- Загальна характеристика родини POLYOMAVIRIDAE

Содержание
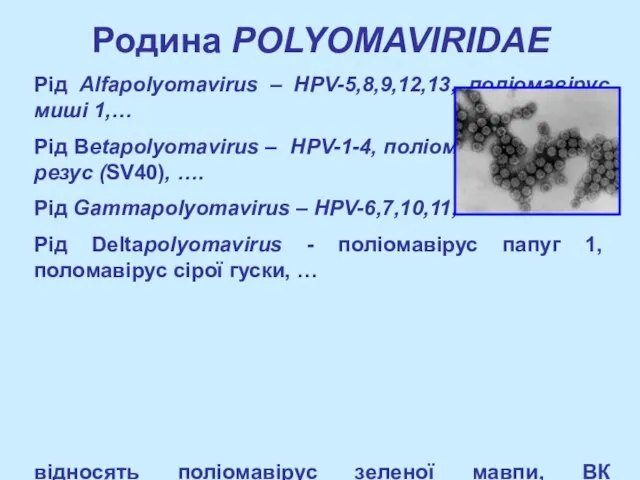
- 2. Родина POLYOMAVIRIDAE Рід Alfapolyomavirus – HPV-5,8,9,12,13, поліомавірус миші 1,… Рід Betapolyomavirus – HPV-1-4, поліомавірус макаки-резус (SV40),
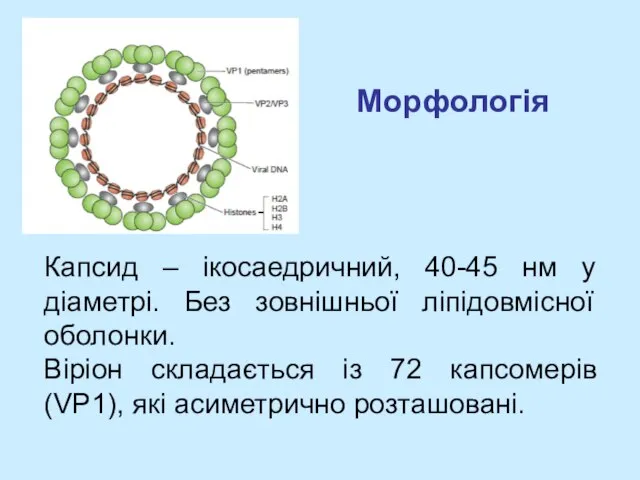
- 4. Морфологія Капсид – ікосаедричний, 40-45 нм у діаметрі. Без зовнішньої ліпідовмісної оболонки. Віріон складається із 72
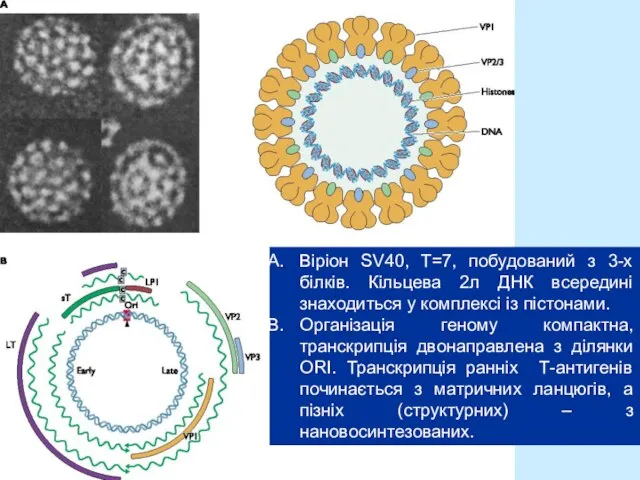
- 5. Віріон SV40, T=7, побудований з 3-х білків. Кільцева 2л ДНК всередині знаходиться у комплексі із пістонами.
- 6. Структура геному Вірусний геном може бути розділений на 3 регіони. Один з яких кодує два функціональні
- 7. Геном поліомавірусу миші
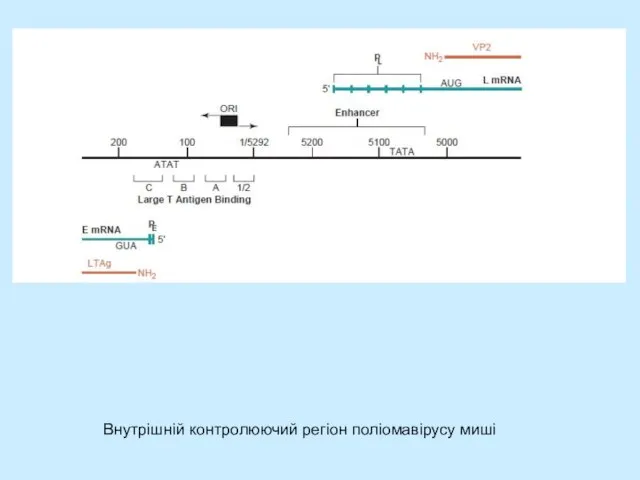
- 8. Внутрішній контролюючий регіон поліомавірусу миші
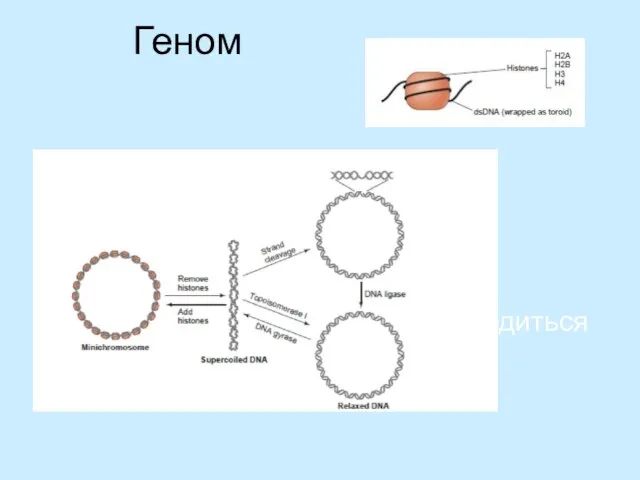
- 9. Геном Кільцева 2л ДНК всередині знаходиться у комплексі із пістонами
- 10. Функції великого Т антигену (SV40) in vitro та in vivo Зв’язування з ДНК (специфічно з ділянкою
- 11. Онкогенні властивості SV-40 та поліомавіруси миші
- 12. Ранні T, mT (у поліомавірусів миші та хомяка), t, Пізні VP1- головний компонент, що формує гексони;
- 13. Ліпіди та вуглеводи у складі віріону не виявлені.
- 14. Фізико-хімічні властивості Маса віріонів 25 x 103 кДа. Щільність у градієнті сахарози та CsCl 1,2 та
- 15. Адсорбція Вірус приєднується до поверхні чутливої клітини шляхом з’єднання VP1 з клітинним рецептором. Поліомавіруси (окрім SV
- 16. Цикл репродукції SV- 40 Проникнення віріону та звільнення серцевини (рецепторний ендоцитоз). Проникнення комплексу ДНК/нуклеосома у ядро.
- 18. Транскрипція SV40 “Використовує” клітинну РНК-полімеразу II. Ранні гени кодують T антигени, що утворюються з мРНК, які
- 20. SV40 virus
- 21. Сигнали поліаденілювання та перекриття 3-кінця Сигнали поліаденілювання та перекриття 3-кінця
- 22. Реплікація Починається через 12-15 годин після ураження клітини. Напрямок – двонаправлений з ділянки origin. Великий Т
- 23. Малий T антиген Малий Т стимулює мар-кіназну
- 24. Середній Т-антиген приєднується до с-Src та активує шлях, що стимулює клітинний метаболізм та клітинний поділ
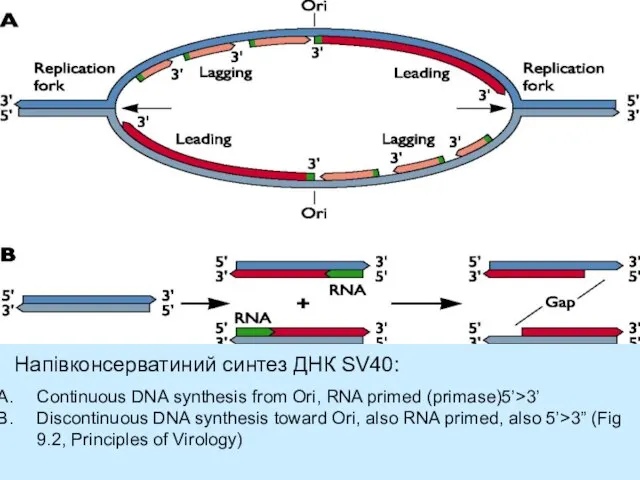
- 25. EM реплікації ДНК SV40, двонаправлена реплікація Схематичне зображення двонаправленої реплікації з єдиного сайту реплікації origin (Ori).
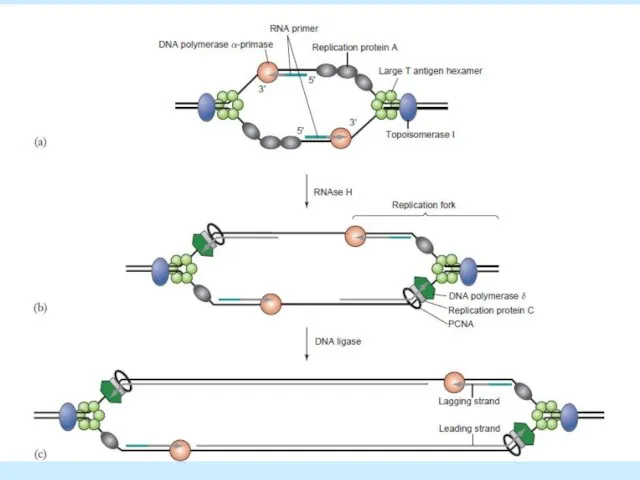
- 26. Continuous DNA synthesis from Ori, RNA primed (primase)5’>3’ Discontinuous DNA synthesis toward Ori, also RNA primed,
- 28. Реплікація SV40 має АТФ-азну активність; може функціонувати як ДНК хеліказа. Розкручує ДНК у ділянці Ori і
- 29. Пізні транскрипти Пізні мРНК: Транскрибуються після реплікації ДНК з ланцюгів, комплементарних ланцюгам, що слугують для транскрипції
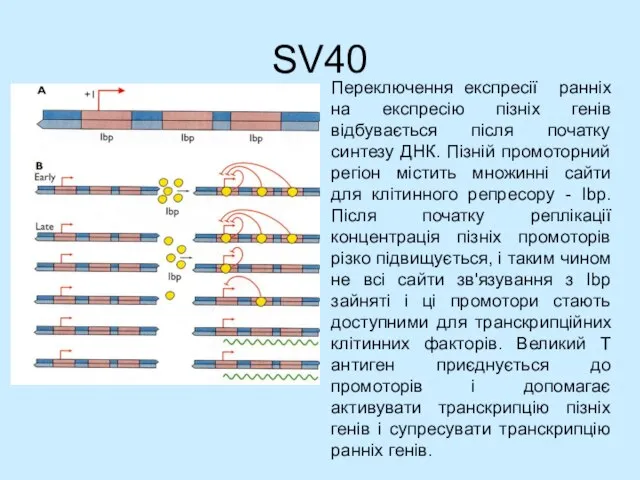
- 30. SV40 Переключення експресії ранніх на експресію пізніх генів відбувається після початку синтезу ДНК. Пізній промоторний регіон
- 31. Механізми формування структурних одиниць віріонів
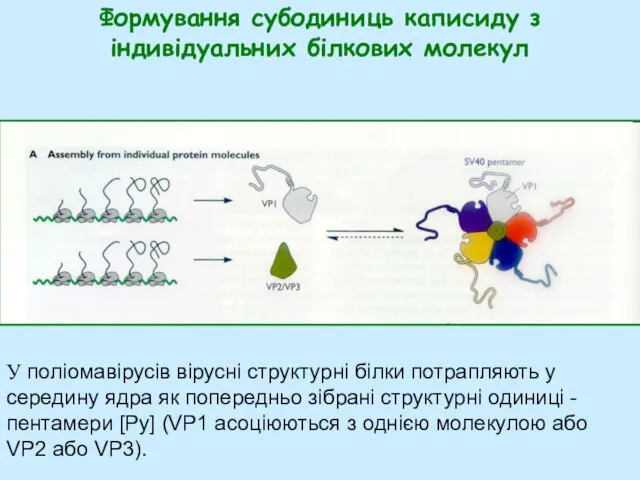
- 32. Формування субодиниць каписиду з індивідуальних білкових молекул У поліомавірусів вірусні структурні білки потрапляють у середину ядра
- 34. Формування вірусних часток та вихід з клітини Збірка: Відбувається в ядрі; Вірусний геном асоціюється з гістонами
- 35. Віруси мають чітко виражене коло хазяїв та розмножуються у певних КК. У непермісивних клітинах вірус може
- 36. Поліомавіруси розповсюджені по всьому світі. Вони часто призводять до розвитку персистентної інфекції. SV-40 є лімфотропним вірусом
- 37. Вірус JC уражує клітини дихальної системи, нирок та мозку. Вірус ВК призводить до респіраторних інфекцій, та
- 40. Механізми передачі 1. Парентеральний; 2. Контактний; 3. Повітряно-крапельний; 4. Трансплацентарний. Віруси розповсюджуються за рахунок реактивації у
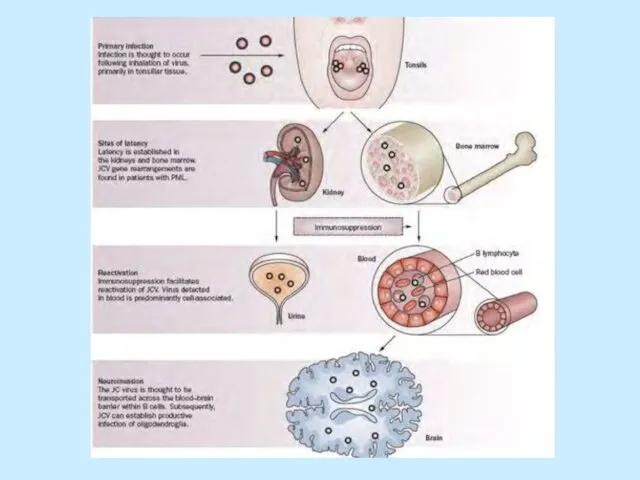
- 41. From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005, Fig.52-8. Патогенез JC/BK Проникнення
- 43. Антитіла до BKV виявляються у 90% дітей до 10 років. Нефропатія, обумовлена BKV, в більшості випадків
- 44. Помилки в діагностуванні (негативні результати) завжди призводить до незворотної нефропатії. Біопсія алотрансплантатів на наявність BKV є
- 45. JCV Людський вірус JCV може викликати руйнування олігодендріоцитів ЦНС, викликаючи при цьому фатальну демієлінізаційну хворобу, яка
- 46. JCV JCV виявляється у сечі 30-70% дорослого населення, у 90% дорослого населення виявляються антитіла до JCV.
- 47. JCV Ураження JCV відбувається в ранньому дитинстві ймовірно через респіраторний тракт. А потім розвивається персистентна інфекція
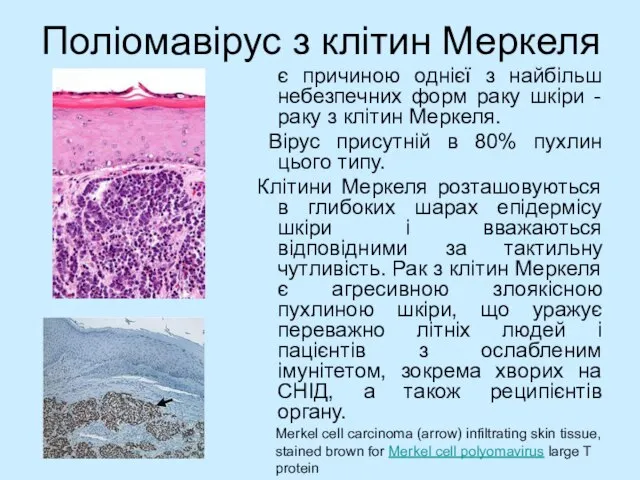
- 48. Поліомавірус з клітин Меркеля є причиною однієї з найбільш небезпечних форм раку шкіри - раку з
- 49. Діагностика Для імуногістологічної діагностики звичайно застосовують анти-SV40 антитіла, які виявляють також ВК-вірус та JC-вірус. Комерційні ДНК-зонди
- 50. Виявлення специфічних клітин у незабарвленій свіжозібраній сечі (трубчасті епітеліальні клітини з ядерними включеннями, асоційованим з BKV)
- 51. Тип I: аморфні включення типу “матове скло” вигляд ядра -“матове скло” 400x (Olympus BH2 microscope) Reference
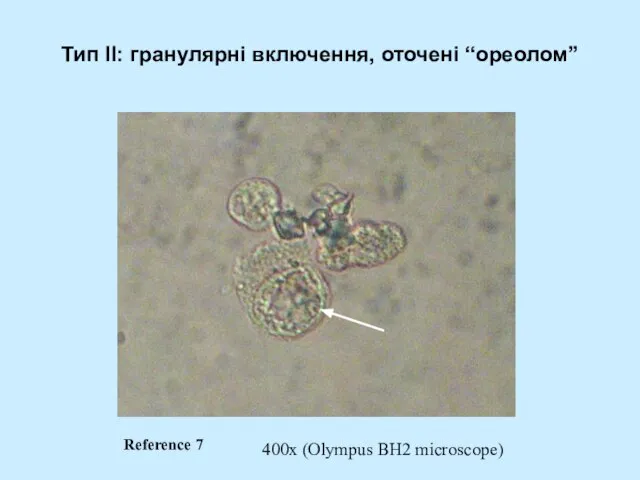
- 52. Тип II: гранулярні включення, оточені “ореолом” 400x (Olympus BH2 microscope) Reference 7
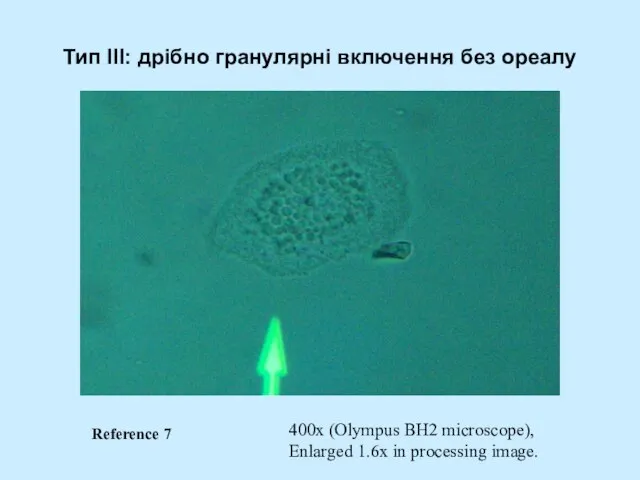
- 53. Тип III: дрібно гранулярні включення без ореалу 400x (Olympus BH2 microscope), Enlarged 1.6x in processing image.
- 54. Тип II/III гібридний: 400x (Olympus BH2 microscope), Enlarged 2x in processing image. Внутрішньоклітинні вакуолі Reference 7
- 55. Тип IV: вaкуолеподібні включення із згрупованим, невпорядкованим хроматином 400x (Olympus BH2 microscope), Enlarged 2x in processing
- 56. Альтернативним діагностичним методом є електронно-мікроскопичне дослідження негативно забарвлених осадків сечі Диференційна діагностики включає виявлення регенерованих змін
- 57. Терапія інфекції, обумовленої вірусом ВК. Основним підходом в лікуванні ВК-нефропатії є зниження імуносупресії. Можливо застосування внутрішньовенного
- 58. Екфективне лікування JCV-інфекції відсутнє. Призначають амантадин (мідантан) або цитозин, арабінозид, аденін арабінозид.
- 60. Скачать презентацию
Слайд 2Родина POLYOMAVIRIDAE
Рід Alfapolyomavirus – HPV-5,8,9,12,13, поліомавірус миші 1,…
Рід Betapolyomavirus – HPV-1-4, поліомавірус
Родина POLYOMAVIRIDAE
Рід Alfapolyomavirus – HPV-5,8,9,12,13, поліомавірус миші 1,…
Рід Betapolyomavirus – HPV-1-4, поліомавірус
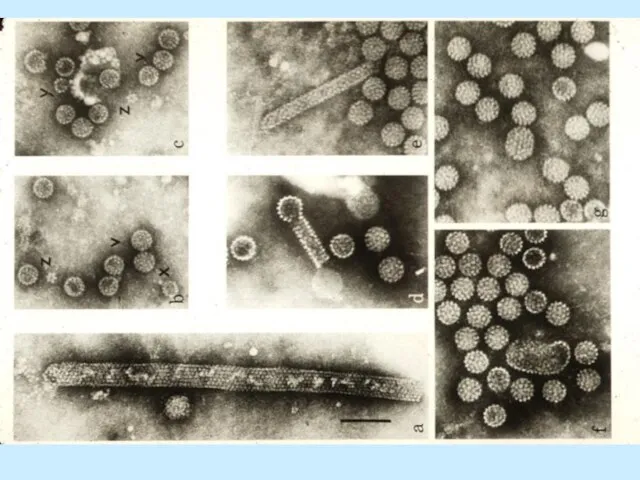
Слайд 4Морфологія
Капсид – ікосаедричний, 40-45 нм у діаметрі. Без зовнішньої ліпідовмісної оболонки.
Морфологія
Капсид – ікосаедричний, 40-45 нм у діаметрі. Без зовнішньої ліпідовмісної оболонки.
Слайд 5Віріон SV40, T=7, побудований з 3-х білків. Кільцева 2л ДНК всередині знаходиться
Віріон SV40, T=7, побудований з 3-х білків. Кільцева 2л ДНК всередині знаходиться
Слайд 6Структура геному
Вірусний геном може бути розділений на 3 регіони. Один з яких
Структура геному
Вірусний геном може бути розділений на 3 регіони. Один з яких
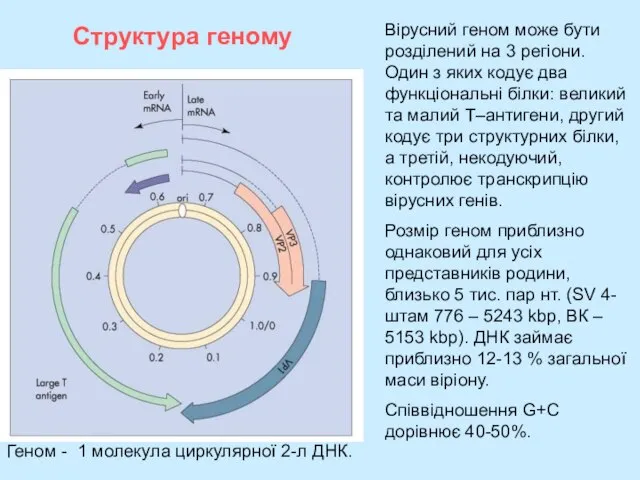
Слайд 7Геном поліомавірусу миші
Геном поліомавірусу миші
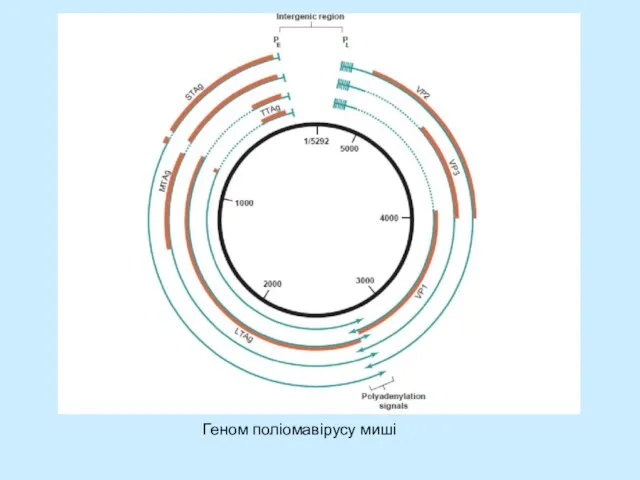
Слайд 8Внутрішній контролюючий регіон поліомавірусу миші
Внутрішній контролюючий регіон поліомавірусу миші
Слайд 9Геном
Кільцева 2л ДНК всередині знаходиться у комплексі із пістонами
Геном
Кільцева 2л ДНК всередині знаходиться у комплексі із пістонами
Слайд 10Функції великого Т антигену (SV40)
in vitro та in vivo
Зв’язування з ДНК (специфічно
Функції великого Т антигену (SV40)
in vitro та in vivo
Зв’язування з ДНК (специфічно

Слайд 11Онкогенні властивості SV-40 та поліомавіруси миші
Онкогенні властивості SV-40 та поліомавіруси миші

Слайд 12Ранні
T, mT (у поліомавірусів миші та хомяка), t,
Пізні
VP1- головний компонент,
Ранні
T, mT (у поліомавірусів миші та хомяка), t,
Пізні
VP1- головний компонент,

Слайд 13Ліпіди та вуглеводи у складі віріону не виявлені.
Ліпіди та вуглеводи у складі віріону не виявлені.

Слайд 14Фізико-хімічні властивості
Маса віріонів 25 x 103 кДа. Щільність у градієнті сахарози та
Фізико-хімічні властивості
Маса віріонів 25 x 103 кДа. Щільність у градієнті сахарози та

Слайд 15Адсорбція
Вірус приєднується до поверхні чутливої клітини шляхом з’єднання VP1 з клітинним
Адсорбція
Вірус приєднується до поверхні чутливої клітини шляхом з’єднання VP1 з клітинним

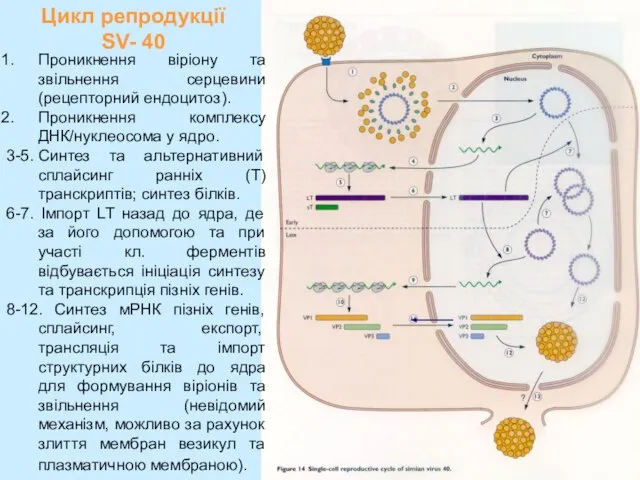
Слайд 16Цикл репродукції SV- 40
Проникнення віріону та звільнення серцевини (рецепторний ендоцитоз).
Проникнення комплексу
Цикл репродукції SV- 40
Проникнення віріону та звільнення серцевини (рецепторний ендоцитоз).
Проникнення комплексу
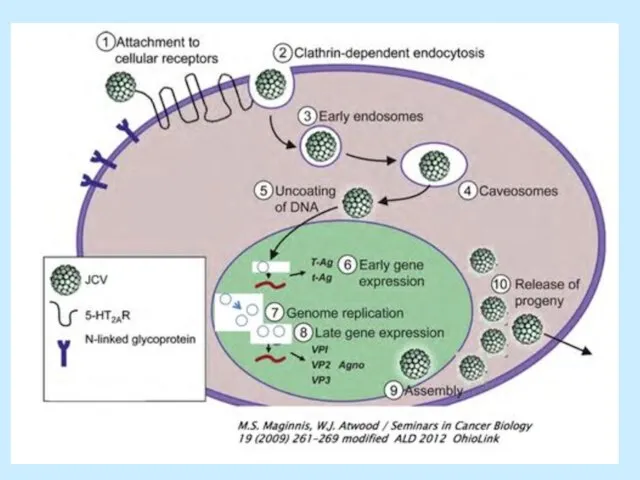
Слайд 18Транскрипція
SV40
“Використовує” клітинну РНК-полімеразу II.
Ранні гени кодують T антигени, що утворюються з
Транскрипція
SV40
“Використовує” клітинну РНК-полімеразу II.
Ранні гени кодують T антигени, що утворюються з

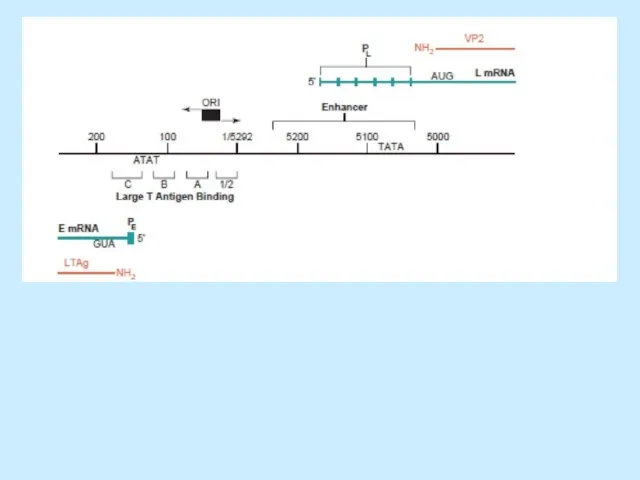
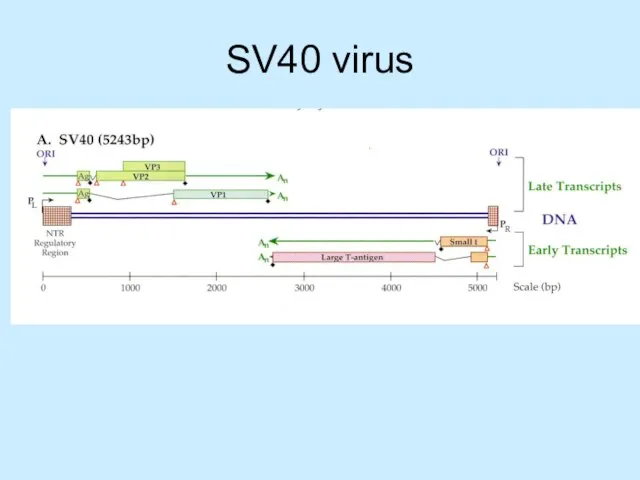
Слайд 20SV40 virus
SV40 virus
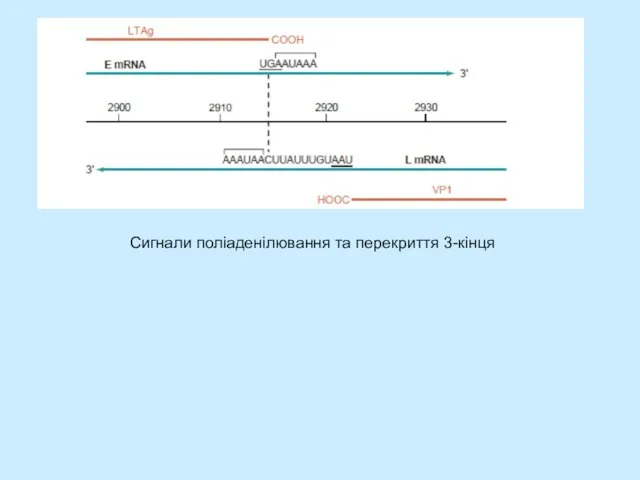
Слайд 21Сигнали поліаденілювання та перекриття 3-кінця
Сигнали поліаденілювання та перекриття 3-кінця
Сигнали поліаденілювання та перекриття 3-кінця
Сигнали поліаденілювання та перекриття 3-кінця
Слайд 22Реплікація
Починається через 12-15 годин після ураження клітини.
Напрямок – двонаправлений з ділянки
Реплікація
Починається через 12-15 годин після ураження клітини.
Напрямок – двонаправлений з ділянки

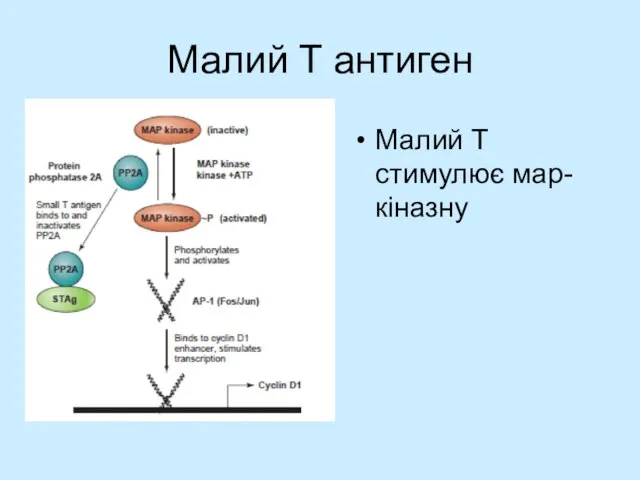
Слайд 23Малий T антиген
Малий Т стимулює мар-кіназну
Малий T антиген
Малий Т стимулює мар-кіназну
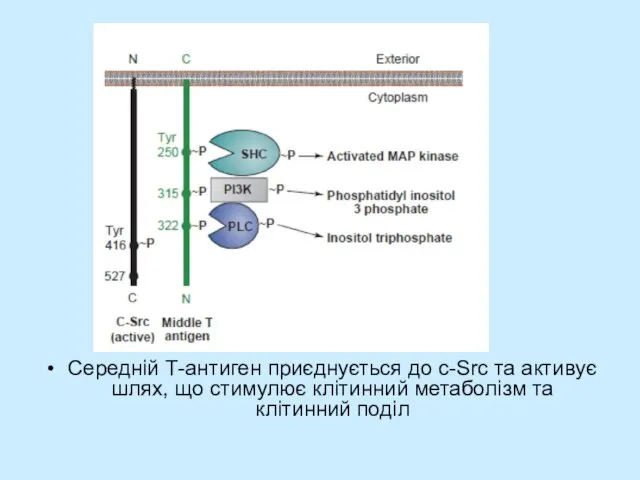
Слайд 24Середній Т-антиген приєднується до с-Src та активує шлях, що стимулює клітинний метаболізм
Середній Т-антиген приєднується до с-Src та активує шлях, що стимулює клітинний метаболізм
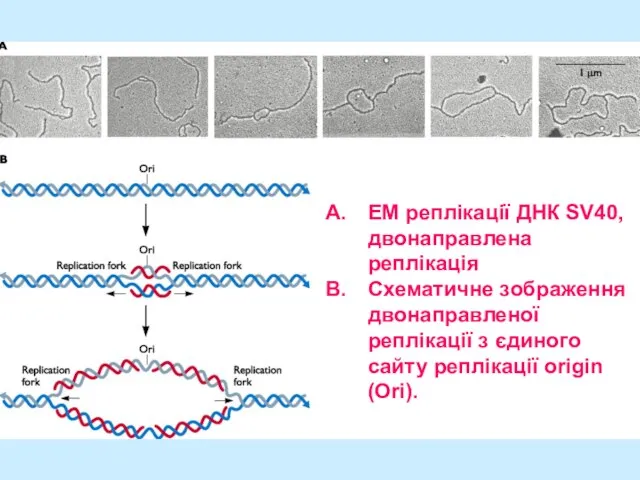
Слайд 25EM реплікації ДНК SV40, двонаправлена реплікація
Схематичне зображення двонаправленої реплікації з єдиного сайту
EM реплікації ДНК SV40, двонаправлена реплікація
Схематичне зображення двонаправленої реплікації з єдиного сайту
Слайд 26Continuous DNA synthesis from Ori, RNA primed (primase)5’>3’
Discontinuous DNA synthesis toward Ori,
Continuous DNA synthesis from Ori, RNA primed (primase)5’>3’
Discontinuous DNA synthesis toward Ori,
Слайд 28Реплікація SV40
має АТФ-азну активність;
може функціонувати як ДНК хеліказа. Розкручує ДНК у
Реплікація SV40
має АТФ-азну активність;
може функціонувати як ДНК хеліказа. Розкручує ДНК у

Слайд 29Пізні транскрипти
Пізні мРНК:
Транскрибуються після реплікації ДНК з ланцюгів, комплементарних ланцюгам, що слугують
Пізні транскрипти
Пізні мРНК:
Транскрибуються після реплікації ДНК з ланцюгів, комплементарних ланцюгам, що слугують

Слайд 30SV40
Переключення експресії ранніх на експресію пізніх генів відбувається після початку синтезу ДНК.
SV40
Переключення експресії ранніх на експресію пізніх генів відбувається після початку синтезу ДНК.
Слайд 31Механізми формування структурних одиниць віріонів
Механізми формування структурних одиниць віріонів

Слайд 32Формування субодиниць каписиду з індивідуальних білкових молекул
У поліомавірусів вірусні структурні білки потрапляють
Формування субодиниць каписиду з індивідуальних білкових молекул
У поліомавірусів вірусні структурні білки потрапляють
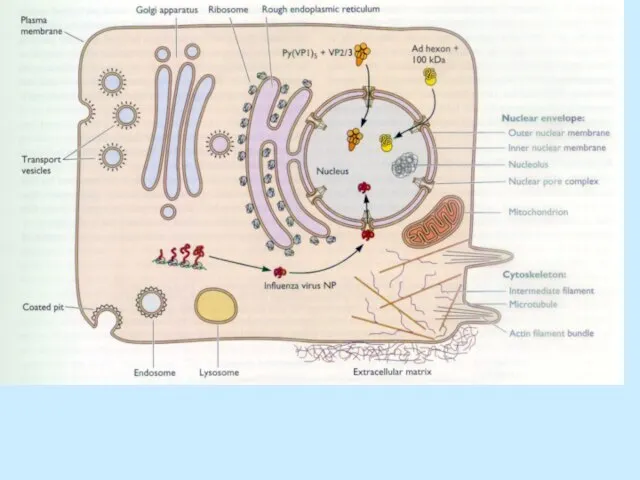
Слайд 34Формування вірусних часток та вихід з клітини
Збірка:
Відбувається в ядрі;
Вірусний геном
Формування вірусних часток та вихід з клітини
Збірка:
Відбувається в ядрі;
Вірусний геном

Слайд 35Віруси мають чітко виражене коло хазяїв та розмножуються у певних КК. У
Віруси мають чітко виражене коло хазяїв та розмножуються у певних КК. У

Слайд 36Поліомавіруси розповсюджені по всьому світі. Вони часто призводять до розвитку персистентної інфекції.
Поліомавіруси розповсюджені по всьому світі. Вони часто призводять до розвитку персистентної інфекції.

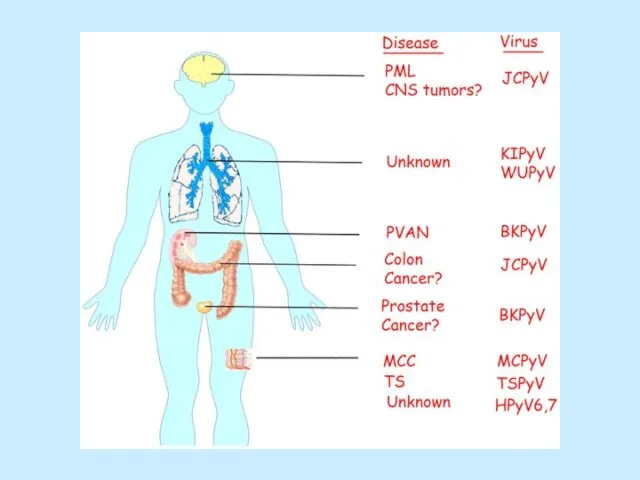
Слайд 37Вірус JC уражує клітини дихальної системи, нирок та мозку.
Вірус ВК призводить до
Вірус JC уражує клітини дихальної системи, нирок та мозку.
Вірус ВК призводить до

Слайд 40Механізми передачі
1. Парентеральний;
2. Контактний;
3. Повітряно-крапельний;
4. Трансплацентарний.
Віруси розповсюджуються
за рахунок реактивації у організмі
Механізми передачі
1. Парентеральний;
2. Контактний;
3. Повітряно-крапельний;
4. Трансплацентарний.
Віруси розповсюджуються
за рахунок реактивації у організмі

Слайд 41From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005,
From Medical Microbiology, 5th ed., Murray, Rosenthal & Pfaller, Mosby Inc., 2005,
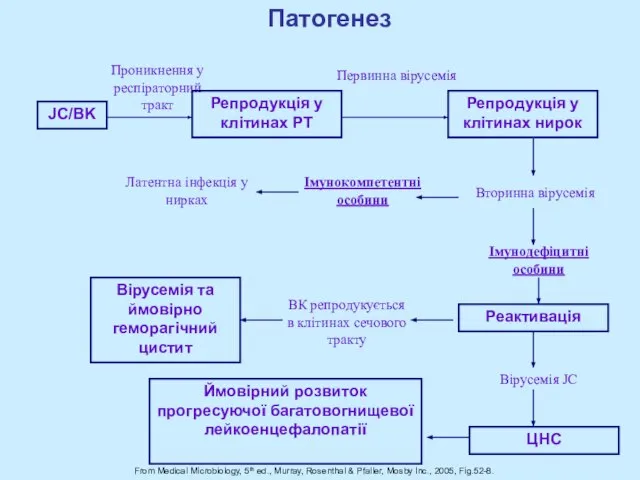
Слайд 43Антитіла до BKV виявляються у 90% дітей до 10 років.
Нефропатія, обумовлена BKV,
Антитіла до BKV виявляються у 90% дітей до 10 років.
Нефропатія, обумовлена BKV,

Слайд 44Помилки в діагностуванні (негативні результати) завжди призводить до незворотної нефропатії.
Біопсія алотрансплантатів на
Помилки в діагностуванні (негативні результати) завжди призводить до незворотної нефропатії.
Біопсія алотрансплантатів на

Слайд 45JCV
Людський вірус JCV може викликати руйнування олігодендріоцитів ЦНС, викликаючи при цьому
JCV
Людський вірус JCV може викликати руйнування олігодендріоцитів ЦНС, викликаючи при цьому

Слайд 46JCV
JCV виявляється у сечі 30-70% дорослого населення, у 90% дорослого населення
JCV
JCV виявляється у сечі 30-70% дорослого населення, у 90% дорослого населення

Слайд 47JCV
Ураження JCV відбувається в ранньому дитинстві ймовірно через респіраторний тракт. А
JCV
Ураження JCV відбувається в ранньому дитинстві ймовірно через респіраторний тракт. А

Слайд 48Поліомавірус з клітин Меркеля
є причиною однієї з найбільш небезпечних форм раку
Поліомавірус з клітин Меркеля
є причиною однієї з найбільш небезпечних форм раку
Слайд 49Діагностика
Для імуногістологічної діагностики звичайно застосовують анти-SV40 антитіла, які виявляють також ВК-вірус
Діагностика
Для імуногістологічної діагностики звичайно застосовують анти-SV40 антитіла, які виявляють також ВК-вірус

Слайд 50
Виявлення специфічних клітин у незабарвленій свіжозібраній сечі
(трубчасті епітеліальні клітини з ядерними
Виявлення специфічних клітин у незабарвленій свіжозібраній сечі (трубчасті епітеліальні клітини з ядерними

Слайд 51Тип I: аморфні включення типу “матове скло”
вигляд ядра -“матове скло”
400x (Olympus
Тип I: аморфні включення типу “матове скло”
вигляд ядра -“матове скло”
400x (Olympus

Слайд 52Тип II: гранулярні включення, оточені “ореолом”
400x (Olympus BH2 microscope)
Reference 7
Тип II: гранулярні включення, оточені “ореолом”
400x (Olympus BH2 microscope)
Reference 7
Слайд 53Тип III: дрібно гранулярні включення без ореалу
400x (Olympus BH2 microscope),
Enlarged 1.6x in
Тип III: дрібно гранулярні включення без ореалу
400x (Olympus BH2 microscope),
Enlarged 1.6x in
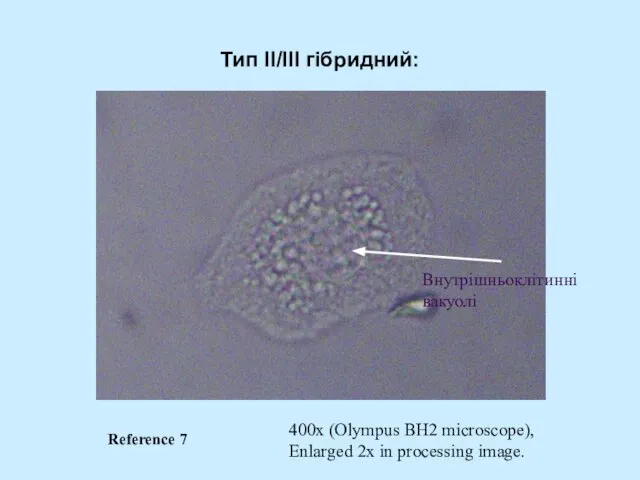
Слайд 54Тип II/III гібридний:
400x (Olympus BH2 microscope),
Enlarged 2x in processing image.
Внутрішньоклітинні
вакуолі
Reference 7
Тип II/III гібридний:
400x (Olympus BH2 microscope),
Enlarged 2x in processing image.
Внутрішньоклітинні
вакуолі
Reference 7
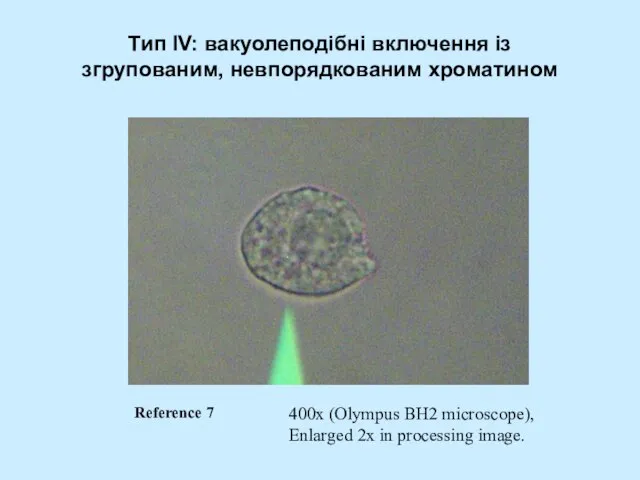
Слайд 55Тип IV: вaкуолеподібні включення із згрупованим, невпорядкованим хроматином
400x (Olympus BH2 microscope),
Enlarged 2x
Тип IV: вaкуолеподібні включення із згрупованим, невпорядкованим хроматином
400x (Olympus BH2 microscope),
Enlarged 2x
Слайд 56Альтернативним діагностичним методом є електронно-мікроскопичне дослідження негативно забарвлених осадків сечі
Диференційна діагностики
Альтернативним діагностичним методом є електронно-мікроскопичне дослідження негативно забарвлених осадків сечі
Диференційна діагностики

Слайд 57Терапія інфекції, обумовленої вірусом ВК.
Основним підходом в лікуванні ВК-нефропатії є зниження імуносупресії.
Терапія інфекції, обумовленої вірусом ВК.
Основним підходом в лікуванні ВК-нефропатії є зниження імуносупресії.

Слайд 58Екфективне лікування JCV-інфекції відсутнє. Призначають амантадин (мідантан) або цитозин, арабінозид, аденін арабінозид.
Екфективне лікування JCV-інфекції відсутнє. Призначають амантадин (мідантан) або цитозин, арабінозид, аденін арабінозид.

 ПОРЯДОК
ПОРЯДОК Место компьютера в современном мире
Место компьютера в современном мире Комплексные числа и арифметические операции над ними.
Комплексные числа и арифметические операции над ними. Горное дело
Горное дело Орнаменты ханты и священные изображения
Орнаменты ханты и священные изображения Взаимосвязь объектов в архитектурном макете
Взаимосвязь объектов в архитектурном макете Модели и типы обучения
Модели и типы обучения  Рецензия на картину Рафаэдя Санти "Сикстинская Мадонна"
Рецензия на картину Рафаэдя Санти "Сикстинская Мадонна" Бабочкина почта
Бабочкина почта Презентация на тему Зима (1 класс)
Презентация на тему Зима (1 класс) SEZUAL зрение для слепых
SEZUAL зрение для слепых ARRP и RARP Топологии Маршрутизаторы Cisco
ARRP и RARP Топологии Маршрутизаторы Cisco Предмет, методы, система экологического права как отрасли права
Предмет, методы, система экологического права как отрасли права Русская ярмарка (3 класс)
Русская ярмарка (3 класс) Твои игрушки
Твои игрушки Музей боевой славы (Иркутск)
Музей боевой славы (Иркутск) Психологический портрет Лукьянова Егора
Психологический портрет Лукьянова Егора Аккредитация ОУ. Предмет "Технология"
Аккредитация ОУ. Предмет "Технология" Статическая и динамическая сцинтиграфия
Статическая и динамическая сцинтиграфия Московский велоклуб«32 спицы»www.32spokes.ru2011 г.
Московский велоклуб«32 спицы»www.32spokes.ru2011 г. Презентация на тему Финансовая математика
Презентация на тему Финансовая математика Пушкины Зинаида Тимофеевна и Борис Борисович
Пушкины Зинаида Тимофеевна и Борис Борисович Презентация на тему Что такое светская этика
Презентация на тему Что такое светская этика  СигналТерритория Детства от Монтессори-центра
СигналТерритория Детства от Монтессори-центра НОРМАТИВНО-ПРАВОВОЕ ОБЕСПЕЧЕНИЕ ВВЕДЕНИЯ ФГОС НОО В ОУ
НОРМАТИВНО-ПРАВОВОЕ ОБЕСПЕЧЕНИЕ ВВЕДЕНИЯ ФГОС НОО В ОУ Понятие о причастии
Понятие о причастии Квиллинг для начинающих
Квиллинг для начинающих Введение в непрерывную интеграцию
Введение в непрерывную интеграцию