- Внутри и межклеточная сигнализация

Содержание
- 2. Клеточная сигнализация затрагивает все стороны биологии клетки Клеточные пути сигнализации регулируют Клеточный цикл- пролиферация Цитоскелет- миграция
- 3. Сигнальный путь это Cинтез сигнальной молекулы (2) Выделение сигнальной молекулы (3) Транспорт сигнальной молекулы к клетке
- 4. Внутриклеточный сигнальный путь – это множество участников: Белки -рецепторы Белки-реле Белки-адапторы Белки- структурные организаторы Белки- усилители
- 5. Откуда пошли сигналы Лиганды-сигналы – это побочные продукты метаболических путей , означавших избыток или недостаток источников
- 6. Что мы уже знаем о сигналах?
- 7. Что предстоит узнать
- 8. Все сигналы можно разделить на липофильные и гидрофильные Липофильные молекулы, взаимодействуют с внутриклеточными рецепторами(стероиды, ретиноиды, NO
- 10. Липофильные сигналы проникают в клетку Часто активируют гены Медленный ответ Гидрофильные сигналы не проникают в клетку
- 11. Некоторые пути обмена сигналами
- 13. Классификация молeкул-сигналов Сигнальные молекулы, построенные из аминокислот Сигналы - сложные белки (тиреотропин, гонадотропины) Сигналы - простые
- 14. Сигналы, построенные из аминокислот
- 15. Сигналы –производные аминокислот тирозина : Тироксин, трииодтиронин адреналин, норадреналин триптофана -серотонин -мелатонин
- 16. Стероиды (производные холестерола и других полиизопренов)
- 17. Производные С20 ненасыщенных жирных кислот (эйкозаноиды)
- 18. Механизмы синтеза сигнальных молекул определяются их химической структурой
- 19. Синтез производных холестерола Ацетил-КоА
- 20. Синтез производных арахидоновой кислоты см. лекции по обмену липидов.
- 21. Сигналы: активные формы кислорода (АФК) Источники : Дыхательная цепь митохондрий Микросомальное окисление (цитохромы Р450 и b5)
- 22. Сигналы: AGE (advanced glycation end products) Конечные продукты гликирования белков
- 23. Оксид азота (NO) образуется из аргинина Фермент синтаза оксида азота (NOS) содержит четыре кофермента –ФАД, ФМН,
- 24. Свойства сигналов Плейотропия – один и тот же сигнал действует на разные клетки, имеющие разные функции
- 25. Особенности сигнальной регуляции Концентрация сигналов во внеклеточной жидкости низкая - от 10-15 до 10-9 моль/л, а
- 26. Рецепторы можно разделить на две большие группы – рецепторы, встроенные в плазматическую мембрану (наиболее распространенная группа)
- 27. 1. 7-ТМС-рецепторы или R, ассоциированные с тримерными G-белками 2. 1-ТМС -рецепторы a) R, не обладающие каталитической
- 31. Рецепторы встроены в специальные участки мембран: рафты(«плоты» )
- 32. Рис. справа Кавеолин (голубой цвет) Рафты могут быть встроены в кавеолы
- 33. В создании внутриклеточных путей передачи сигнала важную роль играют реакции фосфорилирования Фосфорилирование меняет функцию белков путем
- 34. Одна из задач внутриклеточного переноса сигнала – усиление сигнала Механизмы усиления сигнала зависят от типа рецептора:
- 35. 1. 7-ТМС-рецепторы или рецепторы, ассоциированные с тримерными G-белками Среди мембранных рецепторов это наиболее распространенная группа рецепторов
- 37. Механизм работы 7-ТМС рецептора В зависимости от клеток фермент- эффектор (Е) – это: Аденилатциклаза (Gs) или
- 38. Вариант I Е – это аденилатциклаза - мембранный белок, катализирующий образование цАМФ из АТФ. Изменяет свою
- 40. Сигналы, которые активируют образование цАМФ: АКТГ, АДГ, кальцитонин, кортиколиберин, ФСГ, ЛГ, глюкагон, ТСГ, паратирин, адреналин (β-адренэрг.
- 41. Механизм действия вторичных посредников и еще раз к роли протеинкиназ
- 44. Еще один вариант работы 7 ТМС рецептора Вариант II Е – это фосфолипаза Сβ ФлСβ
- 45. Фосфолипаза Сβ -мембранный белок, катализирующий гидролиз фосфатидил инозитолфосфата с образованием ДАГ и инозитол трифосфата (ИФ3). Изменяет
- 47. CaBPs- белки, связывающие кальций
- 48. EF-hand - белки: Названы по форме, образуемой E и F α-спиралями Ca++-связывающего домена; высокое сродство к
- 50. А что ДАГ? Или может стать субстратом для получения арахидоновой кислоты, необходимой для синтеза простагландинов
- 51. Примеры вторичных посредников, образуемых с участием 7-ТМС рецепторов OPO32- Ca2+ цАМФ цГМФ И3Ф ДАГ
- 52. 2. 1-ТМС-рецепторы, не обладающие каталитической активностью, но ассоциированные с цитозольными тирозинкиназами Эти рецепторы открывают серию рецепторов,
- 53. SH2 и PTB связывают участки с фосфотирозином SH3 и WW связывают участки богатые пролином PDZ домен
- 54. Сигнальный путь с участием цитокина JAK: Янус киназа STAT: Signal transducer and activator of transcription
- 55. 3. Рецепторы, обладающие каталитической активностью Тирозинкиназы: рецепторы, фосфорилирующие тирозины молекул переноса сигнала. Тирозинфосфатазы: рецепторы, удаляющие фосфатные
- 56. Рецепторные тирозинкиназы (РТК) Подобно предыдущему классу эти рецепторы димеризуются (исключение составляют рецепторы инсулина и инсулиноподобных факторов
- 57. Соединение с лигандом Фосфорилирование СИР Присоединение ФИ-3К Образование ФИФ3 Присоединение PDK PDK фосфорилирует ПкВ и ПкСζ
- 58. 1.Связывание лиганда ? активирование тирозинкиназного домена рецептора 2. Фосфорилирование СИР 3. Присоединение белков, содержащих SH2 и
- 59. Гуанилатциклазы- бывают мембраносвязанными и растворимыми (цитоплазматическими) НФП-натрий уретический фактор предсердий
- 60. Цитоплазматическая гуанилататциклаза: Активируется NO Катализирует образование цГМФ (кишечник, гладкие мышцы сосудов) цГМФ стимулирует цГМФ-зависимые протеинкиназы (ПкG)
- 61. 4.Ядерные и цитозольные рецепторы Ядерные рецепторы – факторы транскрипции (TF) опосредуют изменение транскрипции в ответ на
- 63. 5 . Рецепторы – лигандзависимые ионные каналы
- 64. Обеспечивают прямое взаимодействие между клетками I. Замыкающие (плотные) контакты II. Прикрепительные соединения 1. c актиновыми филаментами
- 65. Различают Ca2+ - зависимые CAM: кадгерины, селектины, интегрины 2)Ca2+ - независимые CAM Клетки связываются и общаются
- 66. Интегрины обеспечиают СИГНАЛИЗАЦИЮ ИЗ ВНЕКЛЕТОЧНОЙ СРЕДЫ: Внеклеточныый домен связывается с лигандом (ММ или CAM) ? Конформационные
- 67. Секвестрация рецепторов Модификация рецепторов (фосфорилирование) Инактивация белков участвующих в переносе (фосфатазы) Белки- «реле» Изменение концентрации вторичных
- 69. Скачать презентацию
Слайд 2Клеточная сигнализация затрагивает все стороны биологии клетки
Клеточные пути сигнализации регулируют
Клеточный цикл-
Клеточная сигнализация затрагивает все стороны биологии клетки
Клеточные пути сигнализации регулируют
Клеточный цикл-
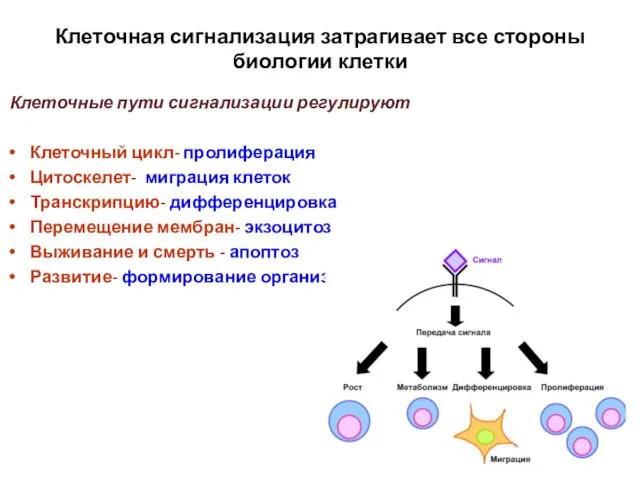
Слайд 3Сигнальный путь это
Cинтез сигнальной молекулы
(2) Выделение сигнальной молекулы
(3) Транспорт
Сигнальный путь это
Cинтез сигнальной молекулы
(2) Выделение сигнальной молекулы
(3) Транспорт
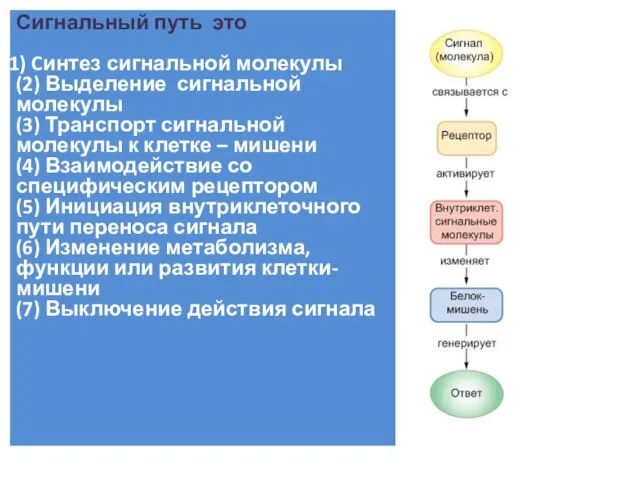
Слайд 4Внутриклеточный сигнальный путь – это множество участников:
Белки -рецепторы
Белки-реле
Белки-адапторы
Белки- структурные организаторы
Белки- усилители и
Внутриклеточный сигнальный путь – это множество участников:
Белки -рецепторы
Белки-реле
Белки-адапторы
Белки- структурные организаторы
Белки- усилители и
Слайд 5Откуда пошли сигналы
Лиганды-сигналы – это побочные продукты
метаболических путей , означавших
Откуда пошли сигналы
Лиганды-сигналы – это побочные продукты
метаболических путей , означавших

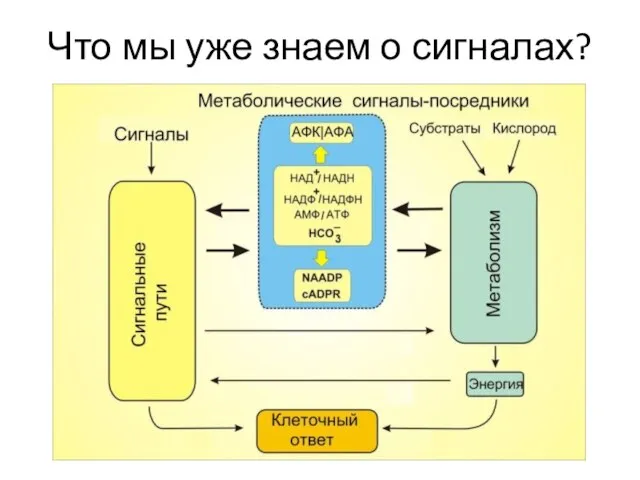
Слайд 6Что мы уже знаем о сигналах?
Что мы уже знаем о сигналах?
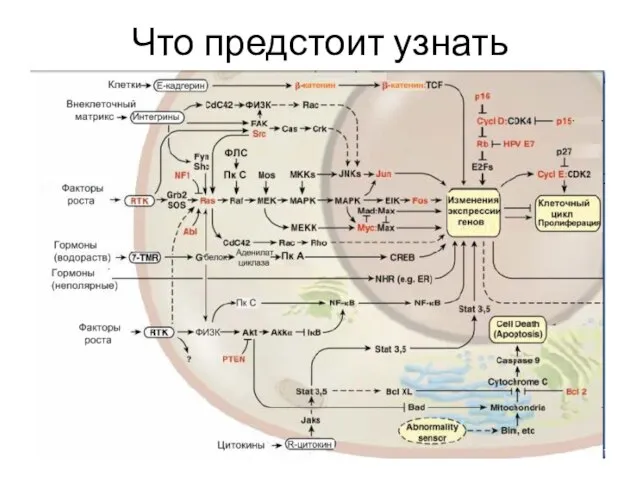
Слайд 7Что предстоит узнать
Что предстоит узнать
Слайд 8 Все сигналы можно разделить на липофильные и гидрофильные
Липофильные молекулы, взаимодействуют с
Все сигналы можно разделить на липофильные и гидрофильные
Липофильные молекулы, взаимодействуют с


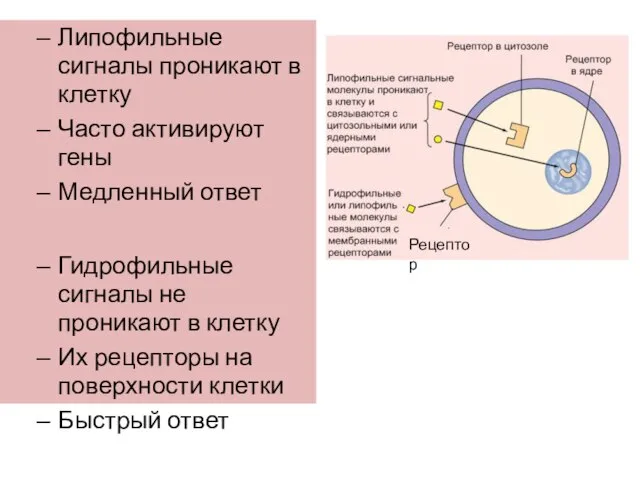
Слайд 10Липофильные сигналы проникают в клетку
Часто активируют гены
Медленный ответ
Гидрофильные сигналы не проникают
Липофильные сигналы проникают в клетку
Часто активируют гены
Медленный ответ
Гидрофильные сигналы не проникают
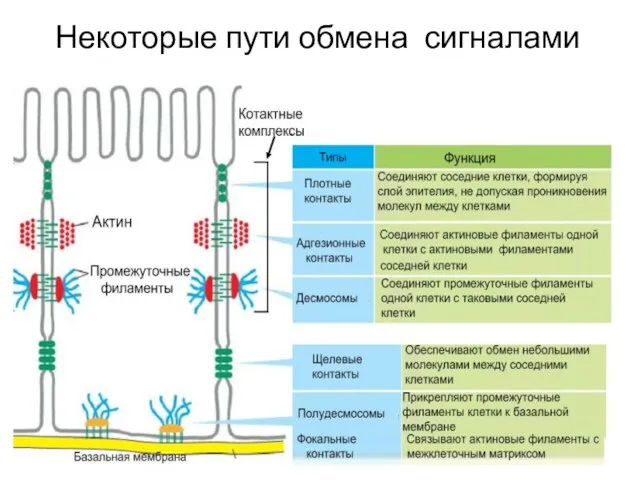
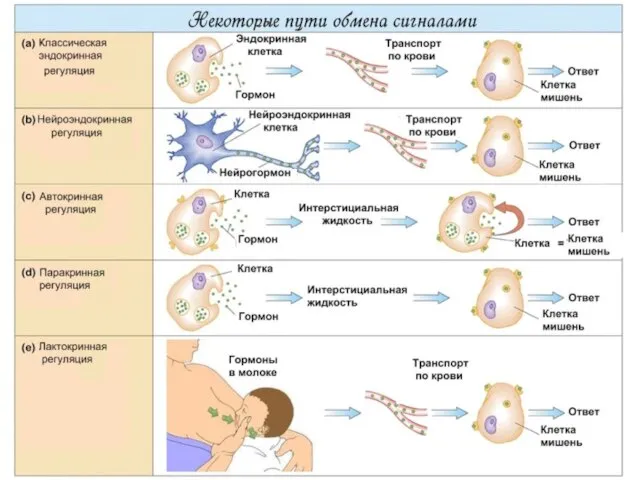
Слайд 11Некоторые пути обмена сигналами
Некоторые пути обмена сигналами
Слайд 13Классификация молeкул-сигналов
Сигнальные молекулы, построенные из аминокислот
Сигналы - сложные белки (тиреотропин,
Классификация молeкул-сигналов
Сигнальные молекулы, построенные из аминокислот
Сигналы - сложные белки (тиреотропин,

Слайд 14Сигналы, построенные из аминокислот
Сигналы, построенные из аминокислот
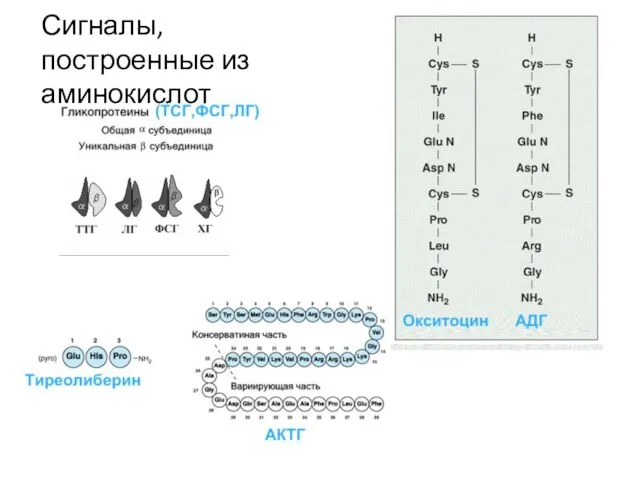
Слайд 15Сигналы –производные аминокислот
тирозина :
Тироксин,
трииодтиронин
адреналин,
норадреналин
триптофана
-серотонин
-мелатонин
Сигналы –производные аминокислот
тирозина :
Тироксин,
трииодтиронин
адреналин,
норадреналин
триптофана
-серотонин
-мелатонин
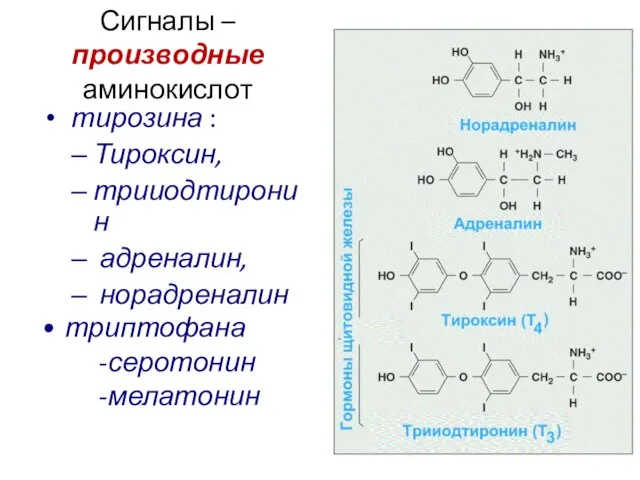
Слайд 16Стероиды (производные холестерола и других полиизопренов)
Стероиды (производные холестерола и других полиизопренов)
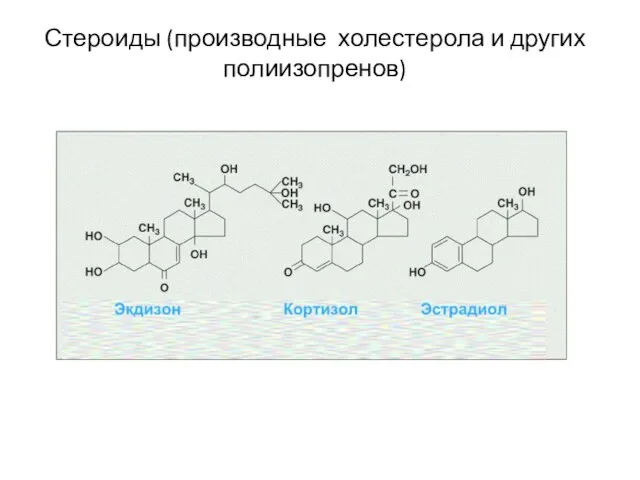
Слайд 17Производные С20 ненасыщенных жирных кислот (эйкозаноиды)
Производные С20 ненасыщенных жирных кислот (эйкозаноиды)
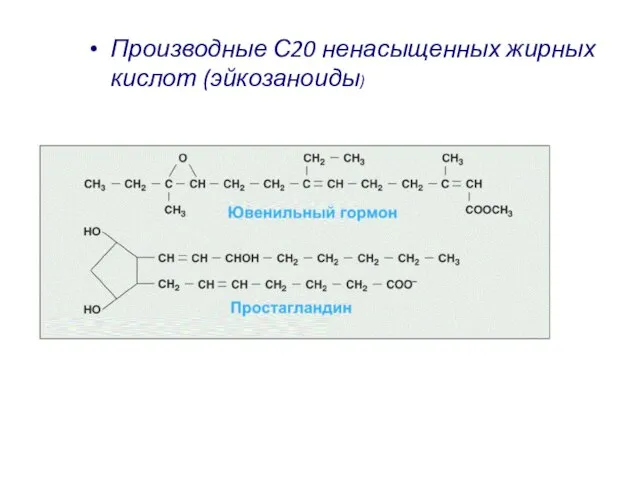
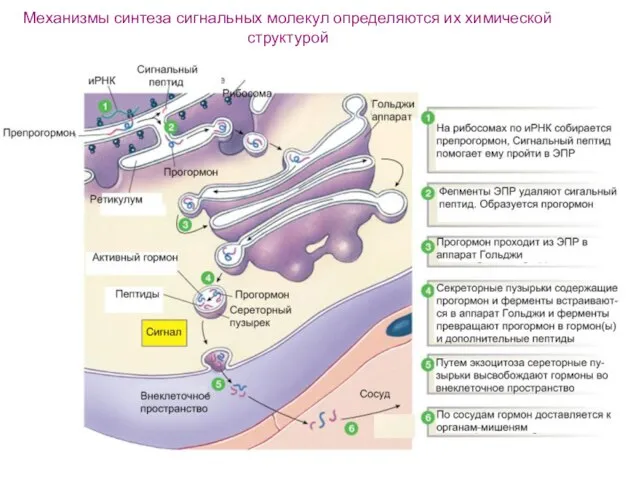
Слайд 18Механизмы синтеза сигнальных молекул определяются их химической структурой
Механизмы синтеза сигнальных молекул определяются их химической структурой
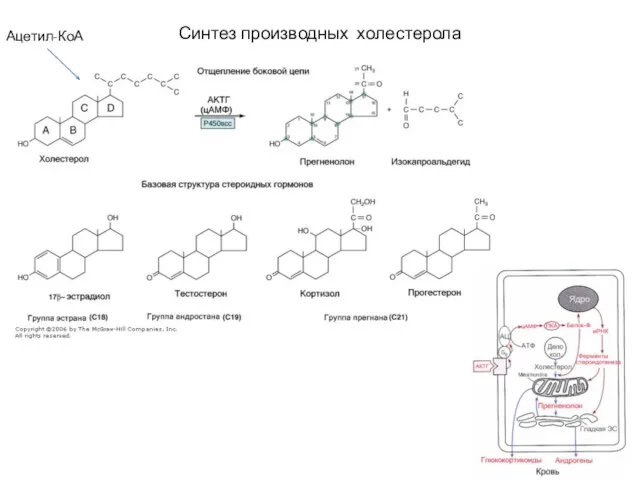
Слайд 19Синтез производных холестерола
Ацетил-КоА
Синтез производных холестерола
Ацетил-КоА
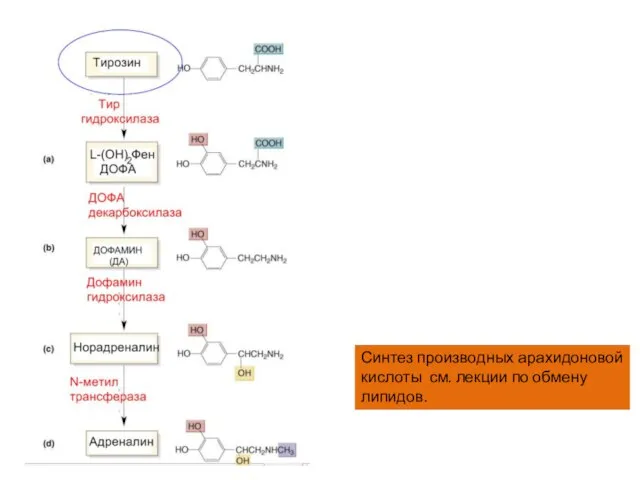
Слайд 20Синтез производных арахидоновой кислоты см. лекции по обмену липидов.
Синтез производных арахидоновой кислоты см. лекции по обмену липидов.
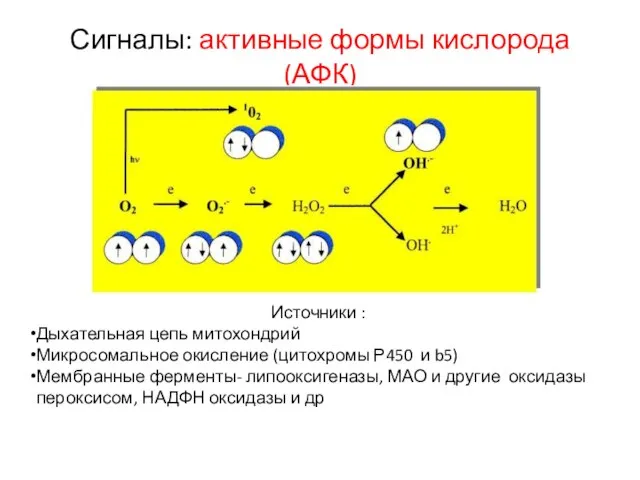
Слайд 21Сигналы: активные формы кислорода (АФК)
Источники :
Дыхательная цепь митохондрий
Микросомальное окисление (цитохромы Р450 и
Сигналы: активные формы кислорода (АФК)
Источники :
Дыхательная цепь митохондрий
Микросомальное окисление (цитохромы Р450 и
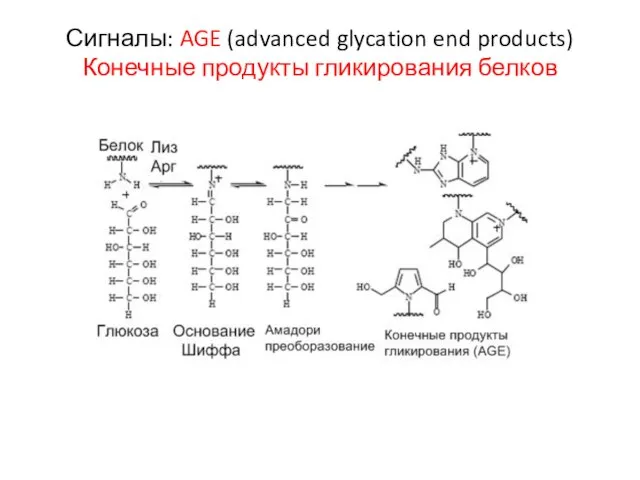
Слайд 22Сигналы: AGE (advanced glycation end products) Конечные продукты гликирования белков
Сигналы: AGE (advanced glycation end products) Конечные продукты гликирования белков
Слайд 23Оксид азота (NO) образуется из аргинина
Фермент синтаза оксида азота (NOS) содержит
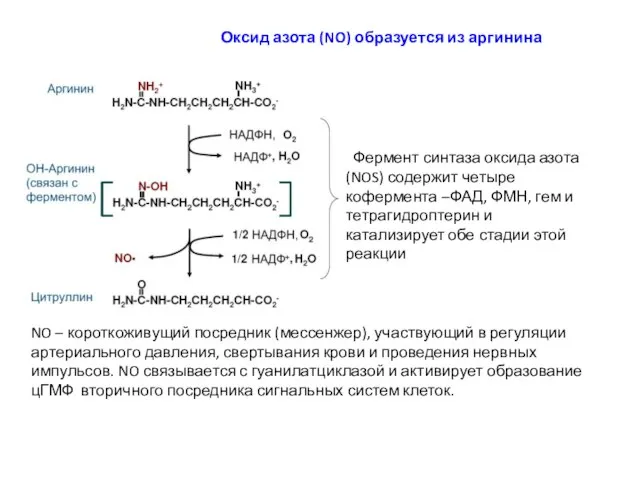
Оксид азота (NO) образуется из аргинина
Фермент синтаза оксида азота (NOS) содержит
Слайд 24Свойства сигналов
Плейотропия – один и тот же сигнал действует на разные клетки,
Свойства сигналов
Плейотропия – один и тот же сигнал действует на разные клетки,

Слайд 25Особенности сигнальной регуляции
Концентрация сигналов во внеклеточной жидкости низкая - от 10-15
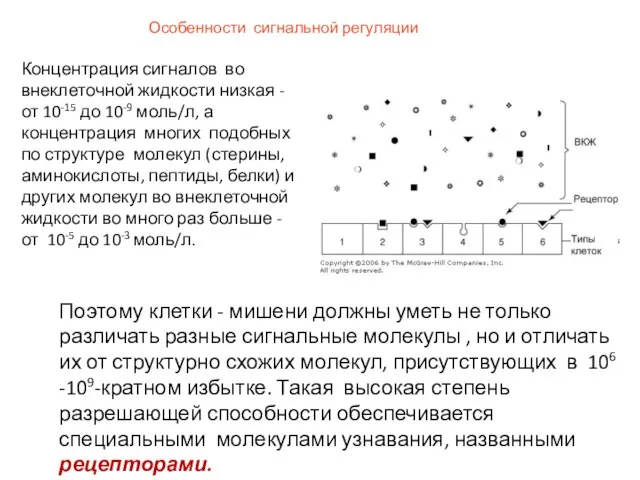
Особенности сигнальной регуляции
Концентрация сигналов во внеклеточной жидкости низкая - от 10-15
Слайд 26Рецепторы можно разделить на две большие группы – рецепторы, встроенные в плазматическую
Рецепторы можно разделить на две большие группы – рецепторы, встроенные в плазматическую

Слайд 271. 7-ТМС-рецепторы или R, ассоциированные с тримерными G-белками
2. 1-ТМС -рецепторы
1. 7-ТМС-рецепторы или R, ассоциированные с тримерными G-белками
2. 1-ТМС -рецепторы

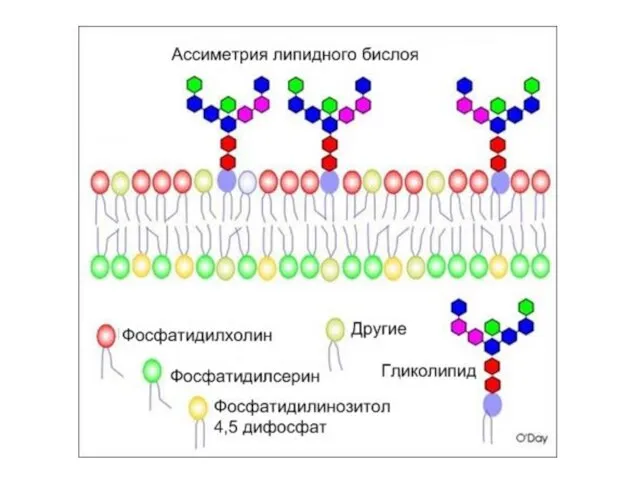
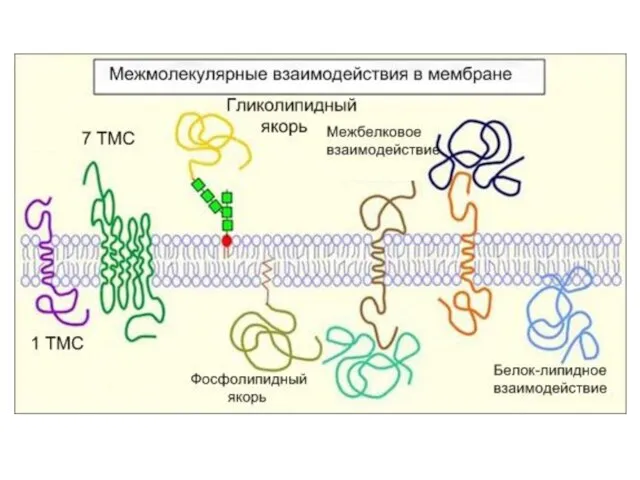
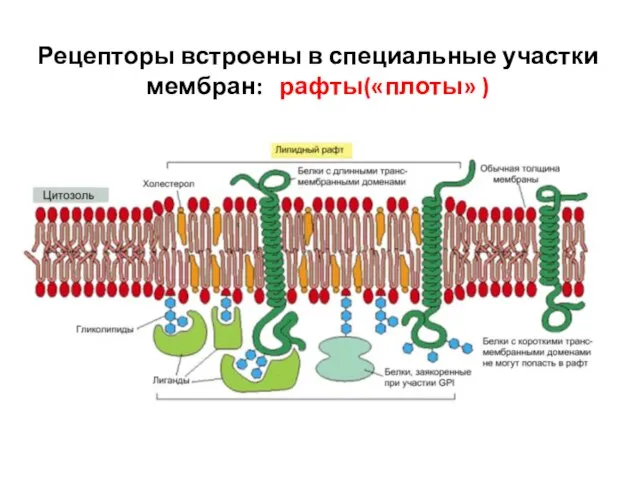
Слайд 31Рецепторы встроены в специальные участки мембран: рафты(«плоты» )
Рецепторы встроены в специальные участки мембран: рафты(«плоты» )
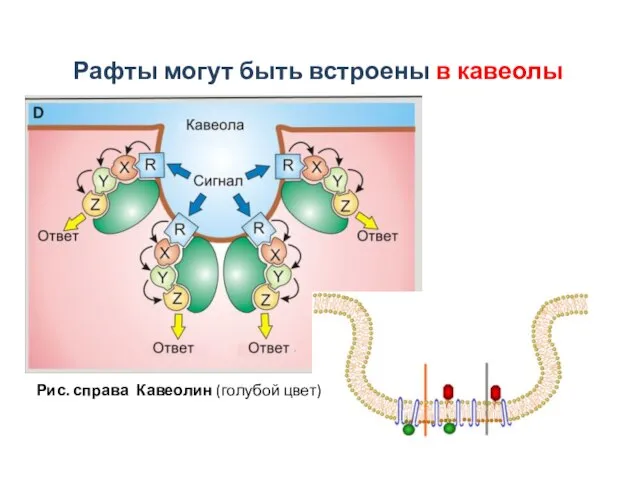
Слайд 32Рис. справа Кавеолин (голубой цвет)
Рафты могут быть встроены в кавеолы
Рис. справа Кавеолин (голубой цвет)
Рафты могут быть встроены в кавеолы
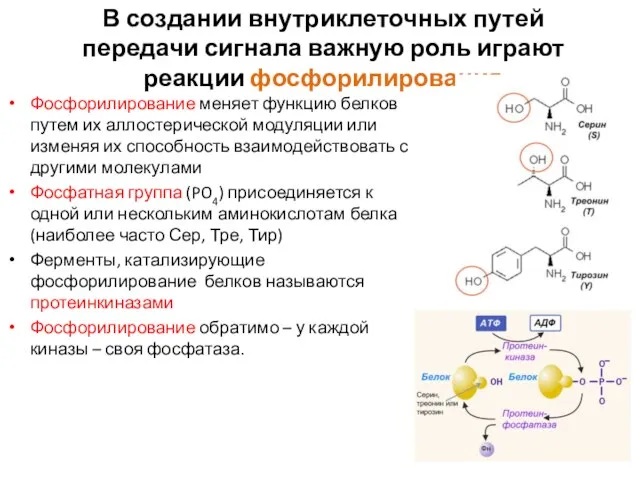
Слайд 33В создании внутриклеточных путей передачи сигнала важную роль играют реакции фосфорилирования
Фосфорилирование меняет
В создании внутриклеточных путей передачи сигнала важную роль играют реакции фосфорилирования
Фосфорилирование меняет
Слайд 34Одна из задач внутриклеточного переноса сигнала – усиление сигнала
Механизмы усиления сигнала зависят
Одна из задач внутриклеточного переноса сигнала – усиление сигнала
Механизмы усиления сигнала зависят

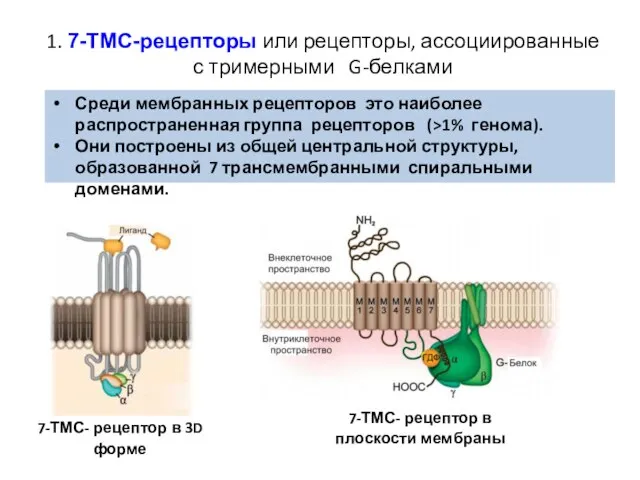
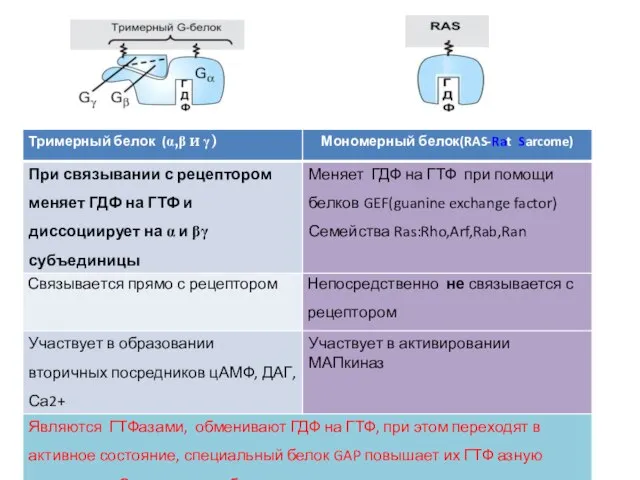
Слайд 351. 7-ТМС-рецепторы или рецепторы, ассоциированные с тримерными G-белками
Среди мембранных рецепторов это
1. 7-ТМС-рецепторы или рецепторы, ассоциированные с тримерными G-белками
Среди мембранных рецепторов это
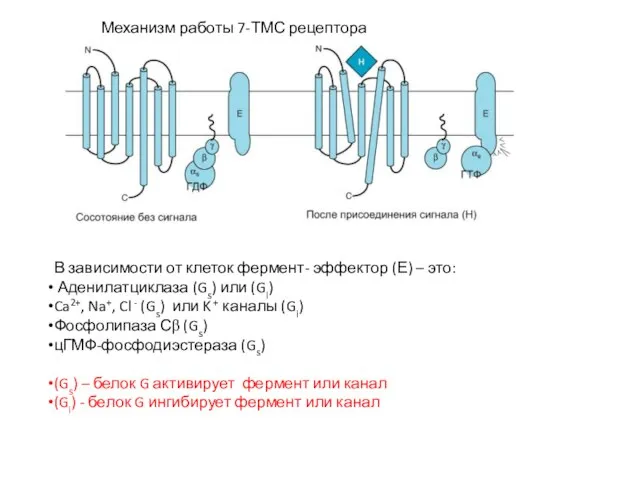
Слайд 37Механизм работы 7-ТМС рецептора
В зависимости от клеток фермент- эффектор (Е) –
Механизм работы 7-ТМС рецептора
В зависимости от клеток фермент- эффектор (Е) –
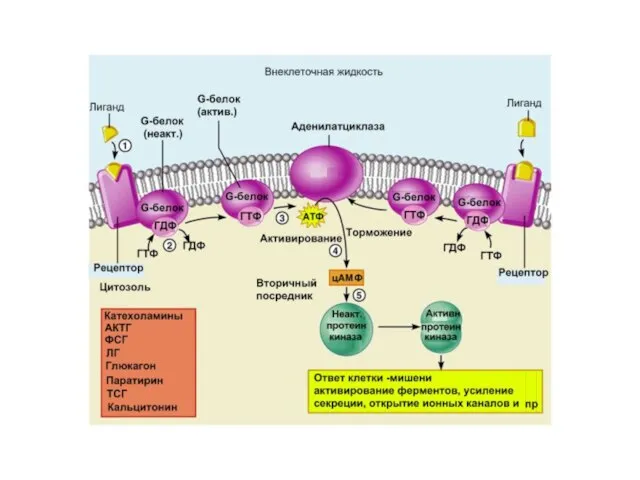
Слайд 38 Вариант I Е – это аденилатциклаза - мембранный белок, катализирующий образование
Вариант I Е – это аденилатциклаза - мембранный белок, катализирующий образование
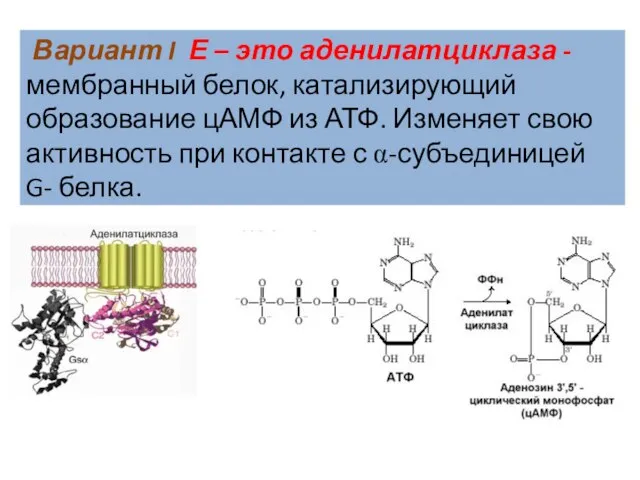
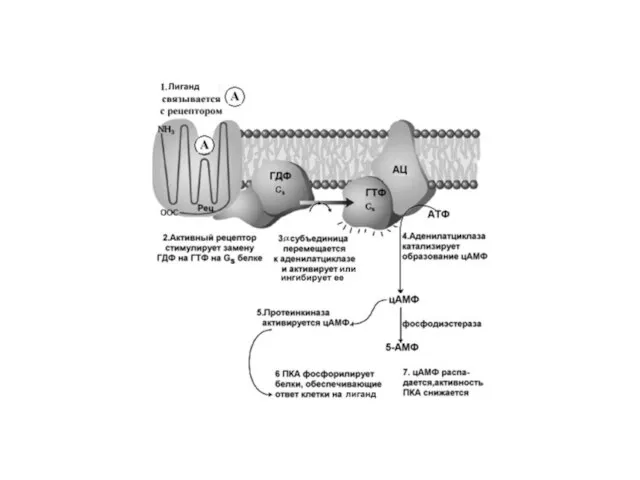
Слайд 40Сигналы, которые активируют образование цАМФ:
АКТГ, АДГ, кальцитонин, кортиколиберин, ФСГ, ЛГ, глюкагон, ТСГ,
Сигналы, которые активируют образование цАМФ:
АКТГ, АДГ, кальцитонин, кортиколиберин, ФСГ, ЛГ, глюкагон, ТСГ,
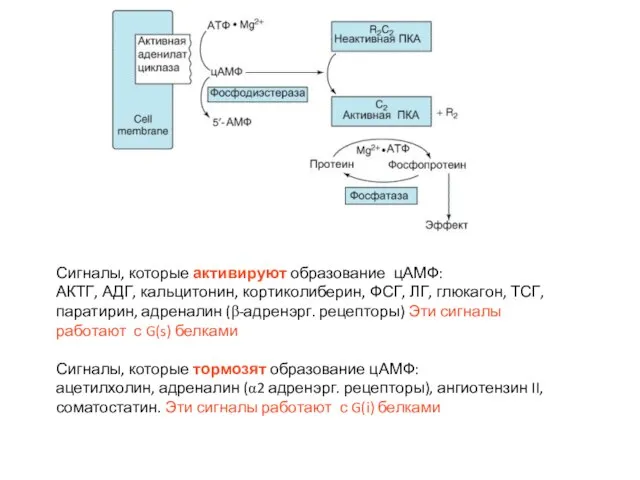
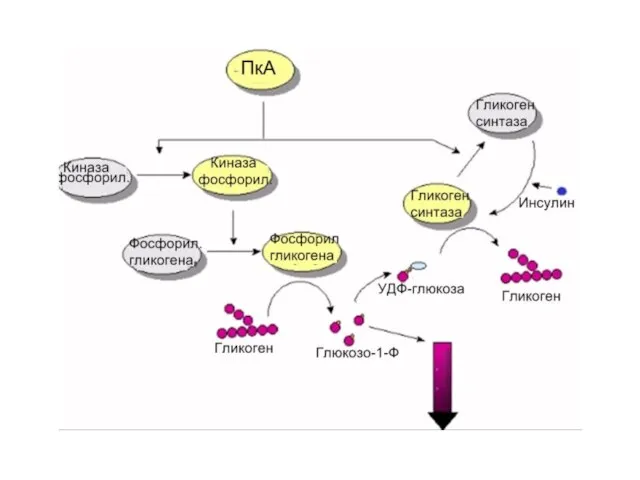
Слайд 41Механизм действия вторичных посредников и еще раз к роли протеинкиназ
Механизм действия вторичных посредников и еще раз к роли протеинкиназ
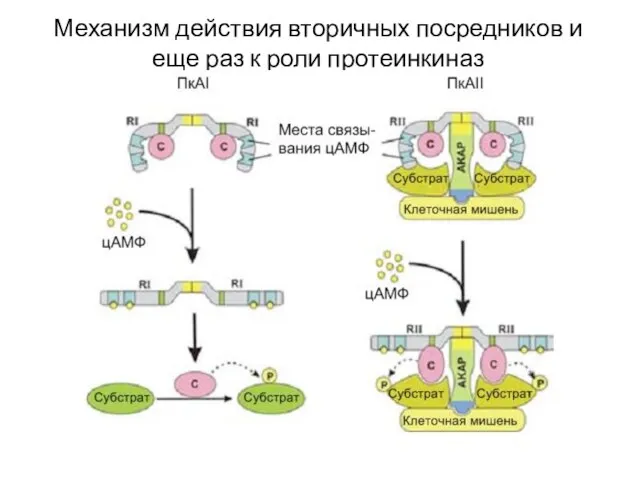
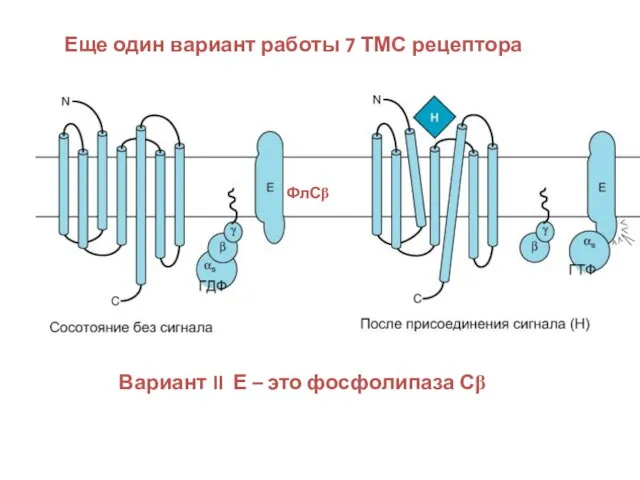
Слайд 44Еще один вариант работы 7 ТМС рецептора
Вариант II Е – это
Еще один вариант работы 7 ТМС рецептора
Вариант II Е – это
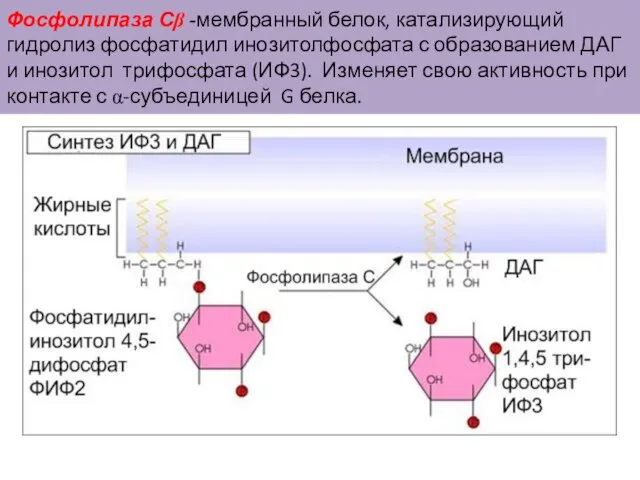
Слайд 45Фосфолипаза Сβ -мембранный белок, катализирующий гидролиз фосфатидил инозитолфосфата с образованием ДАГ и
Фосфолипаза Сβ -мембранный белок, катализирующий гидролиз фосфатидил инозитолфосфата с образованием ДАГ и

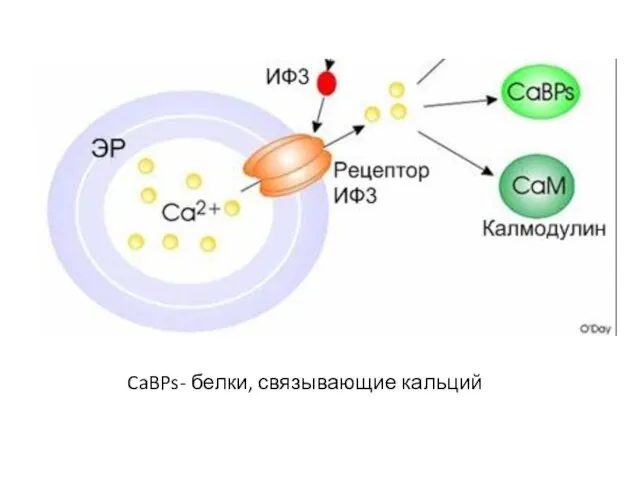
Слайд 47CaBPs- белки, связывающие кальций
CaBPs- белки, связывающие кальций
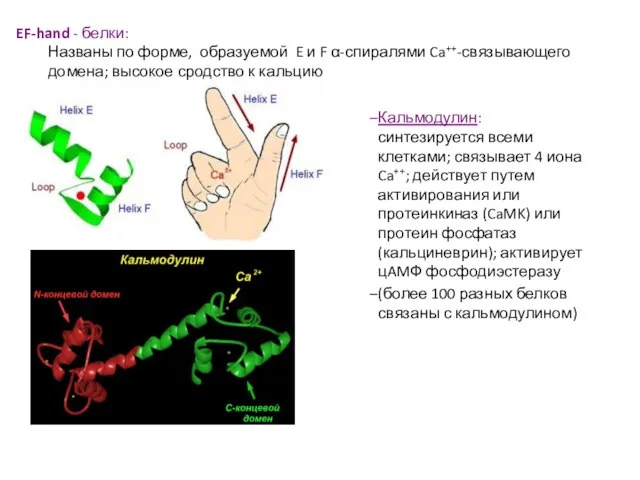
Слайд 48EF-hand - белки:
Названы по форме, образуемой E и F α-спиралями Ca++-связывающего домена;
EF-hand - белки:
Названы по форме, образуемой E и F α-спиралями Ca++-связывающего домена;

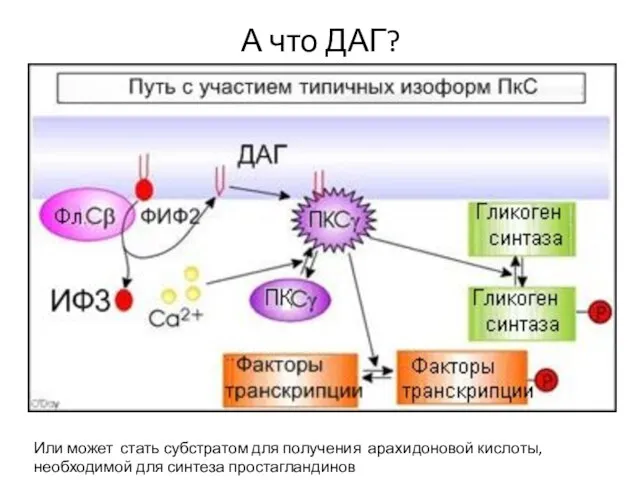
Слайд 50А что ДАГ?
Или может стать субстратом для получения арахидоновой кислоты, необходимой для
А что ДАГ?
Или может стать субстратом для получения арахидоновой кислоты, необходимой для
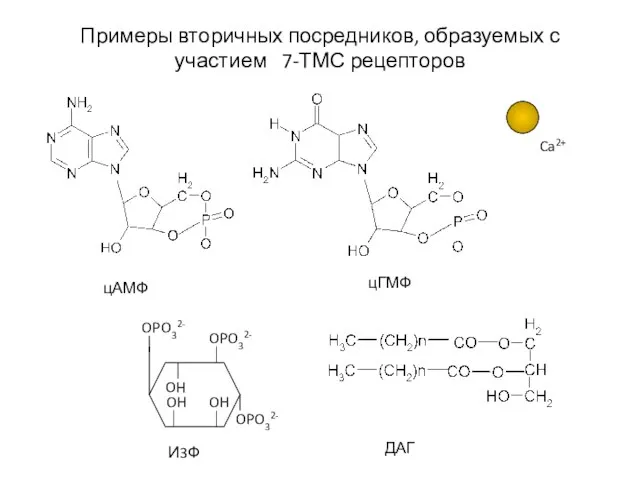
Слайд 51Примеры вторичных посредников, образуемых с участием 7-ТМС рецепторов
OPO32-
Ca2+
цАМФ
цГМФ
И3Ф
ДАГ
Примеры вторичных посредников, образуемых с участием 7-ТМС рецепторов
OPO32-
Ca2+
цАМФ
цГМФ
И3Ф
ДАГ
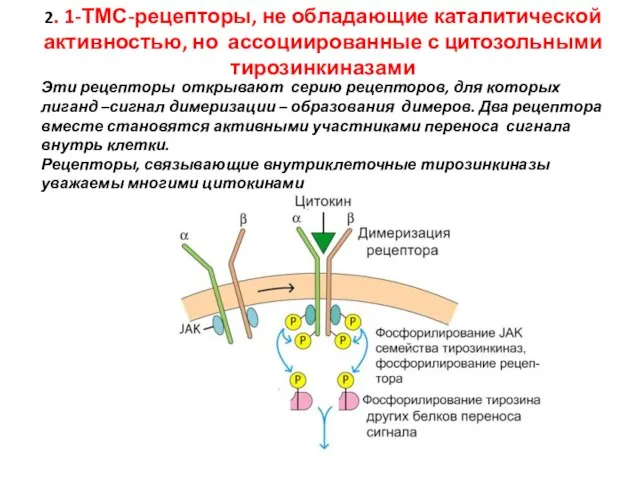
Слайд 522. 1-ТМС-рецепторы, не обладающие каталитической активностью, но ассоциированные с цитозольными тирозинкиназами
Эти рецепторы
2. 1-ТМС-рецепторы, не обладающие каталитической активностью, но ассоциированные с цитозольными тирозинкиназами
Эти рецепторы
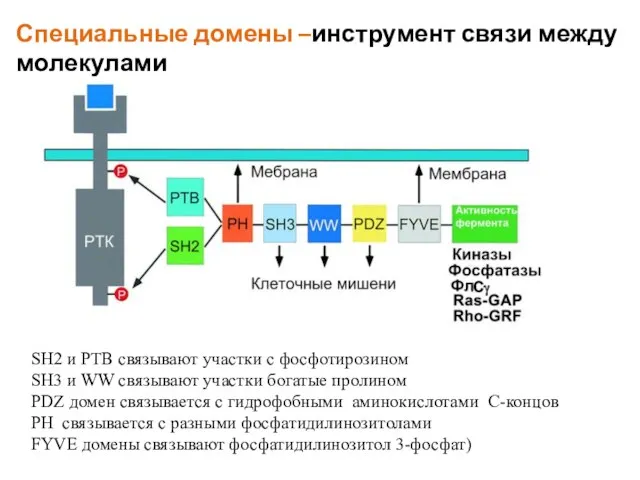
Слайд 53SH2 и PTB связывают участки с фосфотирозином
SH3 и WW связывают участки богатые
SH2 и PTB связывают участки с фосфотирозином
SH3 и WW связывают участки богатые
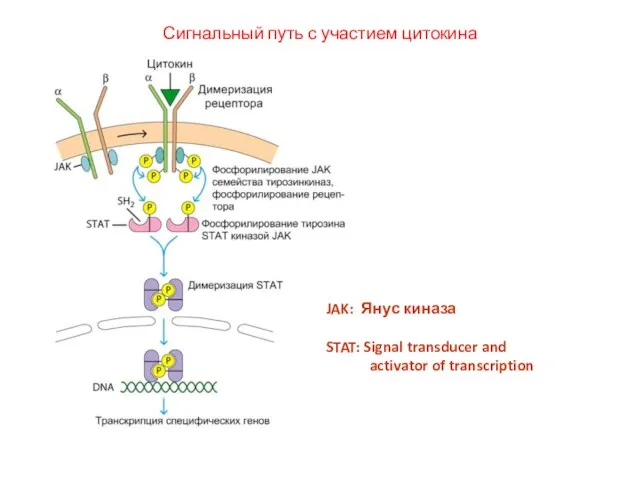
Слайд 54Сигнальный путь с участием цитокина
JAK: Янус киназа
STAT: Signal transducer and
activator
Сигнальный путь с участием цитокина
JAK: Янус киназа
STAT: Signal transducer and
activator
Слайд 553. Рецепторы, обладающие каталитической активностью
Тирозинкиназы: рецепторы, фосфорилирующие тирозины молекул переноса сигнала.
Тирозинфосфатазы: рецепторы,
3. Рецепторы, обладающие каталитической активностью
Тирозинкиназы: рецепторы, фосфорилирующие тирозины молекул переноса сигнала.
Тирозинфосфатазы: рецепторы,

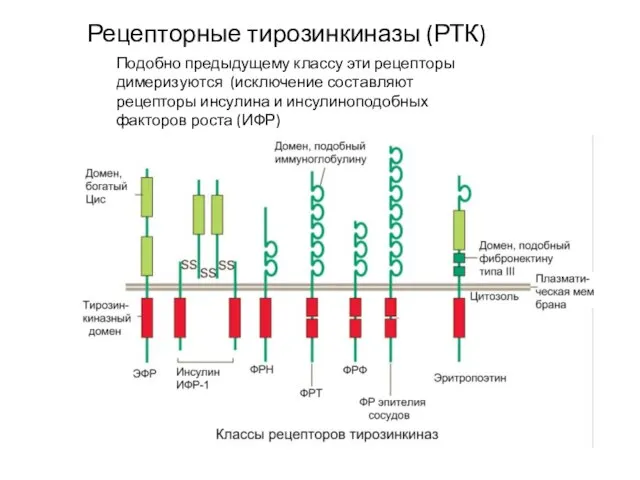
Слайд 56Рецепторные тирозинкиназы (РТК)
Подобно предыдущему классу эти рецепторы димеризуются (исключение составляют рецепторы инсулина
Рецепторные тирозинкиназы (РТК)
Подобно предыдущему классу эти рецепторы димеризуются (исключение составляют рецепторы инсулина
Слайд 57Соединение с лигандом
Фосфорилирование СИР
Присоединение ФИ-3К
Образование ФИФ3
Присоединение PDK
PDK фосфорилирует ПкВ и ПкСζ
ПкВ фосфорилирует
Соединение с лигандом
Фосфорилирование СИР
Присоединение ФИ-3К
Образование ФИФ3
Присоединение PDK
PDK фосфорилирует ПкВ и ПкСζ
ПкВ фосфорилирует
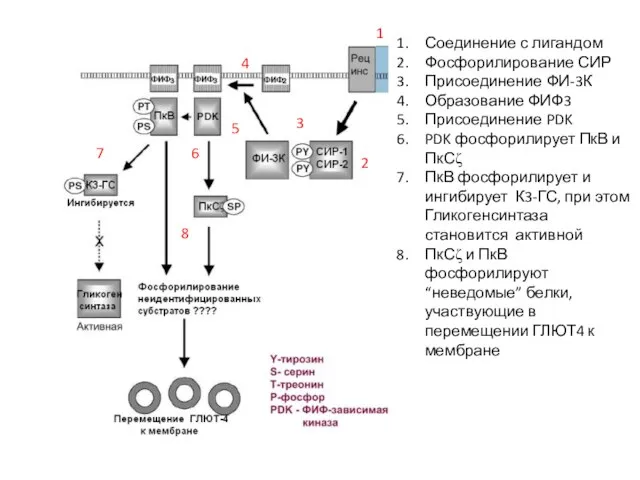
Слайд 581.Связывание лиганда ? активирование тирозинкиназного домена рецептора
2. Фосфорилирование СИР
3. Присоединение белков, содержащих
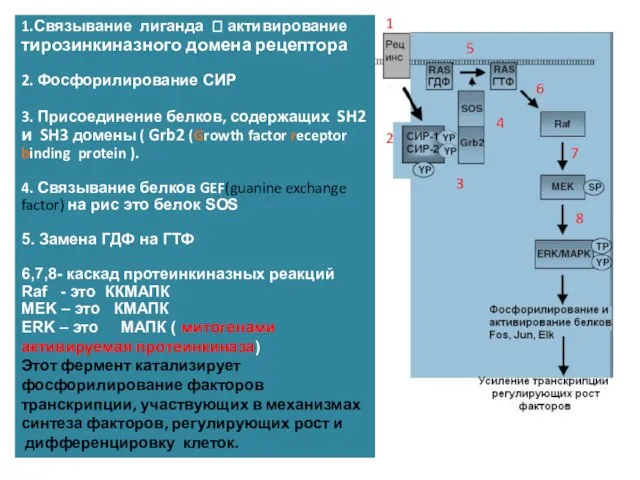
1.Связывание лиганда ? активирование тирозинкиназного домена рецептора
2. Фосфорилирование СИР
3. Присоединение белков, содержащих
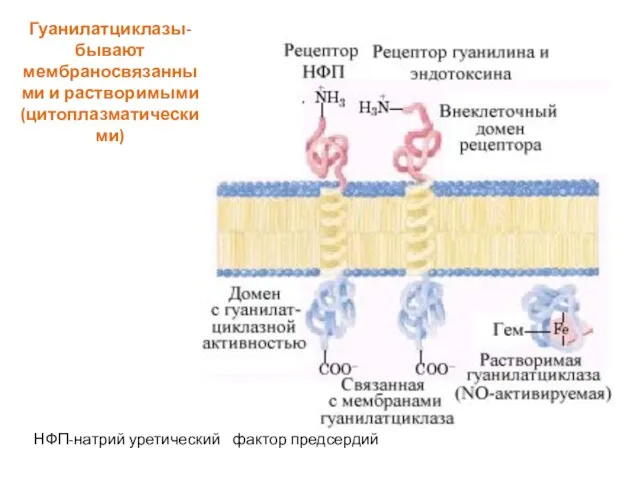
Слайд 59Гуанилатциклазы-
бывают мембраносвязанными и растворимыми (цитоплазматическими)
НФП-натрий уретический фактор предсердий
Гуанилатциклазы-
бывают мембраносвязанными и растворимыми (цитоплазматическими)
НФП-натрий уретический фактор предсердий
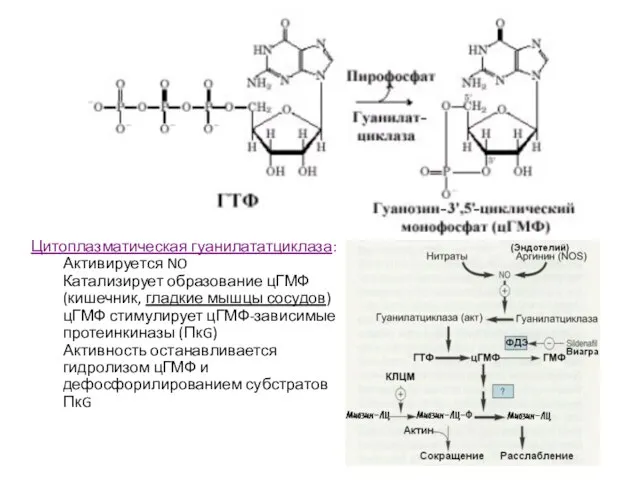
Слайд 60Цитоплазматическая гуанилататциклаза:
Активируется NO
Катализирует образование цГМФ
(кишечник, гладкие мышцы сосудов)
цГМФ стимулирует цГМФ-зависимые протеинкиназы (ПкG)
Активность
Цитоплазматическая гуанилататциклаза:
Активируется NO
Катализирует образование цГМФ
(кишечник, гладкие мышцы сосудов)
цГМФ стимулирует цГМФ-зависимые протеинкиназы (ПкG)
Активность
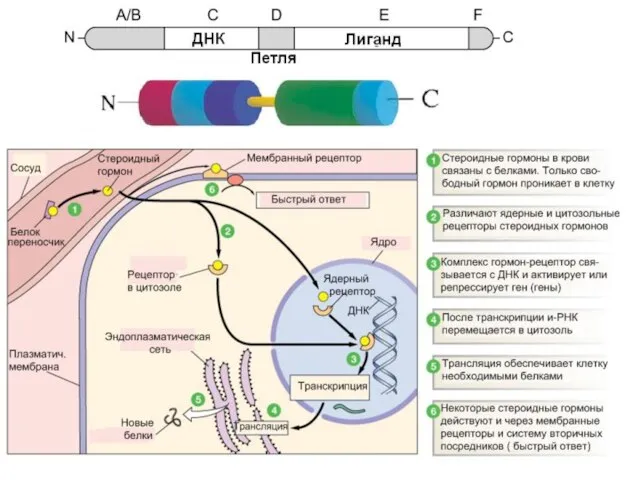
Слайд 614.Ядерные и цитозольные рецепторы
Ядерные рецепторы – факторы транскрипции (TF)
опосредуют изменение транскрипции
4.Ядерные и цитозольные рецепторы
Ядерные рецепторы – факторы транскрипции (TF)
опосредуют изменение транскрипции

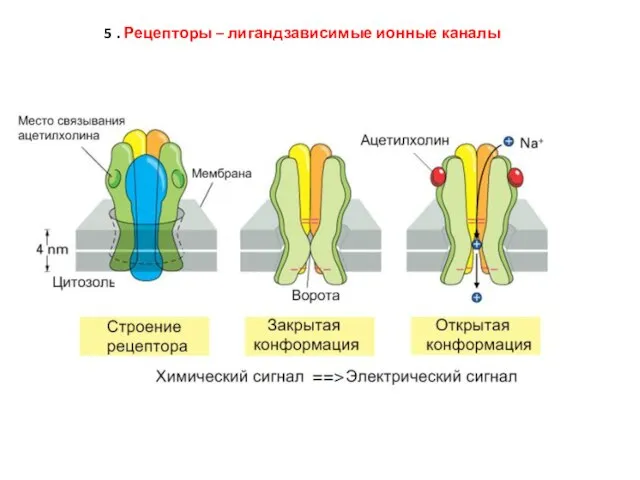
Слайд 635 . Рецепторы – лигандзависимые ионные каналы
5 . Рецепторы – лигандзависимые ионные каналы
Слайд 64Обеспечивают прямое взаимодействие между клетками
I. Замыкающие (плотные) контакты
II. Прикрепительные соединения
1. c актиновыми
Обеспечивают прямое взаимодействие между клетками
I. Замыкающие (плотные) контакты
II. Прикрепительные соединения
1. c актиновыми

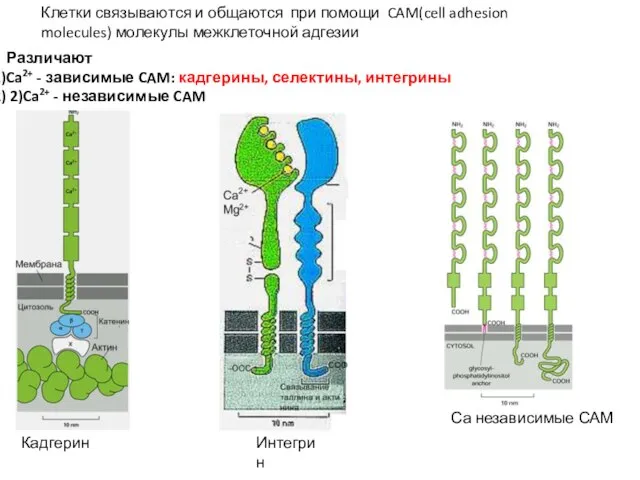
Слайд 65Различают
Ca2+ - зависимые CAM: кадгерины, селектины, интегрины
2)Ca2+ - независимые CAM
Клетки связываются
Различают
Ca2+ - зависимые CAM: кадгерины, селектины, интегрины
2)Ca2+ - независимые CAM
Клетки связываются
Слайд 66Интегрины обеспечиают
СИГНАЛИЗАЦИЮ ИЗ ВНЕКЛЕТОЧНОЙ СРЕДЫ:
Внеклеточныый домен связывается с лигандом
(ММ или CAM)
?
Конформационные
Интегрины обеспечиают
СИГНАЛИЗАЦИЮ ИЗ ВНЕКЛЕТОЧНОЙ СРЕДЫ:
Внеклеточныый домен связывается с лигандом
(ММ или CAM)
?
Конформационные

Слайд 67Секвестрация рецепторов
Модификация рецепторов (фосфорилирование)
Инактивация белков участвующих в переносе (фосфатазы)
Белки- «реле»
Изменение концентрации вторичных
Секвестрация рецепторов
Модификация рецепторов (фосфорилирование)
Инактивация белков участвующих в переносе (фосфатазы)
Белки- «реле»
Изменение концентрации вторичных

 Второстепенные члены предложения. Дополнение
Второстепенные члены предложения. Дополнение Гендерные различия в формировании идеального образа мужчины и женщины у старшеклассников
Гендерные различия в формировании идеального образа мужчины и женщины у старшеклассников Дать систематизированные основы знаний о системе электропитания РЛС РТВ и первичных источниках электрической энергии
Дать систематизированные основы знаний о системе электропитания РЛС РТВ и первичных источниках электрической энергии Международные отношения дипломатия или война
Международные отношения дипломатия или война Институт патронажа в гражданском праве
Институт патронажа в гражданском праве Подготовила учитель начальных классов МОУ СОШ № 9 Романова Ирина Анатольевна
Подготовила учитель начальных классов МОУ СОШ № 9 Романова Ирина Анатольевна Дом карела. Система расселения. Планировка дома
Дом карела. Система расселения. Планировка дома А.Невский
А.Невский Tadbirda kiyinish madaniyati
Tadbirda kiyinish madaniyati ГРАЖДАНСКОЕ ОБЩЕСТВО И УЧАСТИЕ ГРАЖДАН В УПРАВЛЕНИИ
ГРАЖДАНСКОЕ ОБЩЕСТВО И УЧАСТИЕ ГРАЖДАН В УПРАВЛЕНИИ Национальное блюдо тюркоязычных народов - бешбармак
Национальное блюдо тюркоязычных народов - бешбармак Сборно-монолитный каркас. Лекция № 6
Сборно-монолитный каркас. Лекция № 6 Общий план строения стенки сосуда артерия
Общий план строения стенки сосуда артерия Виды графики. Знакомство с эстампом
Виды графики. Знакомство с эстампом Экстремальные виды спорта для людей с безграничными возможностями
Экстремальные виды спорта для людей с безграничными возможностями Эксплуатация МТ. Основные задачи. Приемка в эксплуатацию
Эксплуатация МТ. Основные задачи. Приемка в эксплуатацию Kaws 2
Kaws 2 Психологические особенности обучающихся и их влияние на эффективность учебной деятельности
Психологические особенности обучающихся и их влияние на эффективность учебной деятельности Начнём давайте изготовление аксессуаров с бус и браслетов
Начнём давайте изготовление аксессуаров с бус и браслетов Пейзаж в романе Ивана Сергеевича Тургенева «Отцы и дети»
Пейзаж в романе Ивана Сергеевича Тургенева «Отцы и дети» Илья Ефимович Репин
Илья Ефимович Репин How should someone take a phone and a cup, to use this movement of hand for rennovation of continious health
How should someone take a phone and a cup, to use this movement of hand for rennovation of continious health Как помочь ребенку готовить домашнее задание
Как помочь ребенку готовить домашнее задание Образовательный и коррекционно - развивающий процесс: ключевые компетенции педагога в инклюзивном образовании
Образовательный и коррекционно - развивающий процесс: ключевые компетенции педагога в инклюзивном образовании Викторина В мире этикеток
Викторина В мире этикеток Предложение о приобретении 100% капитала ООО НПО Наногеология
Предложение о приобретении 100% капитала ООО НПО Наногеология Презентация на тему Строение растительной клетки
Презентация на тему Строение растительной клетки Все про луну
Все про луну