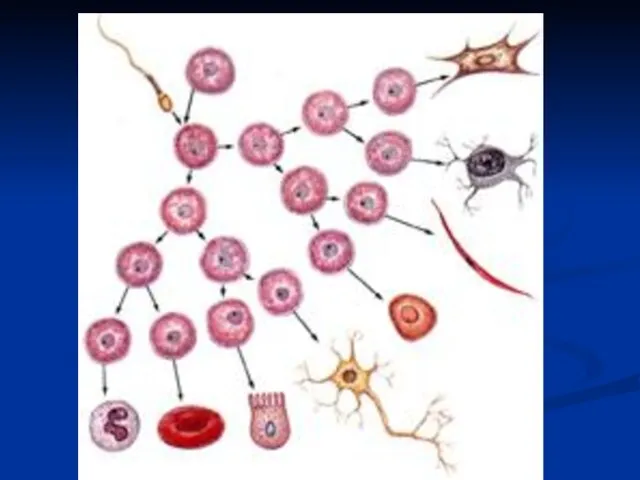
Слайд 12Диференцијација
У процесу развића ћелије се диференцирају у различита ткива и органе
При
Диференцијација
У процесу развића ћелије се диференцирају у различита ткива и органе
При

Слайд 14Диференцијација – на нивоу гена
У основи диференцијације стоји смењивање активности различитих група
Диференцијација – на нивоу гена
У основи диференцијације стоји смењивање активности различитих група

Слайд 15Гаметогенеза
Основни генетски потенцијал новог организма тј. нове генерације потиче из гамета
Гаметогенеза
Основни генетски потенцијал новог организма тј. нове генерације потиче из гамета

Слайд 16Мејоза
У полним органима животиња и биљака настају полне ћелије – гамети.
Мејоза
У полним органима животиња и биљака настају полне ћелије – гамети.

Слайд 17Профаза I
Профаза I

Слайд 18Лептонема (лептотен)
постепена кондензација хромозома
Лептонема (лептотен)
постепена кондензација хромозома

Слайд 19Зигонема (зиготен)
међусобно приближавање хомологих хромозома.
Формирање хромозомских парова – БИВАЛЕНАТА.
Хромозоми се
Зигонема (зиготен)
међусобно приближавање хомологих хромозома.
Формирање хромозомских парова – БИВАЛЕНАТА.
Хромозоми се

Слайд 20Пахинема (пахитен)
оба приљубљена хромозома се деле уздужно на по две хроматиде везане
Пахинема (пахитен)
оба приљубљена хромозома се деле уздужно на по две хроматиде везане

Слайд 21Диплонема (диплотен)
Хроматиде хомологих хромозома се међусобно раздвајају (одмичу)
Остају везане преко хијазми које
Диплонема (диплотен)
Хроматиде хомологих хромозома се међусобно раздвајају (одмичу)
Остају везане преко хијазми које

Слайд 22Дијакинеза (дијакинезис)
Спирализација хромозома и терминализација хијазми достиже највећи степен
Тетраде добијају циклични облик
Једарце
Дијакинеза (дијакинезис)
Спирализација хромозома и терминализација хијазми достиже највећи степен
Тетраде добијају циклични облик
Једарце

Слайд 23Метафаза I
Тетраде се крећу према екваторијалној равни и постављају у њеној
Метафаза I
Тетраде се крећу према екваторијалној равни и постављају у њеној

Слайд 24Анфаза I
Раздвајање хомологих хромозома, кидање терминалних хијазми
Хомологи хромозоми полазе ка супротним половима
Анфаза I
Раздвајање хомологих хромозома, кидање терминалних хијазми
Хомологи хромозоми полазе ка супротним половима

Слайд 25Број хромозома се тако своди на половину, а како се биваленти раздвајају
Број хромозома се тако своди на половину, а како се биваленти раздвајају

Слайд 26Телофаза I
Два хаплоидна сета хромозома се групишу на супротним половима ћелије
Појављује се
Телофаза I
Два хаплоидна сета хромозома се групишу на супротним половима ћелије
Појављује се

Слайд 27Друга мејотичка деоба – Мејоза II
Слична је митози
Број хромозома који улази у
Друга мејотичка деоба – Мејоза II
Слична је митози
Број хромозома који улази у

Слайд 28Гамети воде порекло од примарних герминативних ћелија које се јављају у ембриону
Гамети воде порекло од примарних герминативних ћелија које се јављају у ембриону

Слайд 29Оогенеза
Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском деобом
Оогенеза
Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском деобом

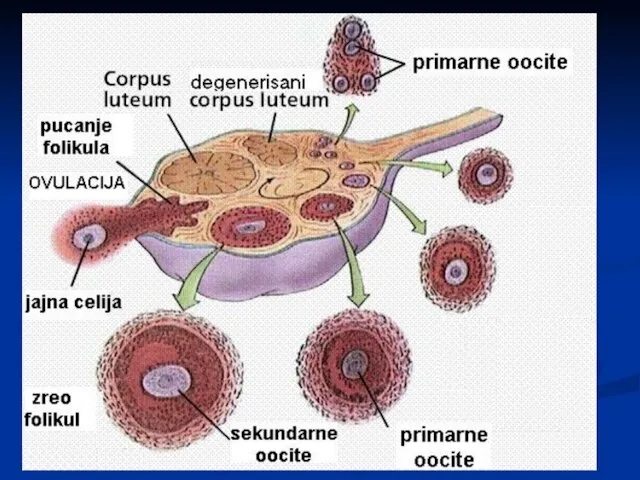
Слайд 31Оогенеза
У време рођења, у оба јајника се налази око 2 милиона примарних
Оогенеза
У време рођења, у оба јајника се налази око 2 милиона примарних

Слайд 32Оогенеза - мејоза
Примарне герминативне ћелије оваријума митозама се деле и дају ООГОНИЈЕ
Оогоније
Оогенеза - мејоза
Примарне герминативне ћелије оваријума митозама се деле и дају ООГОНИЈЕ
Оогоније

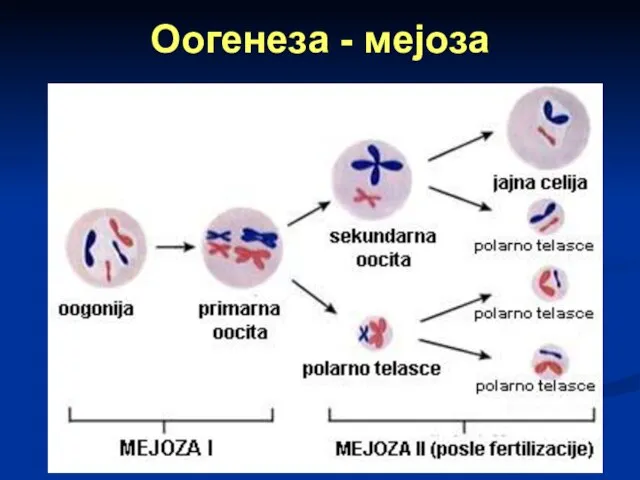
Слайд 33Оогенеза - мејоза
I ДЕОБА: ооцита I реда дели се на две неједнаке
Оогенеза - мејоза
I ДЕОБА: ооцита I реда дели се на две неједнаке

Слайд 34Оогенеза - мејоза
Оогенеза - мејоза
Слайд 35Вителогенеза
Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ
Општи назив за резервни
Вителогенеза
Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ
Општи назив за резервни

Слайд 36Типови јајних ћелија
Зависно од количине жуманцета и његове распрострањености у јајној
Типови јајних ћелија
Зависно од количине жуманцета и његове распрострањености у јајној

Слайд 37Сперматогенеза
Сперматогенеза је процес настанка сперматозоида.
Почиње у пубертету
Она се континуирано одвија
Сперматогенеза
Сперматогенеза је процес настанка сперматозоида.
Почиње у пубертету
Она се континуирано одвија

Слайд 38Улога сперматозоида
Активација јајне ћелије да отпочне са развићем
Да своју хаплоидну гарнитуру хромозома
Улога сперматозоида
Активација јајне ћелије да отпочне са развићем
Да своју хаплоидну гарнитуру хромозома

Слайд 39Сперматогенеза се одвија у 4 фазе:
СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом и
Сперматогенеза се одвија у 4 фазе:
СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом и

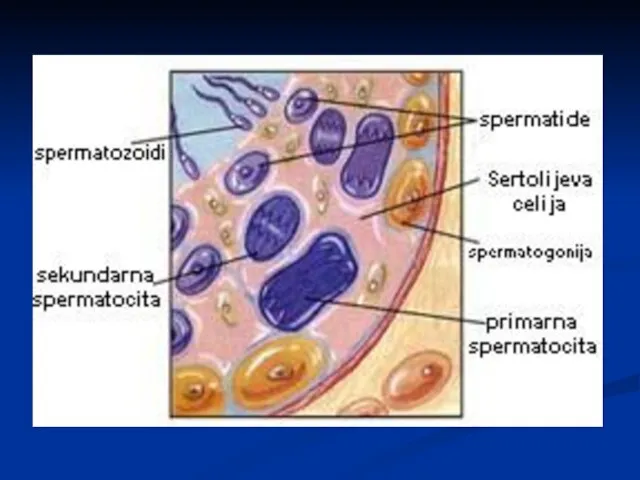
Слайд 41Сперматозоиди
Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm.
Акрозом –
Сперматозоиди
Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm.
Акрозом –

Слайд 42Сертолијеве ћелије
Исхрана исходних клициних ћелија
Ослобађање сперматозоида у лумен семених каналића
Сертолијеве ћелије
Исхрана исходних клициних ћелија
Ослобађање сперматозоида у лумен семених каналића

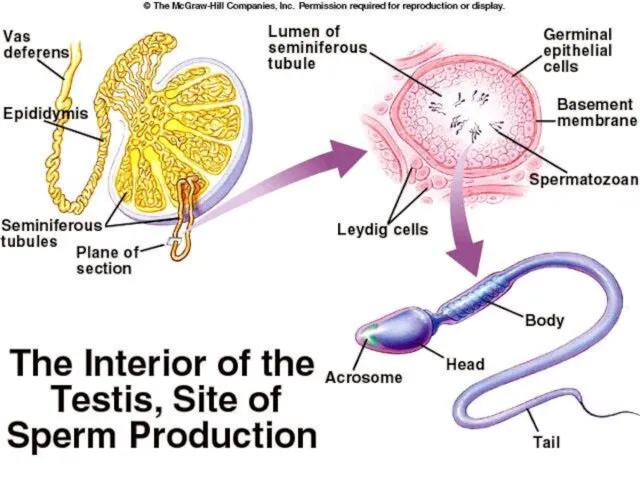
Слайд 44ОПЛОЂЕЊЕ
- Фертилизација -
Три главне функције фертилизације су:
Трансмисија гена са оба родитеља
ОПЛОЂЕЊЕ
- Фертилизација -
Три главне функције фертилизације су:
Трансмисија гена са оба родитеља

Слайд 45TIPOVI OPLOĐENJA I RAZVIĆA
U životinjskom svetu se , prema mestu odvijanja, razlikuju
TIPOVI OPLOĐENJA I RAZVIĆA
U životinjskom svetu se , prema mestu odvijanja, razlikuju

Слайд 46Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće embriona
Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće embriona

Слайд 47Фертилизација пролази кроз следеће фазе:
Контакт између сперматозоида и јајне ћелије (акрозомална реакција)
Улазак
Фертилизација пролази кроз следеће фазе:
Контакт између сперматозоида и јајне ћелије (акрозомална реакција)
Улазак

Слайд 48Aktivacija spermatozoida
počinje kada on dodirne omotač jajne ćelije što dovodi do
Aktivacija spermatozoida
počinje kada on dodirne omotač jajne ćelije što dovodi do

Слайд 49ФУЗИЈА НУКЛЕУСА
Након продора сперматозоида у јајну ћелију, долази до значајних промена у
ФУЗИЈА НУКЛЕУСА
Након продора сперматозоида у јајну ћелију, долази до значајних промена у

Слайд 50Ране деобе
(браздање)
Ране деобе
(браздање)
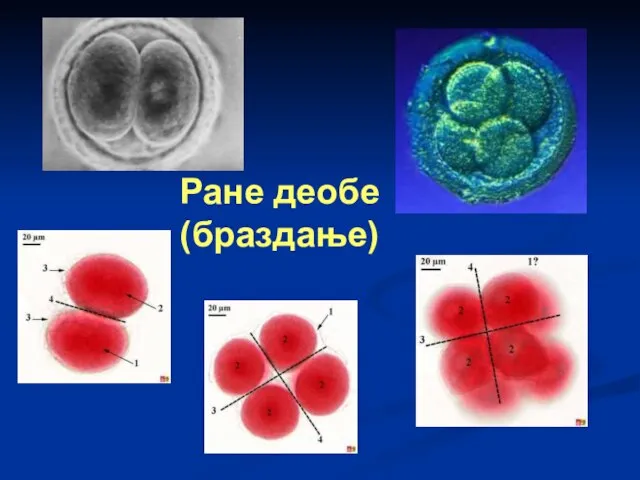
Слайд 51Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу је,
Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу је,

Слайд 52Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома брзо
Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома брзо

Слайд 53Типови браздања
Радијално браздање - на ступњу од 8 бластомера свака бластомера горњег
Типови браздања
Радијално браздање - на ступњу од 8 бластомера свака бластомера горњег

Слайд 54Жуманце има велики утицај на браздање.
Вителус смањује браздање на вегетативном полу ембриона.
Жуманце има велики утицај на браздање.
Вителус смањује браздање на вегетативном полу ембриона.

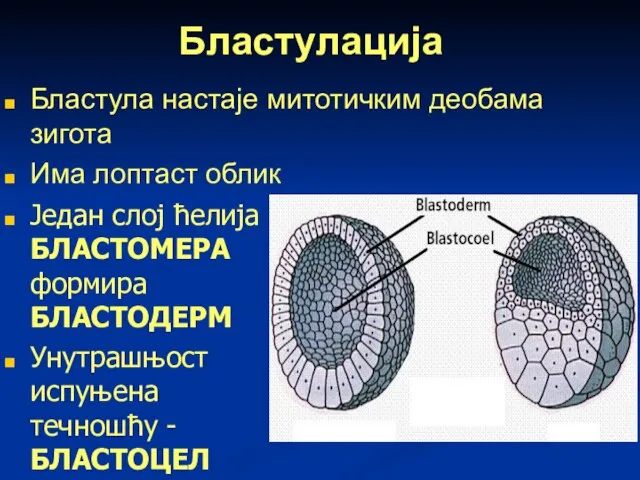
Слайд 55Бластулација
Бластула настаје митотичким деобама зигота
Има лоптаст облик
Један слој ћелија БЛАСТОМЕРА формира
Бластулација
Бластула настаје митотичким деобама зигота
Има лоптаст облик
Један слој ћелија БЛАСТОМЕРА формира
Слайд 56БЛАСТУЛА
БЛАСТУЛА
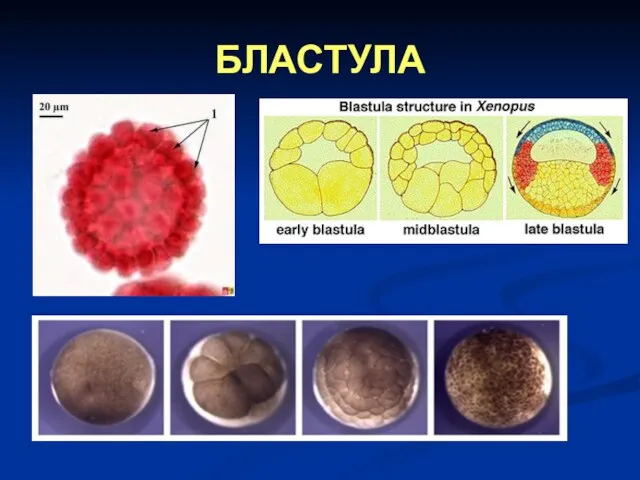
Слайд 57Типови бластула
Постоје четири типа бластула:
целобластула – то је лопта на чијој поврчини
Типови бластула
Постоје четири типа бластула:
целобластула – то је лопта на чијој поврчини

Слайд 58Сисари
Од изолецитног оплођеног јајета образује се морула, а потом бластодиск у
Сисари
Од изолецитног оплођеног јајета образује се морула, а потом бластодиск у

Слайд 59Гаструлација
Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где ће
Гаструлација
Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где ће
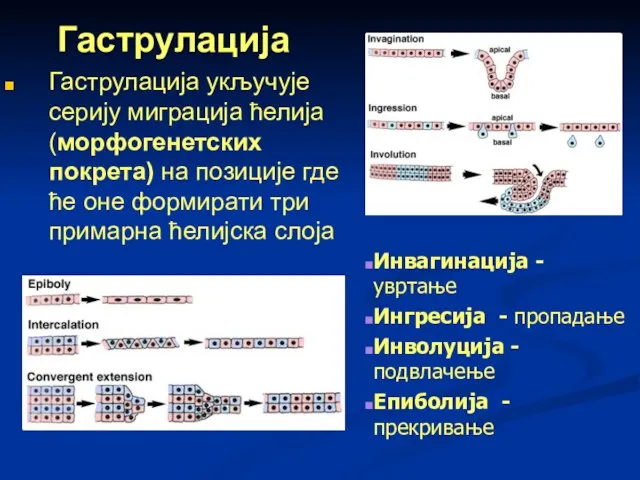
Слайд 60Начини формирања гаструле су:
инвагинација – гаструла настаје из целобластуле, која се на
Начини формирања гаструле су:
инвагинација – гаструла настаје из целобластуле, која се на

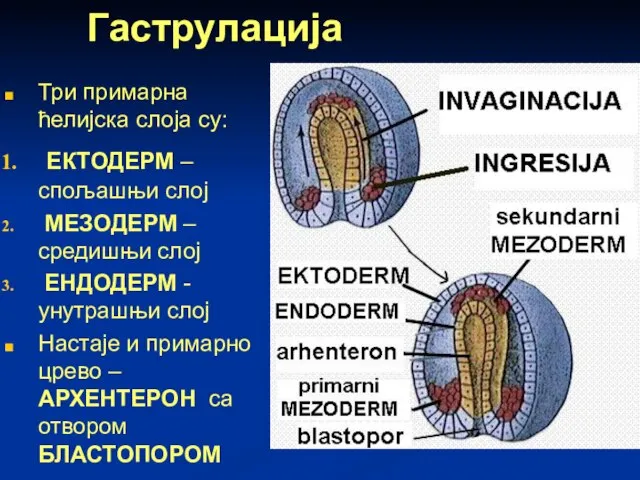
Слайд 61Гаструлација
Три примарна ћелијска слоја су:
ЕКТОДЕРМ – спољашњи слој
МЕЗОДЕРМ – средишњи
Гаструлација
Три примарна ћелијска слоја су:
ЕКТОДЕРМ – спољашњи слој
МЕЗОДЕРМ – средишњи
Слайд 62Морфогенеза – нижи хордати
Морфогенеза је прогресивно стицање телесног облика и развој одређених
Морфогенеза – нижи хордати
Морфогенеза је прогресивно стицање телесног облика и развој одређених

Слайд 63Развиће амфиоксуса
Браздањем настаје бластула
Гаструлација почиње на ступњу од око 800 ћел
Долази до
Развиће амфиоксуса
Браздањем настаје бластула
Гаструлација почиње на ступњу од око 800 ћел
Долази до

Слайд 64Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев која
Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев која

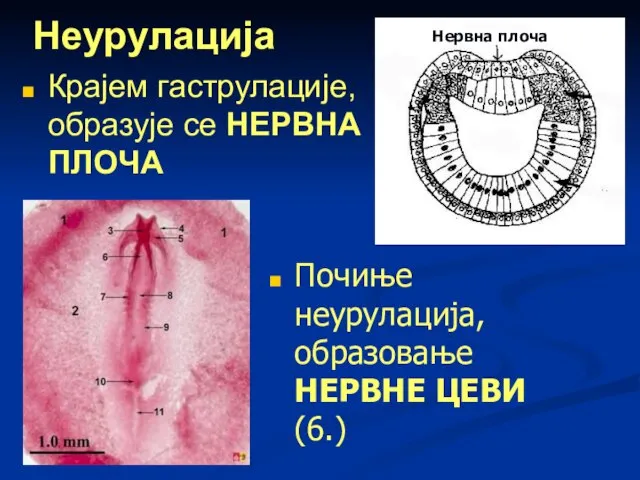
Слайд 65Неурулација
Крајем гаструлације, образује се НЕРВНА ПЛОЧА
Нервна плоча
Почиње неурулација, образовање НЕРВНЕ ЦЕВИ
Неурулација
Крајем гаструлације, образује се НЕРВНА ПЛОЧА
Нервна плоча
Почиње неурулација, образовање НЕРВНЕ ЦЕВИ
Слайд 66Развиће водоземаца
Крајем гаструлације код водоземаца се формира се нервна плоча – то
Развиће водоземаца
Крајем гаструлације код водоземаца се формира се нервна плоча – то

Слайд 67Неурулација и ембрионална индукција
На формирање организма битан утицај имају околна ткива, тј.
Неурулација и ембрионална индукција
На формирање организма битан утицај имају околна ткива, тј.

Слайд 68Околна ткива индукују настанак одређеног ткива и органа.
Тако настанак нервне цеви
Околна ткива индукују настанак одређеног ткива и органа.
Тако настанак нервне цеви

Слайд 69Органогенеза
Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом
Сва
Органогенеза
Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом
Сва

Слайд 70Сматра се да су за то одговорни градијенти информационих молекула унутар ембриона
Сматра се да су за то одговорни градијенти информационих молекула унутар ембриона

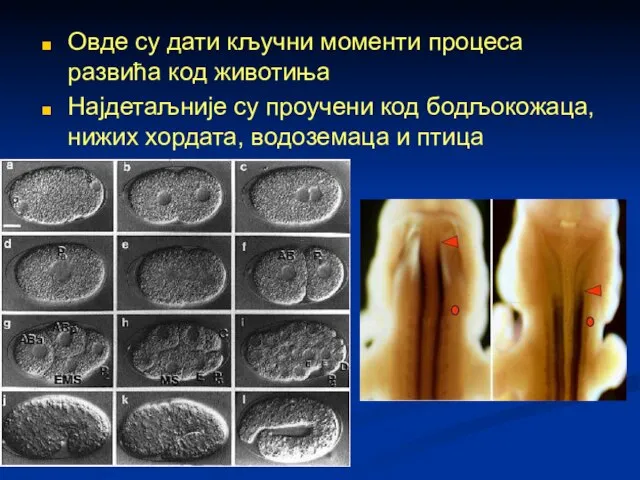
Слайд 71Овде су дати кључни моменти процеса развића код животиња
Најдетаљније су проучени код
Овде су дати кључни моменти процеса развића код животиња
Најдетаљније су проучени код
Слайд 72Развиће код различитих група
Развиће код различитих група
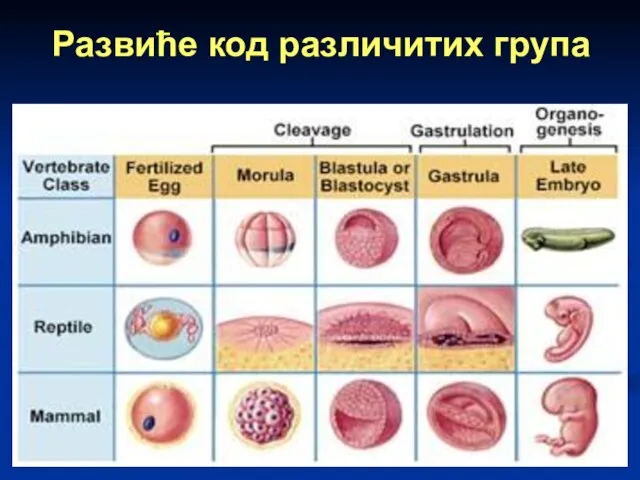
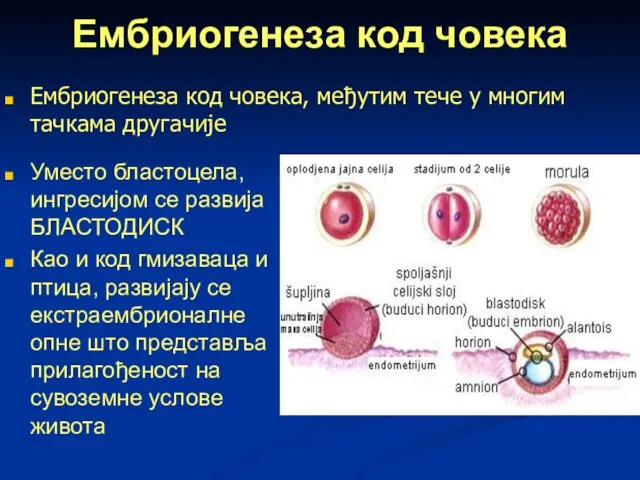
Слайд 73Ембриогенеза код човека
Уместо бластоцела, ингресијом се развија БЛАСТОДИСК
Као и код гмизаваца и
Ембриогенеза код човека
Уместо бластоцела, ингресијом се развија БЛАСТОДИСК
Као и код гмизаваца и
Слайд 74Екстраембрионалне опне
Код гмизаваца и птица
Жуманцетна кеса – садржи жуманце, има крвне судове
Екстраембрионалне опне
Код гмизаваца и птица
Жуманцетна кеса – садржи жуманце, има крвне судове

Слайд 75ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја
ОВОВИВИПАРИТЕТ – доношење на
ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја
ОВОВИВИПАРИТЕТ – доношење на

Слайд 76ПЛАЦЕНТАЦИЈА
ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива
ПО ГРАЂИ се деле
ПЛАЦЕНТАЦИЈА
ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива
ПО ГРАЂИ се деле

Слайд 77Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива
ПЛАЦЕНТАЛНА БАРИЈЕРА (слој ткива
Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива
ПЛАЦЕНТАЛНА БАРИЈЕРА (слој ткива

Слайд 78Повећањем површине контакта
Развитак наставака на површини хориона, тзв. ресица (villi)
ДИФУЗНА ПЛАЦЕНТА –
Повећањем површине контакта
Развитак наставака на површини хориона, тзв. ресица (villi)
ДИФУЗНА ПЛАЦЕНТА –

Слайд 79Смањење дебљине плаценталне баријере
Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари и
Смањење дебљине плаценталне баријере
Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари и

Слайд 80Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из ткива
Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из ткива

 Понятие автономного образовательного учреждения и его отличия от государственного и муниципального образовательного учреждени
Понятие автономного образовательного учреждения и его отличия от государственного и муниципального образовательного учреждени КОЛЫБЕЛИ НАДЕЖДЫ:право на жизнь для новорожденных в Украине
КОЛЫБЕЛИ НАДЕЖДЫ:право на жизнь для новорожденных в Украине Работа психолога в спорте-1
Работа психолога в спорте-1 Логистика как инструмент снижения издержек
Логистика как инструмент снижения издержек  СО стороны ФОМС
СО стороны ФОМС Классицизм в архитектуре Ульяновска
Классицизм в архитектуре Ульяновска Political Science — A political order
Political Science — A political order  «Феномен женщины на войне в общественном сознании» (исследовательский реферат)направление : социология
«Феномен женщины на войне в общественном сознании» (исследовательский реферат)направление : социология Проект «Моя заветная мечта»
Проект «Моя заветная мечта» ПТИЦЫ
ПТИЦЫ Лексика Антонимы Фразеологические обороты
Лексика Антонимы Фразеологические обороты Грамматическое значение причастий
Грамматическое значение причастий Чтобы не было пожара
Чтобы не было пожара Город оригами - Великий Устюг. Кадетский класс
Город оригами - Великий Устюг. Кадетский класс Элементы логики в пропедевтическом курсе информатики и ИКТ
Элементы логики в пропедевтическом курсе информатики и ИКТ Выразительно-графические свойства композиции (точка, линия, пятно)
Выразительно-графические свойства композиции (точка, линия, пятно) Защита от иска правообладателя товарного знака
Защита от иска правообладателя товарного знака Романтизм в живописи
Романтизм в живописи Создание фотоальбома в программе Windows Movie Maker
Создание фотоальбома в программе Windows Movie Maker Статистические методы в изучении предпринимательства
Статистические методы в изучении предпринимательства Общая психология. Воображение
Общая психология. Воображение Основы сетевого администрирования
Основы сетевого администрирования  представляет 4 Уникальная система для хирургии переднего и заднего отделов глаза Megatron® S4 Благодаря множеству применяемых инновац
представляет 4 Уникальная система для хирургии переднего и заднего отделов глаза Megatron® S4 Благодаря множеству применяемых инновац «Мы -за правильное питание» МБОУ «Гмелинская СОШ» 4 «А»класс
«Мы -за правильное питание» МБОУ «Гмелинская СОШ» 4 «А»класс Роль отца в формировании личности ребенка
Роль отца в формировании личности ребенка Материалы электронной техники
Материалы электронной техники БиоАгроТех. Предложение по сушке и сжиганию клеточного помета с выработкой тепла на ГВС и отопление
БиоАгроТех. Предложение по сушке и сжиганию клеточного помета с выработкой тепла на ГВС и отопление Проект выполнили ученицы 11 А класса СОШ №3 Смирнова Вера и Алексеева Оксана
Проект выполнили ученицы 11 А класса СОШ №3 Смирнова Вера и Алексеева Оксана