- Биологическое окисление

Содержание
- 2. Метаболизм представляет собой совокупность двух разнонаправленных процессов: катаболизма, анаболизма и энергетический обмен. Обмен веществ включает: поступление
- 3. Экзергонические реакции ΔG 0 1г. белков – 17кДж (4,2ккал) 1г. углеводов ̴ 17кДж (4,1ккал) 1г. жиров
- 4. Центральную роль в энергическом обмене играет АТФ Аденозинтрифосфорная кислота (АТФ). В молекуле АТФ две высокоэнергетические (макроэргические)
- 5. Энергия Гиббса гидролиза некоторых соединений Если энергия, освобождающаяся при реакциях гидролиза превышает 30 кДж/моль, то гидролизуемую
- 6. Наиболее важными экзергоническими реакциями метаболизма являются реакции окисления органических веществ, в которых используется кислород и образуется
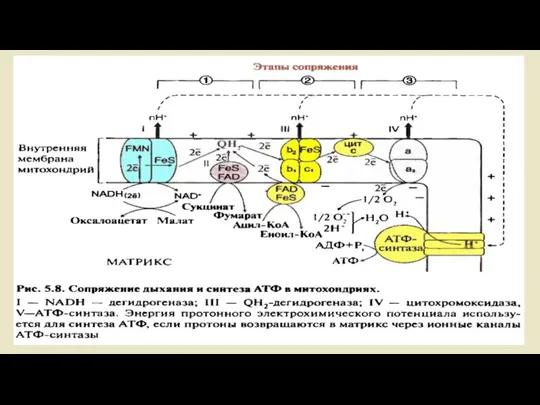
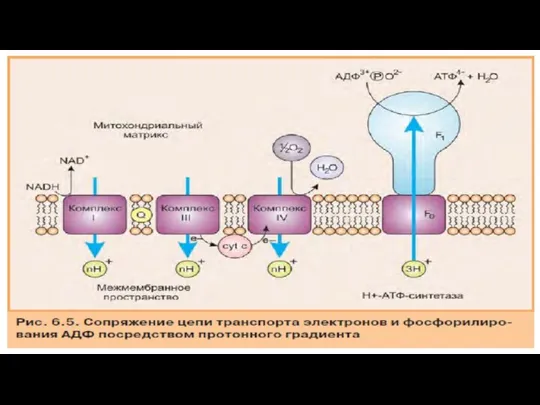
- 7. Синтез АТФ из АДФ и Н3РО4 за счет энергии, выделяющейся при тканевом дыхании, называется окислительным фосфорилированием.
- 8. Функция дыхательной цепи заключается в ситезе АТФ, которая затем может использоваться для энергетически невыгодных реакциях. Хемиосмотическая
- 10. Дыхательная цепь как цепь переноса электронов -Химическая реакция, лежащая в основе образования воды в дыхательной цепи,
- 13. Комплексы дыхательной цепи Комплекс 1: НАДН-убихинон-редуктаза ln комплекс I происходит перенос электронов от NADH + H
- 15. Кластер железасерный встречаются в разных вариантах. В качестве простетической группы, они координированы с различными остатками цистеинабелка.
- 16. Комплекс II: Сукцинат-убихинон-редуктаза Комплекс II дыхательной цепи одновременно является ферментом цитратного цикла, а именно сукцинатдегидрогеназой. Это
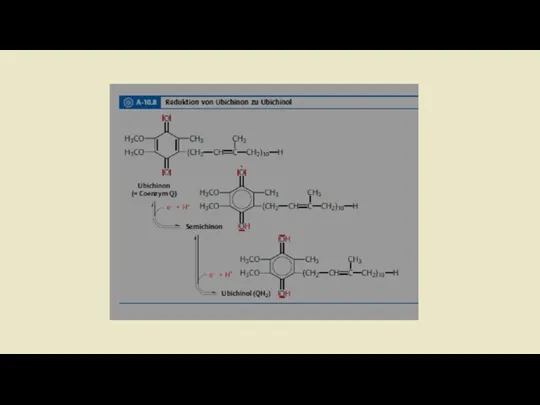
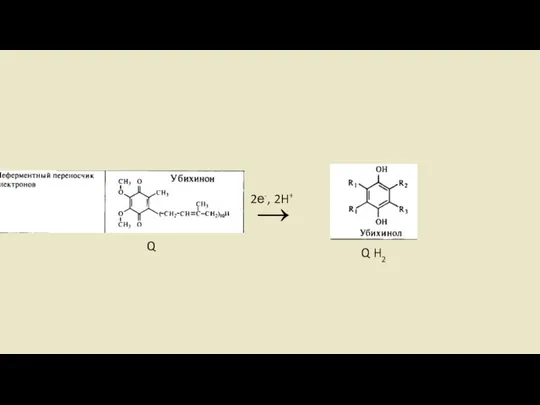
- 18. Ubichinon-Ubichino Системы Убихинон или кофермент Q состоит из одного хинона и одной изопреновая боковой цепи. Структурно
- 20. Комплекс III: Убихинол-цитохром с-редуктаза Задача комплекса 3 теперь заключается в передаче электронов убихинола на цитохром С.
- 22. Цитохромы Цитохромы - это гемсодержащие белки. В видимом свете спектра поглощения востановительные группы гема имеют три
- 23. Комплекс IV: цитохром-с-оксидаза Задача комплекса IV заключается в переносе электронов с цитохрома с на молекулярный кислород.
- 25. Комплекс V: АТФ-синтаза Комплексы I. lll и IV способствуют созданию протонного градиента через внутреннюю мембрану митохондрий.
- 26. Строение и механизм действия АТФ - синтетазы. А – F0 и F1 – комплексы АТФ –
- 28. Коэффициент фосфорилирования P/O Коэффициент Р/О – это отношение количества неорганического фосфата, включенного в молекулу АТФ АТФ-синтазой,
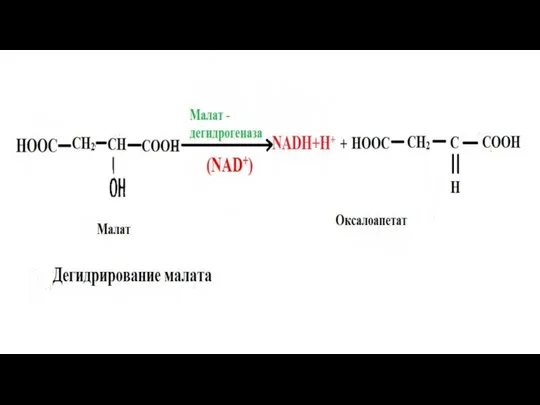
- 31. Большинство дегидрогеназ, поставляющих электроны в межмембранные пространства содержат НАД. Они катализируют реакции типа R – CHOH
- 32. В матриксе митохондрии идут реакции окисления субстратов (метаболитов распада углеводов, липидов, глутаминовой кислоты) путем дегидрирования, т.
- 36. Строение железо-серных центров Ι - FeS - центр; атом железа связан координационными связями с четырьмя атомами
- 37. → 2е-, 2H+ Q Q H2
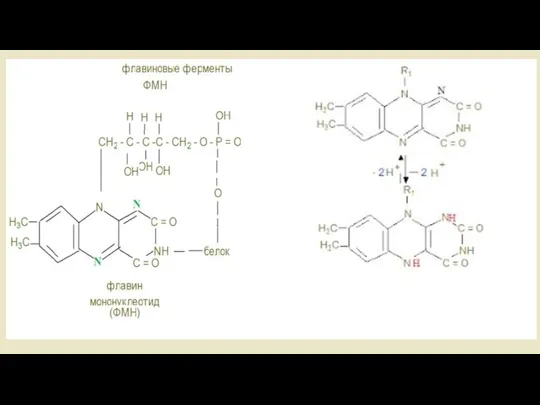
- 38. Витамин В2 - рабочая часть коферментов ФМН и ФАД, отщепляющая водород от –СН2- СН2- в отличие
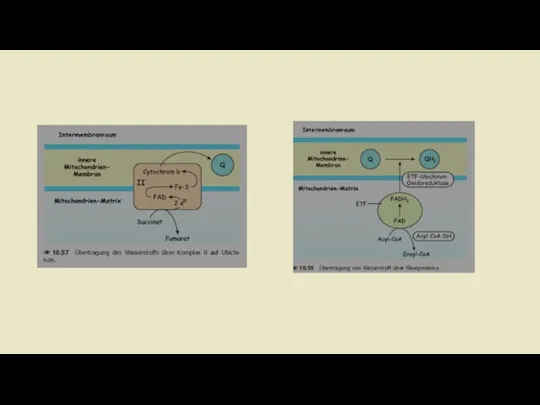
- 41. На внутренней мембране митохондрии существует система переносчиков, их действие векторно. Они переносят протоны и электроны с
- 43. Протоны перенесенные в межмембранное пространство создают протонный градиент подкисляя его 1,4 рН. Кроме того, появляется разность
- 45. Механизм разобщения дыхания и фосфорилирования. Протонированная форма 2,4 – динитрофенола переносит протоны через внутреннюю мембрану митохондрий
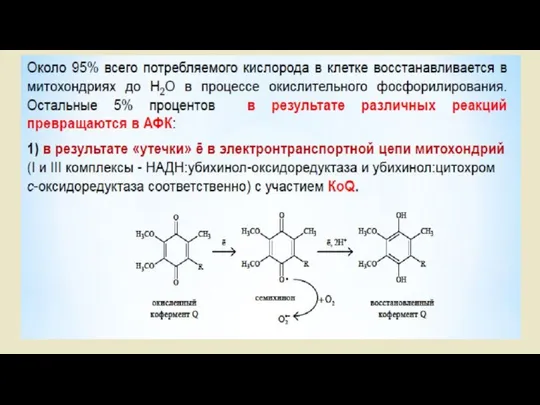
- 46. Активные формы кислорода и антиоксидантные системы. Проблема свободных радикалов в последнее десятилетие произвела настоящую революцию в
- 47. В эритроцитах окисление иона железа гемоглобина способствует образованию супероксидоного аниона. Hb (Fe2+) + O2 →MtHb (Fe+3)
- 48. Примеры свободных радикалов.
- 49. 2. ионы – гипохлорит -ион ClO4 – сопряженное основание для хлорноватистой кислоты, активный компонент отбеливателей. 3.
- 50. Образование токсичных форм при постепенном восстановлении кислорода.
- 53. Свободно-радикальное окисление нарушает структуру нуклеиновых кислот, белков и, особенно, непредельные жирные кислоты (липиды), принимающие электрон от
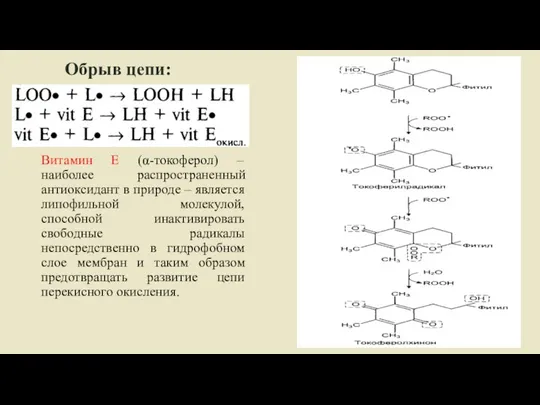
- 54. Обрыв цепи: Витамин Е (α-токоферол) – наиболее распространенный антиоксидант в природе – является липофильной молекулой, способной
- 55. В эритроцитах:
- 56. Трипептид глутатион (γ-глутамилцистеинилглицин)
- 57. Кроме витамина Е антиоксидантами являются: витамин С, β−каротин. Во-первых, витамин С восстанавливает окисленную форму витамина Е.
- 61. Скачать презентацию
Слайд 2Метаболизм представляет собой совокупность двух разнонаправленных процессов: катаболизма, анаболизма и энергетический обмен.
Обмен
Метаболизм представляет собой совокупность двух разнонаправленных процессов: катаболизма, анаболизма и энергетический обмен.
Обмен

Слайд 3Экзергонические реакции ΔG<0 Эндергонические реакции ΔG>0
1г. белков – 17кДж (4,2ккал)
1г. углеводов ̴
Экзергонические реакции ΔG<0 Эндергонические реакции ΔG>0
1г. белков – 17кДж (4,2ккал)
1г. углеводов ̴

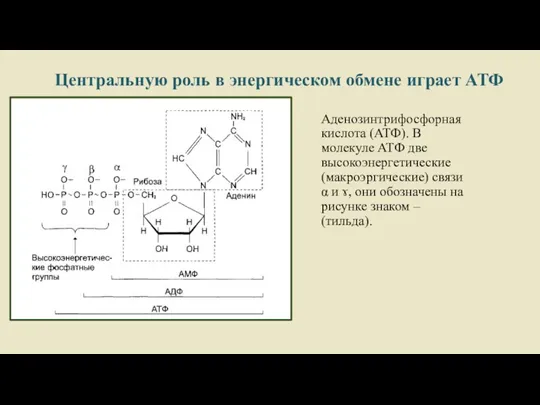
Слайд 4Центральную роль в энергическом обмене играет АТФ
Аденозинтрифосфорная кислота (АТФ). В молекуле АТФ
Центральную роль в энергическом обмене играет АТФ
Аденозинтрифосфорная кислота (АТФ). В молекуле АТФ
Слайд 5Энергия Гиббса гидролиза некоторых соединений
Если энергия, освобождающаяся при реакциях гидролиза превышает
Энергия Гиббса гидролиза некоторых соединений
Если энергия, освобождающаяся при реакциях гидролиза превышает

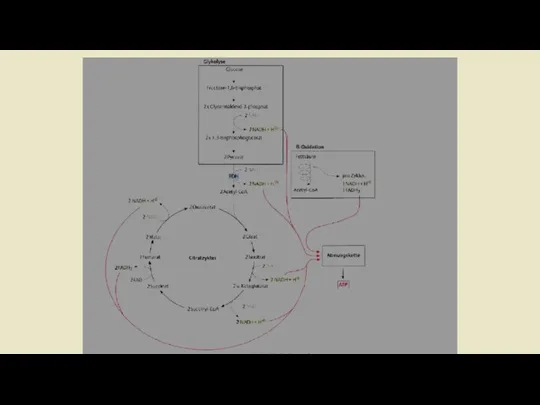
Слайд 6 Наиболее важными экзергоническими реакциями метаболизма являются реакции окисления органических веществ, в
Наиболее важными экзергоническими реакциями метаболизма являются реакции окисления органических веществ, в

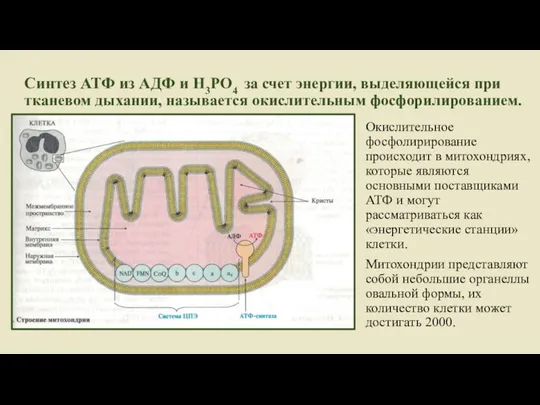
Слайд 7Синтез АТФ из АДФ и Н3РО4 за счет энергии, выделяющейся при тканевом
Синтез АТФ из АДФ и Н3РО4 за счет энергии, выделяющейся при тканевом
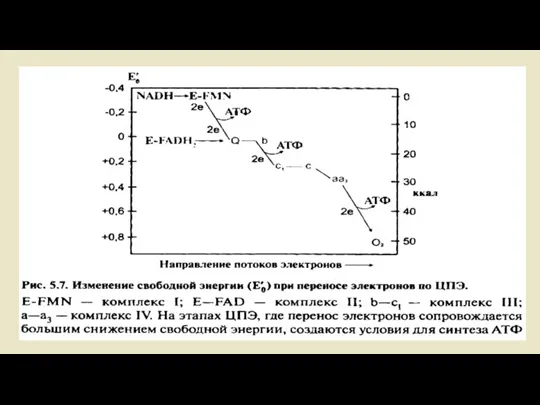
Слайд 8Функция дыхательной цепи заключается в ситезе АТФ, которая затем может использоваться для
Функция дыхательной цепи заключается в ситезе АТФ, которая затем может использоваться для

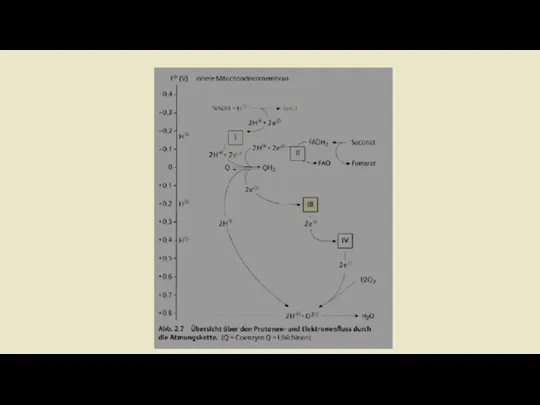
Слайд 10Дыхательная цепь как цепь переноса электронов
-Химическая реакция, лежащая в основе образования воды
Дыхательная цепь как цепь переноса электронов
-Химическая реакция, лежащая в основе образования воды

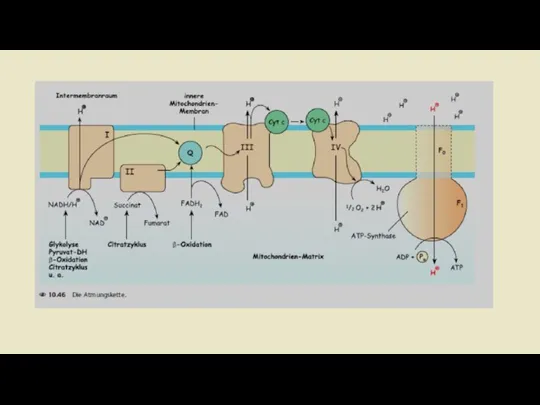
Слайд 13Комплексы дыхательной цепи
Комплекс 1: НАДН-убихинон-редуктаза
ln комплекс I происходит перенос электронов от NADH
Комплексы дыхательной цепи
Комплекс 1: НАДН-убихинон-редуктаза
ln комплекс I происходит перенос электронов от NADH

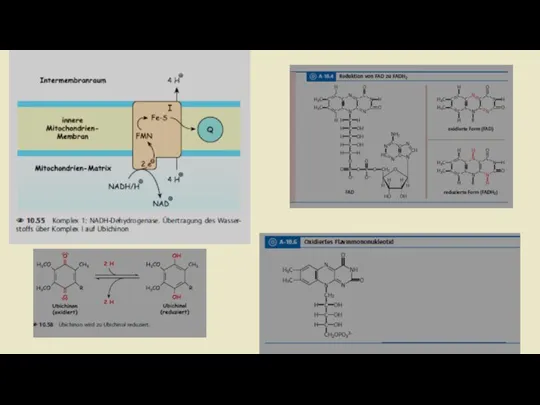
Слайд 15Кластер железасерный встречаются в разных вариантах. В качестве простетической группы, они координированы
Кластер железасерный встречаются в разных вариантах. В качестве простетической группы, они координированы

Слайд 16Комплекс II: Сукцинат-убихинон-редуктаза
Комплекс II дыхательной цепи одновременно является ферментом цитратного цикла, а
Комплекс II: Сукцинат-убихинон-редуктаза
Комплекс II дыхательной цепи одновременно является ферментом цитратного цикла, а

Слайд 18Ubichinon-Ubichino Системы
Убихинон или кофермент Q состоит из одного хинона и одной изопреновая
Ubichinon-Ubichino Системы
Убихинон или кофермент Q состоит из одного хинона и одной изопреновая

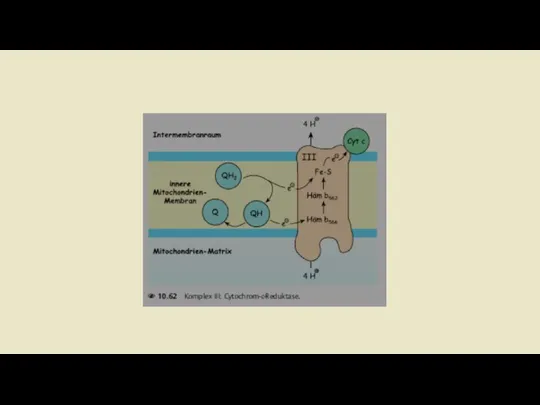
Слайд 20Комплекс III: Убихинол-цитохром с-редуктаза
Задача комплекса 3 теперь заключается в передаче электронов убихинола
Комплекс III: Убихинол-цитохром с-редуктаза
Задача комплекса 3 теперь заключается в передаче электронов убихинола

Слайд 22Цитохромы
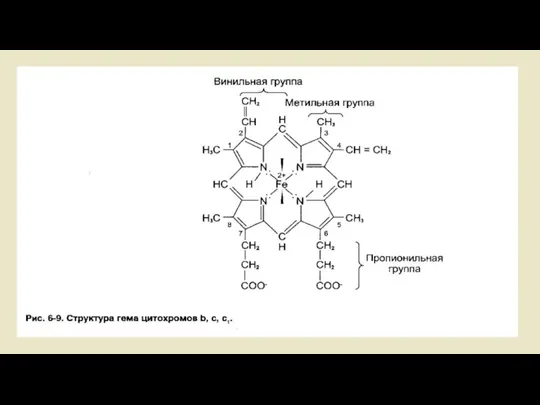
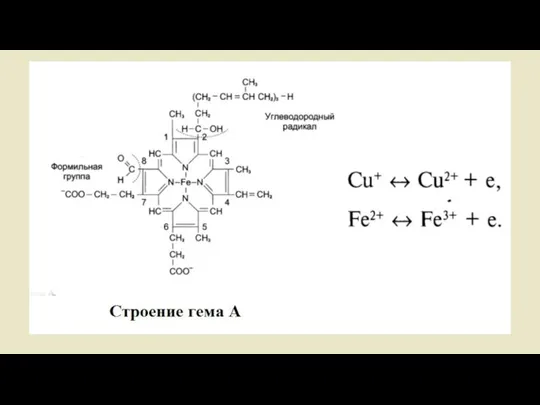
Цитохромы - это гемсодержащие белки. В видимом свете спектра поглощения востановительные группы
Цитохромы
Цитохромы - это гемсодержащие белки. В видимом свете спектра поглощения востановительные группы

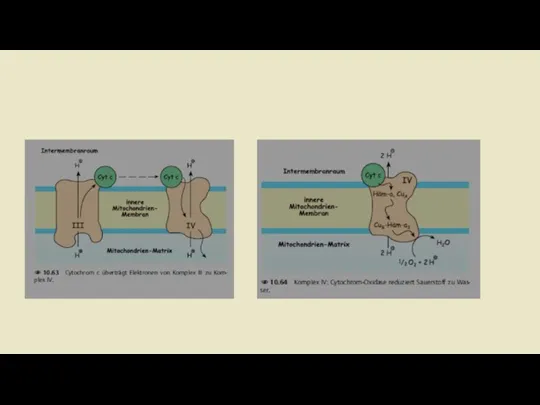
Слайд 23Комплекс IV: цитохром-с-оксидаза
Задача комплекса IV заключается в переносе электронов с цитохрома с
Комплекс IV: цитохром-с-оксидаза
Задача комплекса IV заключается в переносе электронов с цитохрома с

Слайд 25Комплекс V: АТФ-синтаза
Комплексы I. lll и IV способствуют созданию протонного градиента через
Комплекс V: АТФ-синтаза
Комплексы I. lll и IV способствуют созданию протонного градиента через

Слайд 26Строение и механизм действия
АТФ - синтетазы.
А – F0 и F1
Строение и механизм действия
АТФ - синтетазы.
А – F0 и F1


Слайд 28Коэффициент фосфорилирования P/O
Коэффициент Р/О – это отношение количества неорганического фосфата, включенного в молекулу АТФ АТФ-синтазой,
Коэффициент фосфорилирования P/O Коэффициент Р/О – это отношение количества неорганического фосфата, включенного в молекулу АТФ АТФ-синтазой,


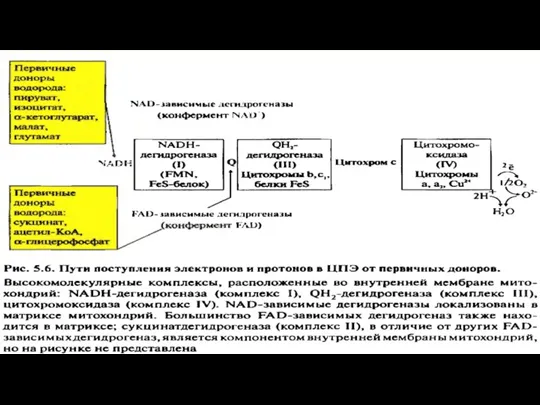
Слайд 31 Большинство дегидрогеназ, поставляющих электроны в межмембранные пространства содержат НАД. Они катализируют
Большинство дегидрогеназ, поставляющих электроны в межмембранные пространства содержат НАД. Они катализируют

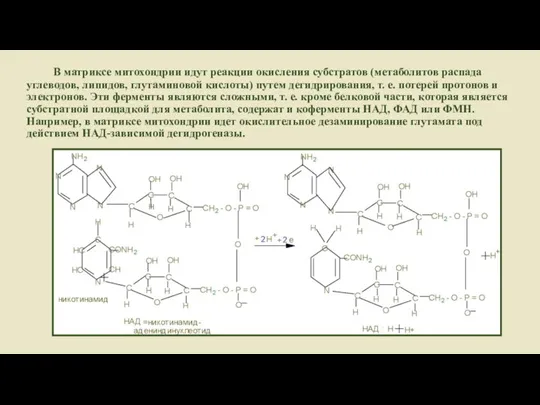
Слайд 32 В матриксе митохондрии идут реакции окисления субстратов (метаболитов распада углеводов, липидов,
В матриксе митохондрии идут реакции окисления субстратов (метаболитов распада углеводов, липидов,
Слайд 36Строение железо-серных центров
Ι - FeS - центр; атом железа связан координационными связями
Строение железо-серных центров
Ι - FeS - центр; атом железа связан координационными связями

Слайд 37 →
2е-, 2H+
Q
Q H2
→
2е-, 2H+
Q
Q H2
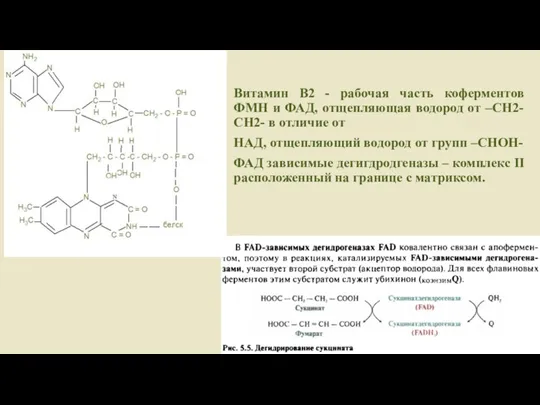
Слайд 38Витамин В2 - рабочая часть коферментов ФМН и ФАД, отщепляющая водород от
Витамин В2 - рабочая часть коферментов ФМН и ФАД, отщепляющая водород от
Слайд 41 На внутренней мембране митохондрии существует система переносчиков, их действие векторно. Они
На внутренней мембране митохондрии существует система переносчиков, их действие векторно. Они

Слайд 43 Протоны перенесенные в межмембранное пространство создают протонный градиент подкисляя его 1,4
Протоны перенесенные в межмембранное пространство создают протонный градиент подкисляя его 1,4

Слайд 45Механизм разобщения дыхания и фосфорилирования.
Протонированная форма 2,4 – динитрофенола переносит протоны
Механизм разобщения дыхания и фосфорилирования.
Протонированная форма 2,4 – динитрофенола переносит протоны

Слайд 46Активные формы кислорода и антиоксидантные системы.
Проблема свободных радикалов в последнее десятилетие
Проблема свободных радикалов в последнее десятилетие

Слайд 47В эритроцитах окисление иона железа гемоглобина способствует образованию супероксидоного аниона. Hb (Fe2+)
В эритроцитах окисление иона железа гемоглобина способствует образованию супероксидоного аниона. Hb (Fe2+)

Слайд 48Примеры свободных радикалов.
Примеры свободных радикалов.

Слайд 492. ионы – гипохлорит -ион ClO4 – сопряженное основание для хлорноватистой кислоты,
2. ионы – гипохлорит -ион ClO4 – сопряженное основание для хлорноватистой кислоты,

Слайд 50Образование токсичных форм при постепенном восстановлении кислорода.
Образование токсичных форм при постепенном восстановлении кислорода.

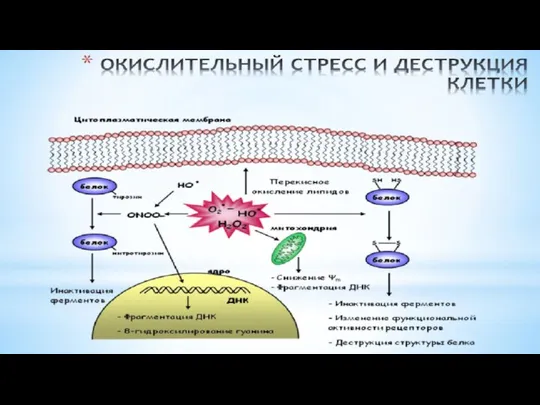
Слайд 53 Свободно-радикальное окисление нарушает структуру нуклеиновых кислот, белков и, особенно, непредельные жирные
Свободно-радикальное окисление нарушает структуру нуклеиновых кислот, белков и, особенно, непредельные жирные

Слайд 54Обрыв цепи:
Витамин Е (α-токоферол) – наиболее распространенный антиоксидант в природе – является
Обрыв цепи:
Витамин Е (α-токоферол) – наиболее распространенный антиоксидант в природе – является
Слайд 55В эритроцитах:
В эритроцитах:

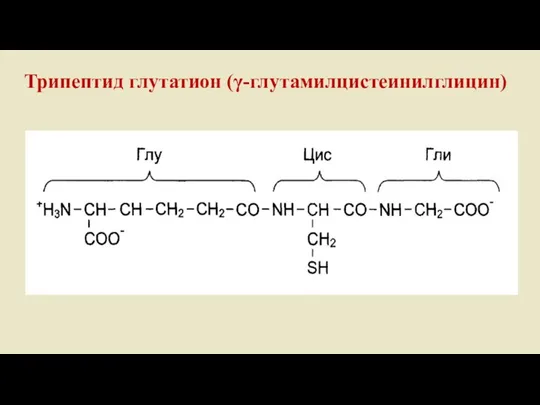
Слайд 56Трипептид глутатион (γ-глутамилцистеинилглицин)
Трипептид глутатион (γ-глутамилцистеинилглицин)
Слайд 57Кроме витамина Е антиоксидантами являются: витамин С, β−каротин.
Во-первых, витамин С восстанавливает окисленную
Кроме витамина Е антиоксидантами являются: витамин С, β−каротин. Во-первых, витамин С восстанавливает окисленную


 Земноводные Тюменской области
Земноводные Тюменской области Энергетический обмен
Энергетический обмен Движение крови по сосудам
Движение крови по сосудам Экологические факторы и их влияние на организм
Экологические факторы и их влияние на организм Сравнительная характеристика типов червей
Сравнительная характеристика типов червей Založení neziskové organizace
Založení neziskové organizace Тип Членистоногие
Тип Членистоногие Применения информационных технологий в изучении влияния фитогормонов на рост орхидей
Применения информационных технологий в изучении влияния фитогормонов на рост орхидей Клык нижней челюсти
Клык нижней челюсти Химический состав клетки
Химический состав клетки Жизнь на Земле
Жизнь на Земле Паращитоподібні, надниркові, статеві залози, підшлункова і тимус
Паращитоподібні, надниркові, статеві залози, підшлункова і тимус 5dc9ea4996b8de47
5dc9ea4996b8de47 Биосинтез белка: Транскрипция. Процессинг мРНК. Трансляция. Фолдинг
Биосинтез белка: Транскрипция. Процессинг мРНК. Трансляция. Фолдинг Ткани человека. 3
Ткани человека. 3 Отличие одного вида растений от другого на основании морфологического критерия
Отличие одного вида растений от другого на основании морфологического критерия Птицы. Внешнее строение
Птицы. Внешнее строение Жүйкелік және гуморальдік реттелудің механизмі
Жүйкелік және гуморальдік реттелудің механизмі Пищеварение полостное, внутриклеточное
Пищеварение полостное, внутриклеточное Изменение живых организмов в зависимости от среды обитания
Изменение живых организмов в зависимости от среды обитания Оценка перспективных селекционных линий ярового ячменя в условиях Западной Сибири
Оценка перспективных селекционных линий ярового ячменя в условиях Западной Сибири Лекарственные растения
Лекарственные растения Фізіологія серця, судин, гемодинаміка
Фізіологія серця, судин, гемодинаміка Занимательная биология
Занимательная биология Формы естественного отбора. Задание
Формы естественного отбора. Задание Назовите признаки жизни. 9 класс
Назовите признаки жизни. 9 класс Презентация на тему Пути достижения биологического прогресса
Презентация на тему Пути достижения биологического прогресса  Закономерности происхождения и развития жизни на Земле. 9 класс
Закономерности происхождения и развития жизни на Земле. 9 класс