Двигательные рефлексы спинного мозга. Локомоция: центры конечностей, тонические и фазические управляющие влияния
- Двигательные рефлексы спинного мозга. Локомоция: центры конечностей, тонические и фазические управляющие влияния

Содержание
- 2. Внимание! 16 мая состоится тестирование по лекциям 11-14 по особому расписанию: Группы 1-2 – с 9-00
- 3. произвольные: новые движения в новых условиях; управляет кора больших полушарий, используя сенсорный (прежде всего, зрительный) контроль;
- 4. Рефлекторные движения: Возникают в ответ на стимул (врожденно заданный или ставший значимым в результате обучения). В
- 5. Серое в-во спинного мозга: задние рога (1), боковые рога (2), передние рога (3), промежуточное ядро (4).
- 6. Пример: болевой стимул запускает сгибание конечнос-ти и вегетативные реакции; но тормозные влияния головного мозга способны задержать
- 7. Пример: болевой стимул запускает сгибание конечнос-ти и вегетативные реакции; но тормозные влияния головного мозга способны задержать
- 8. Характерные свойства сгибательного рефлекса: рефлекс стабилен и видоспецифичен (как и все врожденные рефлексы); запуск – болевой
- 9. Коленный рефлекс – частный случай миотатического рефлекса (рефлекса на растяжение мышцы): мышца (в данной случае 4-хглавая
- 10. Бицепс: сегменты C5-C6 Трицепс: C6-C8 4-хглавая мышца бедра: L2-L4
- 11. Еще примеры двигательных рефлексов: рефлекс захвата пальца (на прикосновение к ладони); рефлекс Бабинского (разгибание пальцев стопы
- 12. Л о к о м о ц и я : ритмически повторяющиеся движения (прежде всего, сгибания
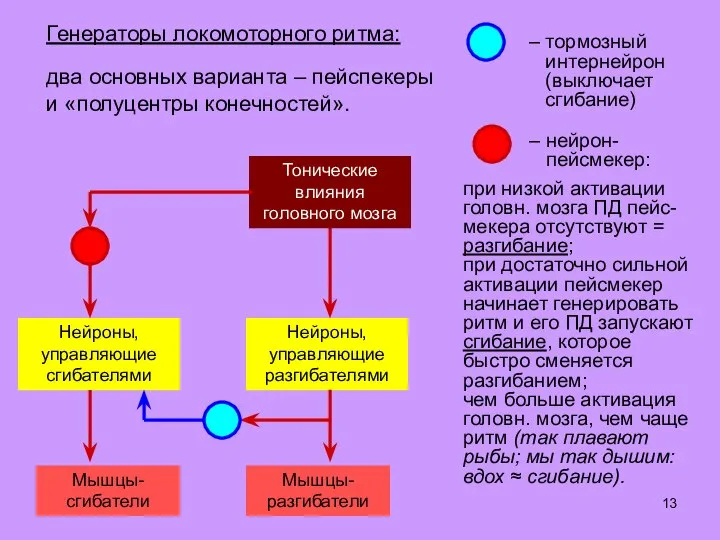
- 13. Генераторы локомоторного ритма: два основных варианта – пейспекеры и «полуцентры конечностей».
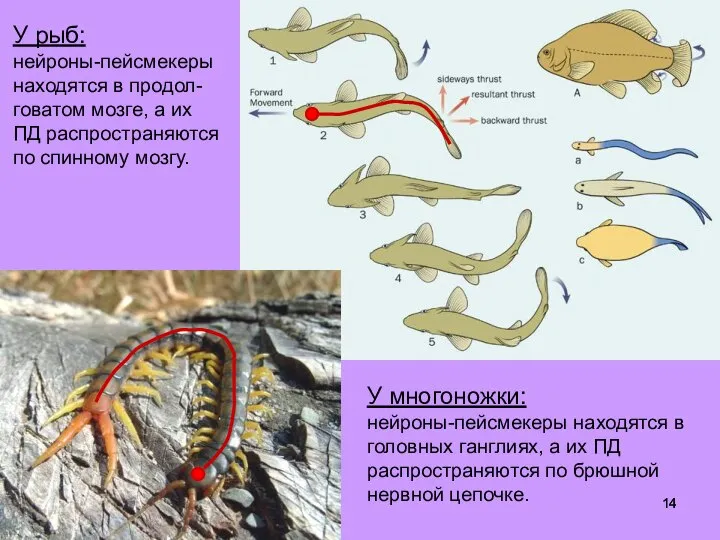
- 14. У рыб: нейроны-пейсмекеры находятся в продол-говатом мозге, а их ПД распространяются по спинному мозгу. У многоножки:
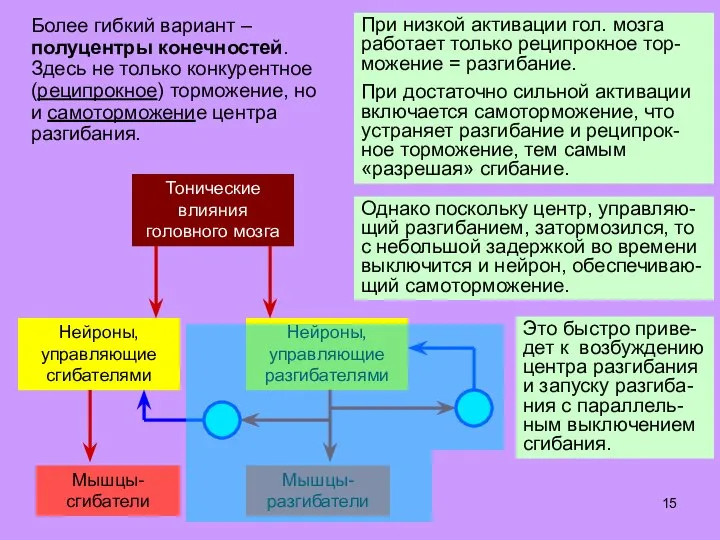
- 15. Более гибкий вариант – полуцентры конечностей. Здесь не только конкурентное (реципрокное) торможение, но и самоторможение центра
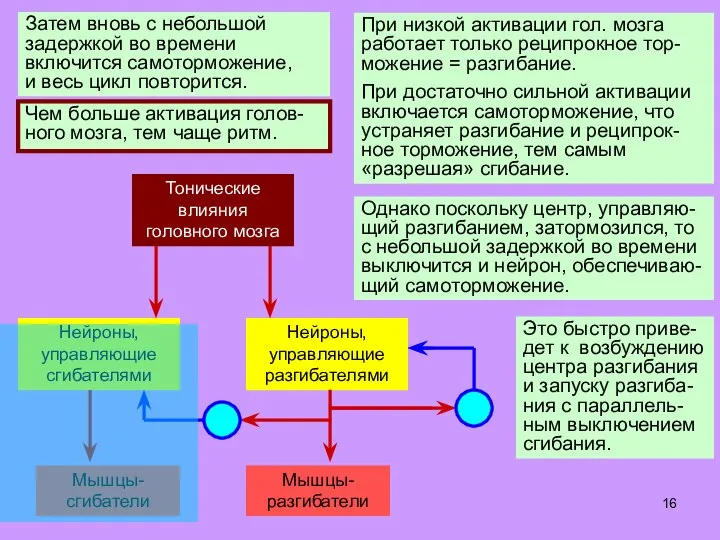
- 16. При низкой активации гол. мозга работает только реципрокное тор-можение = разгибание. При достаточно сильной активации включается
- 17. Запуск локомоции – за счет тонических влияний головного мозга. Траекторию «восьмерки» создают цепи интернейронов спинного мозга,
- 18. Запуск локомоции – за счет тонических влияний головного мозга. Траекторию «восьмерки» создают цепи интернейронов спинного мозга,
- 19. РЫСЬ Максимальная выраженность тонических влияний приводит к переключению на самый быст-рый аллюр – галоп (две передние
- 20. Мышечная чувстви-тельность Все три варианта циркуляции возбуждения при локомоции (шаг, рысь / иноходь и галоп) врожденно
- 21. Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него частично заходят красное ядро
- 22. Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него частично заходят красное ядро
- 23. Эти тракты – составные части экстрапира- мидной системы.
- 24. Произвольные движения: новые движения в новых условиях; управляет кора больших полушарий, используя сенсорный (зрительный, тактильный и
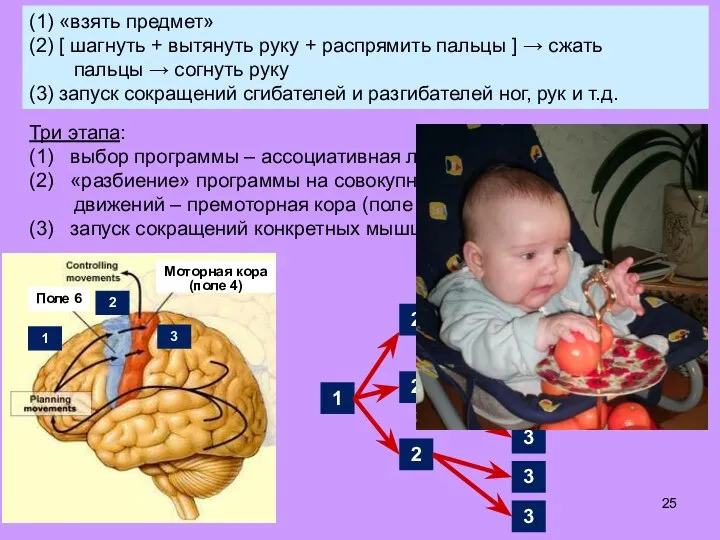
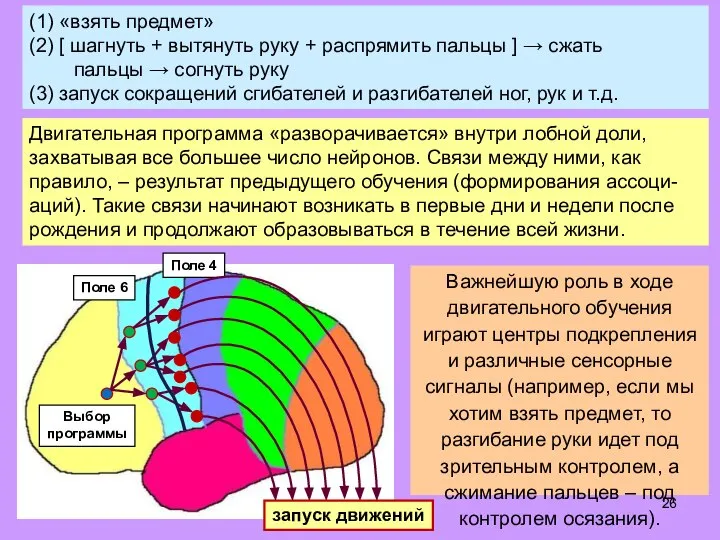
- 25. (1) «взять предмет» (2) [ шагнуть + вытянуть руку + распрямить пальцы ] → сжать пальцы
- 26. (1) «взять предмет» (2) [ шагнуть + вытянуть руку + распрямить пальцы ] → сжать пальцы
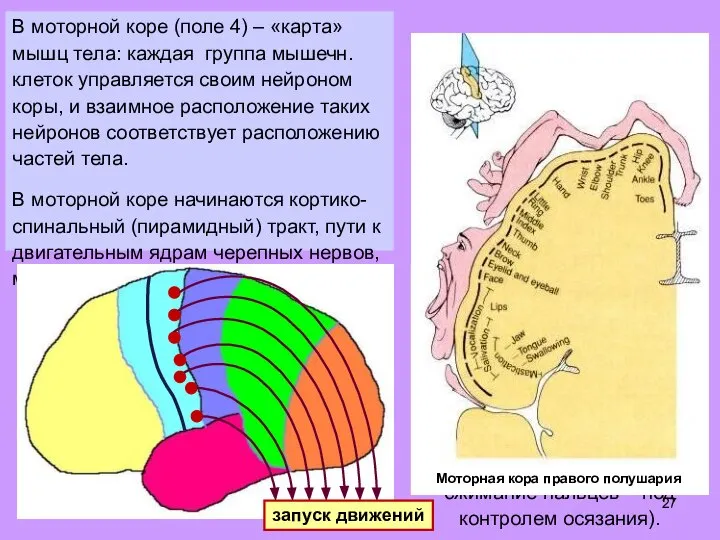
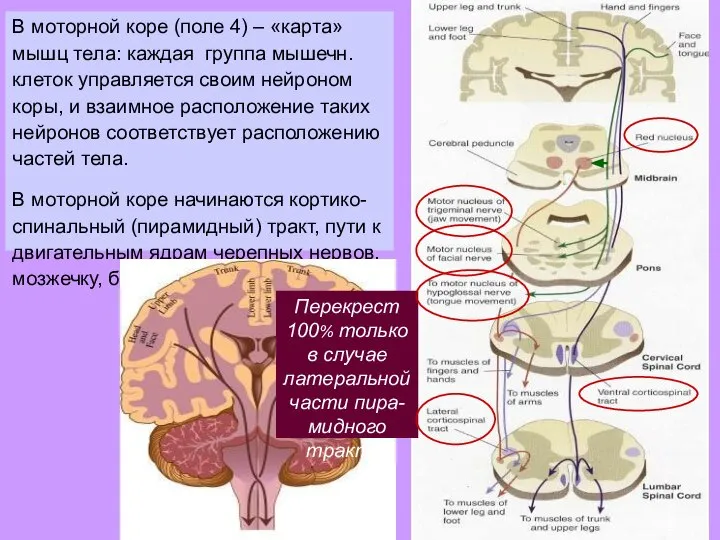
- 27. В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн. клеток управляется своим нейроном
- 28. В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн. клеток управляется своим нейроном
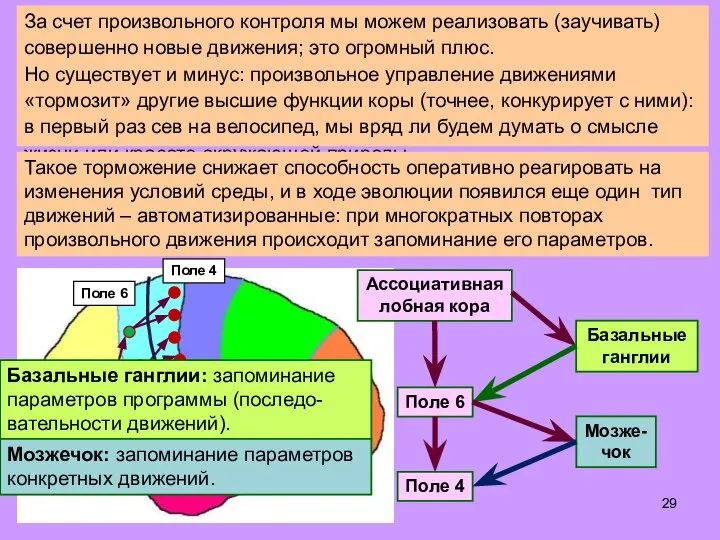
- 29. За счет произвольного контроля мы можем реализовать (заучивать) совершенно новые движения; это огромный плюс. Но существует
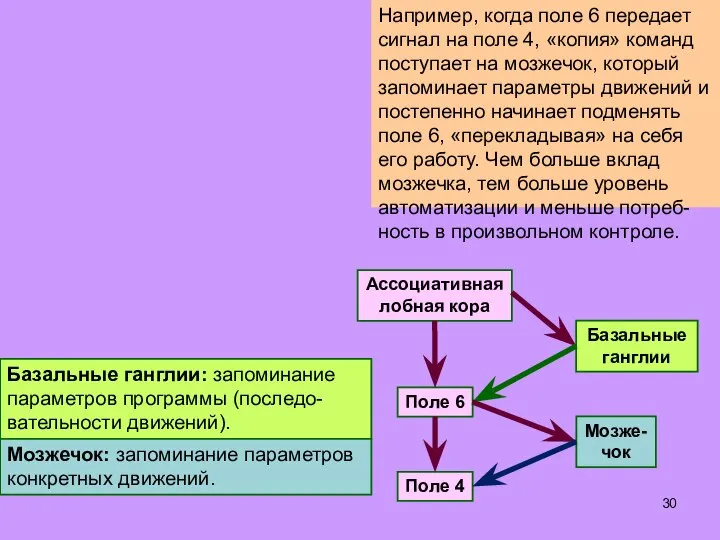
- 30. Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает на мозжечок, который запоминает
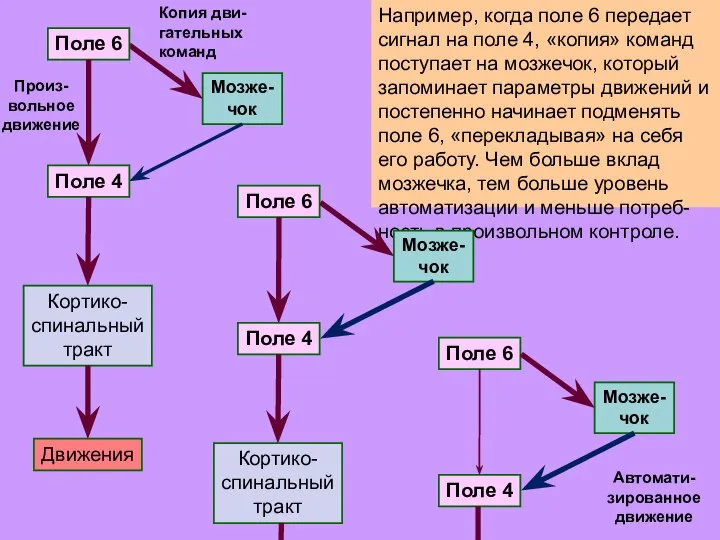
- 31. Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает на мозжечок, который запоминает
- 32. МОЗЖЕЧОК. новая часть [кора наружной зоны полушарий + зубчатые ядра] – автоматизация произвольных движений, запускаемых корой
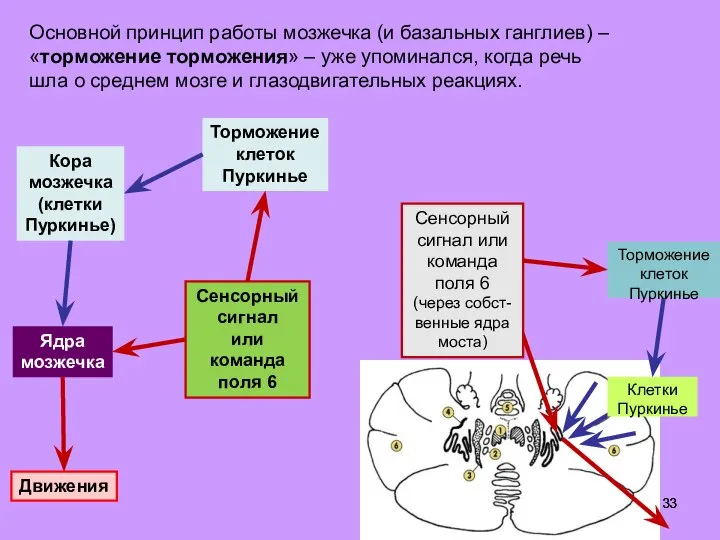
- 33. Основной принцип работы мозжечка (и базальных ганглиев) – «торможение торможения» – уже упоминался, когда речь шла
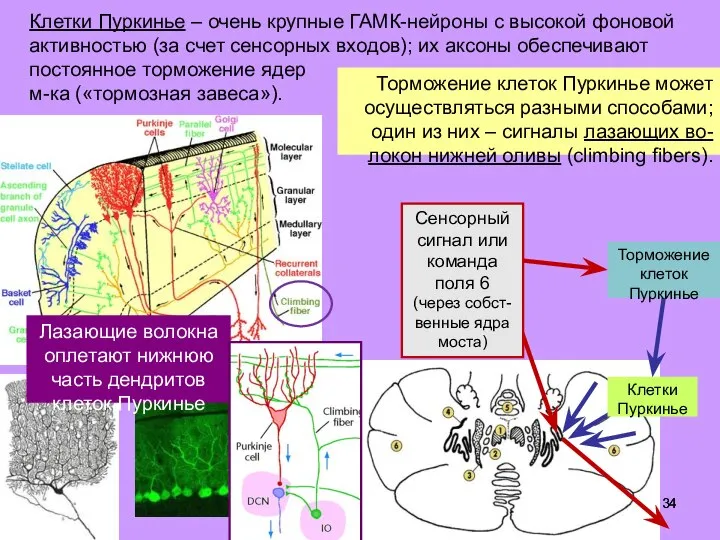
- 34. Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет сенсорных входов); их аксоны
- 35. Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет сенсорных входов); их аксоны
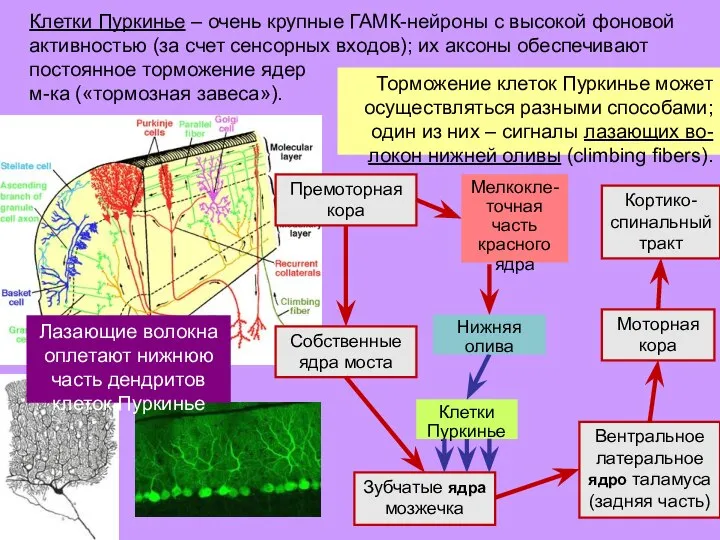
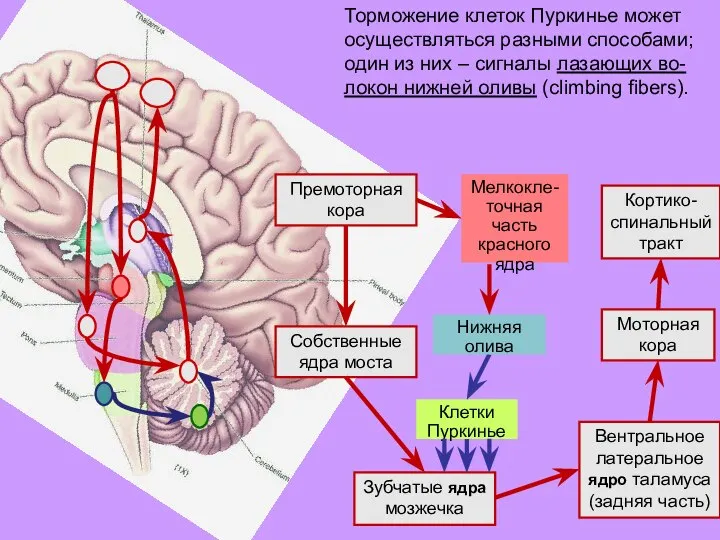
- 36. Торможение клеток Пуркинье может осуществляться разными способами; один из них – сигналы лазающих во-локон нижней оливы
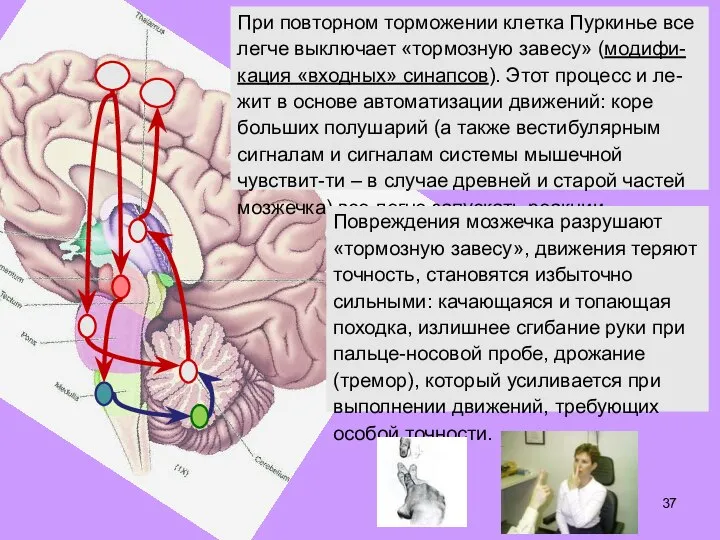
- 37. При повторном торможении клетка Пуркинье все легче выключает «тормозную завесу» (модифи-кация «входных» синапсов). Этот процесс и
- 38. Асс. лобн. кора Премоторная кора Черная субстанция Субталами- ческие ядра
- 39. В этой системе передняя часть вентра-льного латерального ядра таламуса является аналогом зубчатого ядра мозжечка, а нейроны
- 40. Помимо «классического» паркинсонизма, возникающего в результате дегенерации черной субстанции, акинезия, ригидность и тремор наблюдаются также при
- 41. В особо тяжелых случаях возможна таламотомия (разрушение вентрального латерального ядра таламуса для «прерыва-ния» патологических команд). В
- 42. Черная субстанция: положи-тельные эмоции, связанные с движениями.
- 44. Скачать презентацию

Слайд 3произвольные: новые движения в новых условиях; управляет кора больших полушарий, используя сенсорный
произвольные: новые движения в новых условиях; управляет кора больших полушарий, используя сенсорный

Слайд 4Рефлекторные движения:
Возникают в ответ на стимул (врожденно заданный или ставший значимым в
Рефлекторные движения:
Возникают в ответ на стимул (врожденно заданный или ставший значимым в
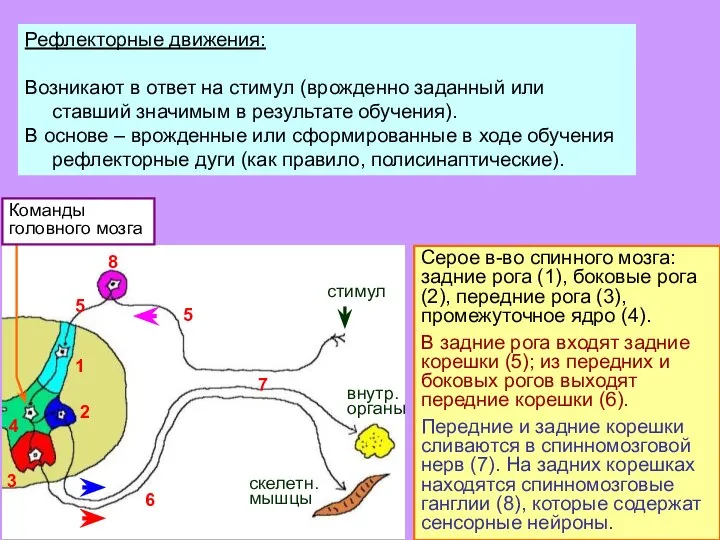
Слайд 5Серое в-во спинного мозга: задние рога (1), боковые рога (2), передние рога
Серое в-во спинного мозга: задние рога (1), боковые рога (2), передние рога
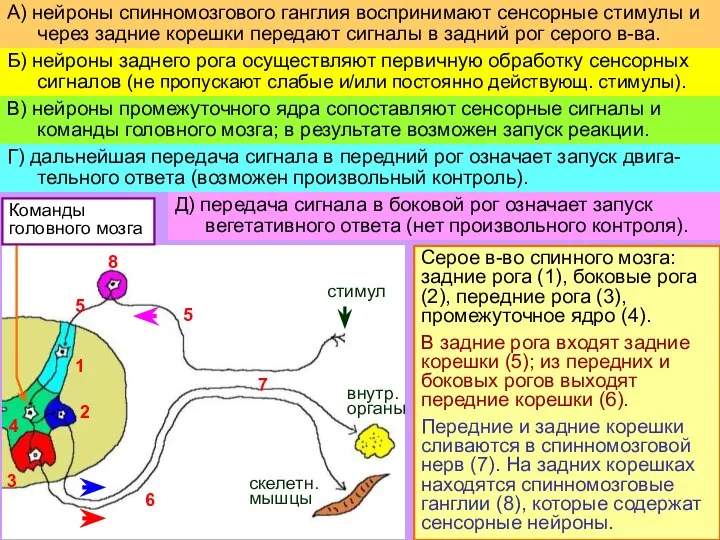
Слайд 6Пример: болевой стимул
запускает сгибание конечнос-ти и вегетативные реакции;
но тормозные влияния
Пример: болевой стимул
запускает сгибание конечнос-ти и вегетативные реакции;
но тормозные влияния
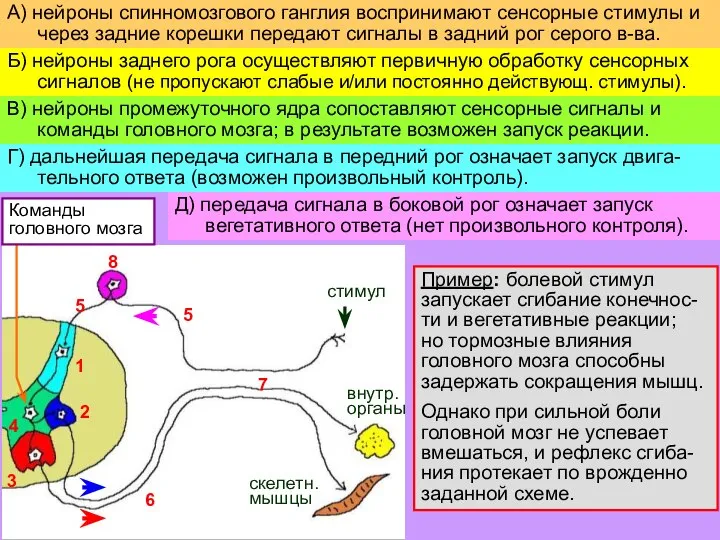
Слайд 7Пример: болевой стимул
запускает сгибание конечнос-ти и вегетативные реакции;
но тормозные влияния
Пример: болевой стимул
запускает сгибание конечнос-ти и вегетативные реакции;
но тормозные влияния
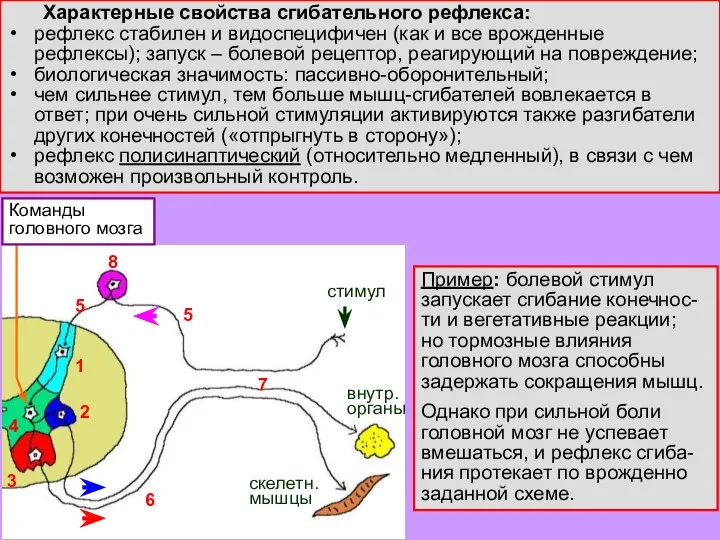
Слайд 8 Характерные свойства сгибательного рефлекса:
рефлекс стабилен и видоспецифичен (как и все врожденные рефлексы);
Характерные свойства сгибательного рефлекса:
рефлекс стабилен и видоспецифичен (как и все врожденные рефлексы);
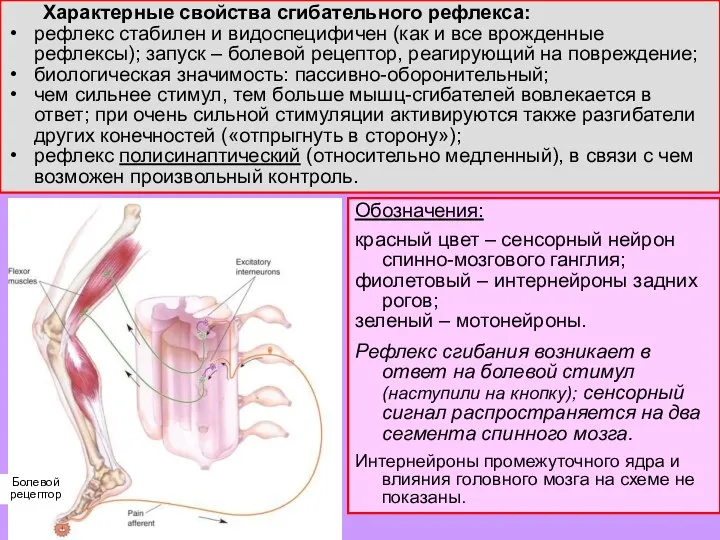
Слайд 9 Коленный рефлекс – частный случай миотатического рефлекса (рефлекса на растяжение мышцы):
мышца (в
Коленный рефлекс – частный случай миотатического рефлекса (рефлекса на растяжение мышцы):
мышца (в
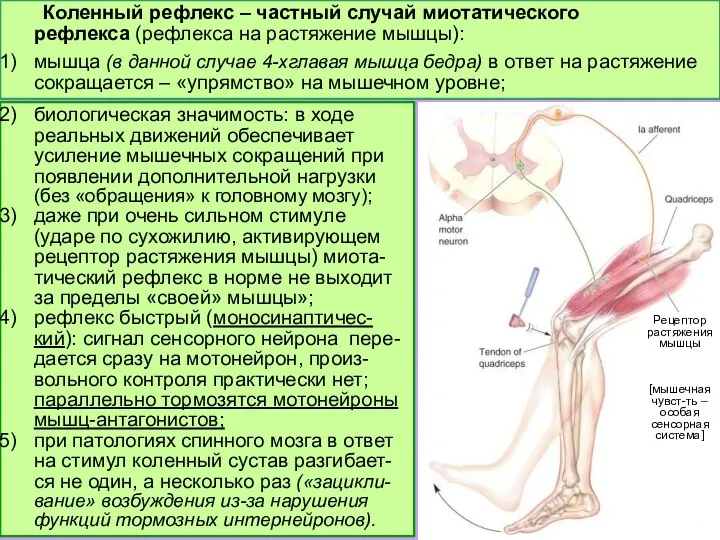
Слайд 10Бицепс: сегменты C5-C6
Трицепс: C6-C8
4-хглавая мышца бедра: L2-L4
Бицепс: сегменты C5-C6
Трицепс: C6-C8
4-хглавая мышца бедра: L2-L4
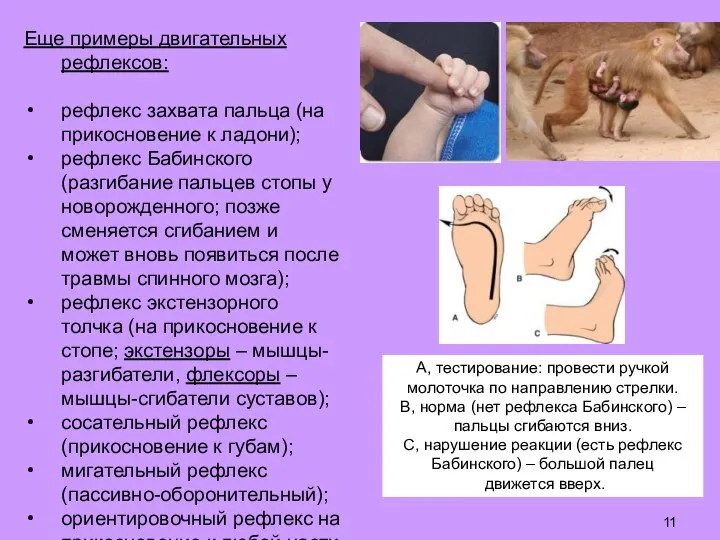
Слайд 11Еще примеры двигательных рефлексов:
рефлекс захвата пальца (на прикосновение к ладони);
рефлекс Бабинского (разгибание
Еще примеры двигательных рефлексов:
рефлекс захвата пальца (на прикосновение к ладони);
рефлекс Бабинского (разгибание
Слайд 12Л о к о м о ц и я :
ритмически повторяющиеся движения
Л о к о м о ц и я :
ритмически повторяющиеся движения

Слайд 13Генераторы локомоторного ритма:
два основных варианта – пейспекеры
и «полуцентры конечностей».
Генераторы локомоторного ритма:
два основных варианта – пейспекеры
и «полуцентры конечностей».
Слайд 14У рыб:
нейроны-пейсмекеры
находятся в продол-говатом мозге, а их ПД распространяются по спинному мозгу.
У
У рыб:
нейроны-пейсмекеры
находятся в продол-говатом мозге, а их ПД распространяются по спинному мозгу.
У
Слайд 15Более гибкий вариант – полуцентры конечностей.
Здесь не только конкурентное (реципрокное) торможение, но
Более гибкий вариант – полуцентры конечностей.
Здесь не только конкурентное (реципрокное) торможение, но
Слайд 16При низкой активации гол. мозга работает только реципрокное тор-можение = разгибание.
При достаточно
При низкой активации гол. мозга работает только реципрокное тор-можение = разгибание.
При достаточно
Слайд 17Запуск локомоции – за счет тонических влияний головного мозга.
Траекторию «восьмерки» создают цепи
Траекторию «восьмерки» создают цепи
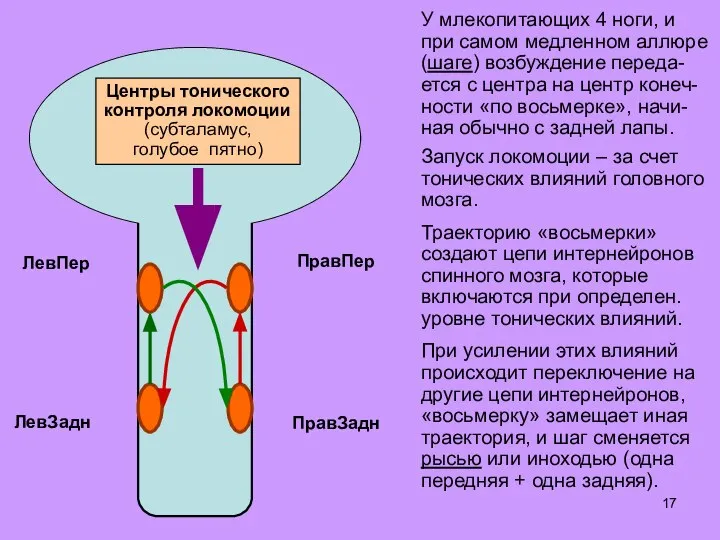
Слайд 18Запуск локомоции – за счет тонических влияний головного мозга.
Траекторию «восьмерки» создают цепи
Траекторию «восьмерки» создают цепи
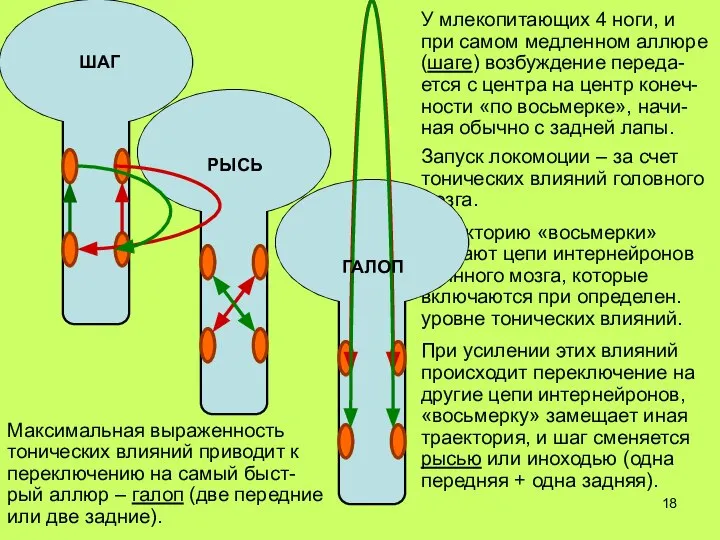
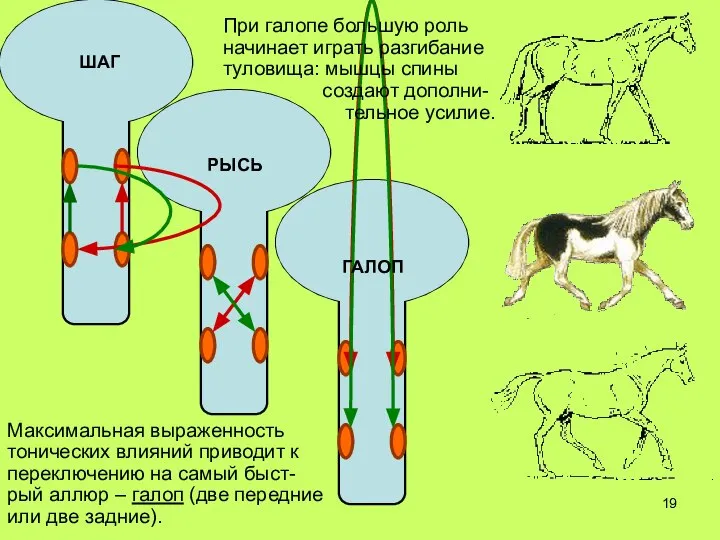
Слайд 19РЫСЬ
Максимальная выраженность тонических влияний приводит к переключению на самый быст-рый аллюр –
РЫСЬ
Максимальная выраженность тонических влияний приводит к переключению на самый быст-рый аллюр –
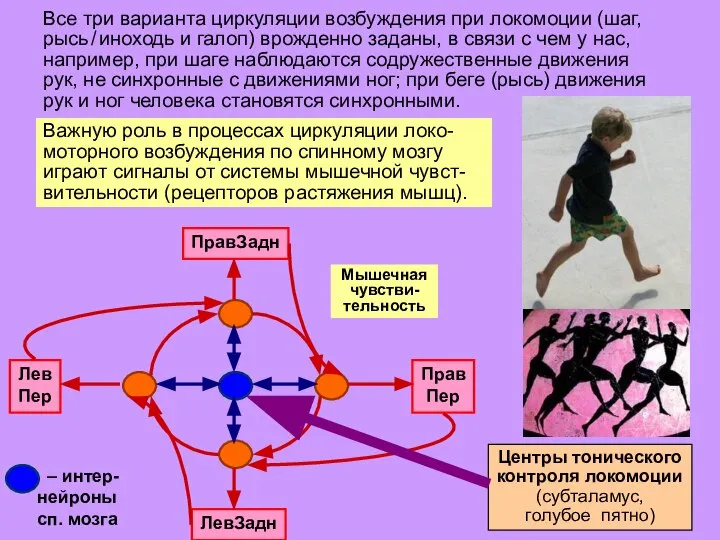
Слайд 20Мышечная
чувстви-тельность
Все три варианта циркуляции возбуждения при локомоции (шаг, рысь / иноходь и
Мышечная
чувстви-тельность
Все три варианта циркуляции возбуждения при локомоции (шаг, рысь / иноходь и
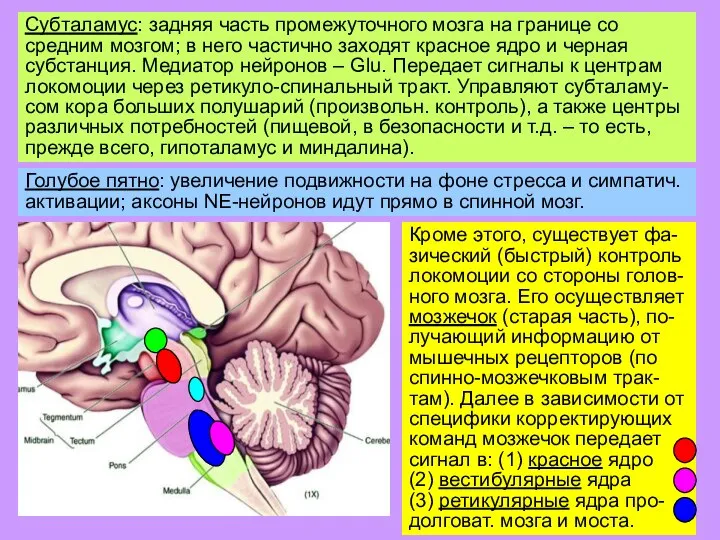
Слайд 21Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него
Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него
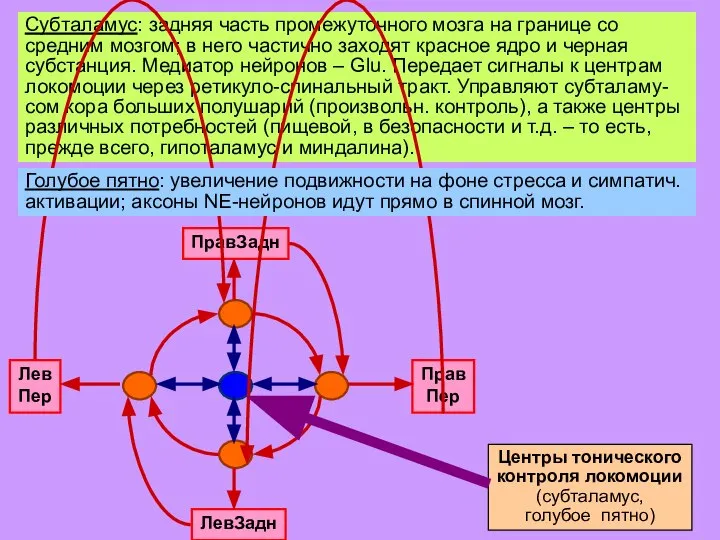
Слайд 22Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него
Субталамус: задняя часть промежуточного мозга на границе со средним мозгом; в него
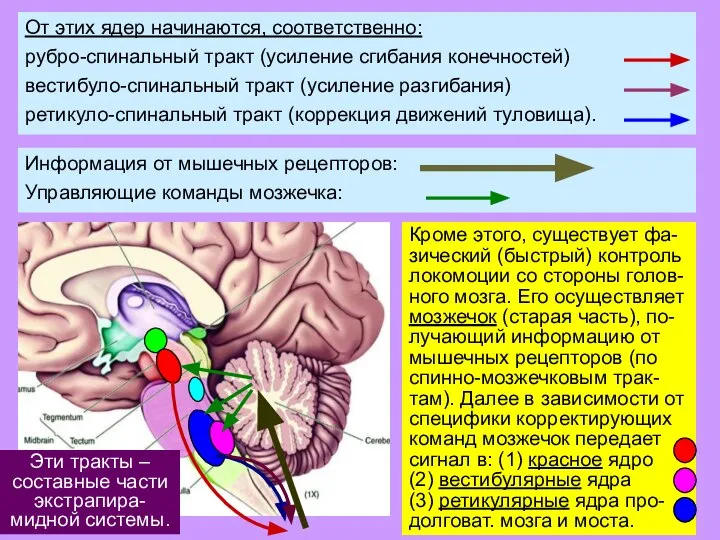
Слайд 23Эти тракты –
составные части экстрапира-
мидной системы.
Эти тракты –
составные части экстрапира-
мидной системы.
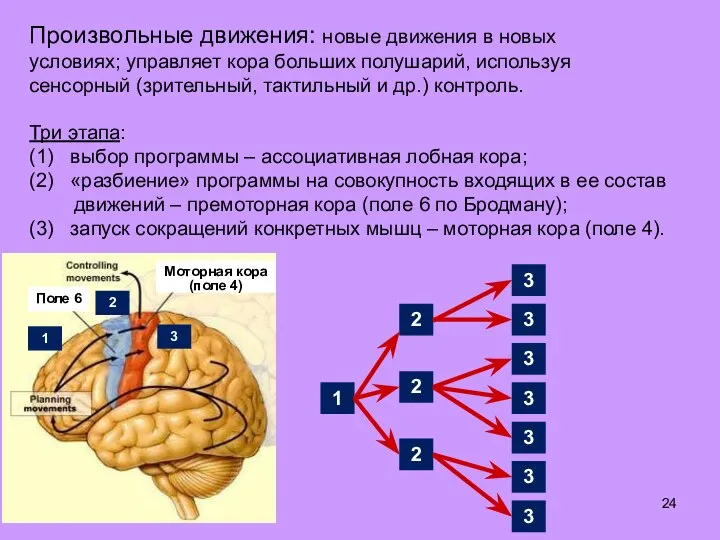
Слайд 24Произвольные движения: новые движения в новых условиях; управляет кора больших полушарий, используя
Произвольные движения: новые движения в новых условиях; управляет кора больших полушарий, используя
Слайд 25(1) «взять предмет»
(2) [ шагнуть + вытянуть руку + распрямить пальцы
(1) «взять предмет»
(2) [ шагнуть + вытянуть руку + распрямить пальцы
Слайд 26(1) «взять предмет»
(2) [ шагнуть + вытянуть руку + распрямить пальцы
(1) «взять предмет»
(2) [ шагнуть + вытянуть руку + распрямить пальцы
Слайд 27В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн.
В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн.
Слайд 28В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн.
В моторной коре (поле 4) – «карта» мышц тела: каждая группа мышечн.
Слайд 29За счет произвольного контроля мы можем реализовать (заучивать) совершенно новые движения; это
За счет произвольного контроля мы можем реализовать (заучивать) совершенно новые движения; это
Слайд 30Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает
Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает
Слайд 31Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает
Например, когда поле 6 передает сигнал на поле 4, «копия» команд поступает
Слайд 32МОЗЖЕЧОК.
новая часть [кора наружной зоны полушарий + зубчатые ядра] – автоматизация произвольных
МОЗЖЕЧОК.
новая часть [кора наружной зоны полушарий + зубчатые ядра] – автоматизация произвольных
![МОЗЖЕЧОК. новая часть [кора наружной зоны полушарий + зубчатые ядра] – автоматизация](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1049996/slide-31.jpg)
Слайд 33Основной принцип работы мозжечка (и базальных ганглиев) – «торможение торможения» – уже
Основной принцип работы мозжечка (и базальных ганглиев) – «торможение торможения» – уже
Слайд 34Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет
Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет
Слайд 35Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет
Клетки Пуркинье – очень крупные ГАМК-нейроны с высокой фоновой активностью (за счет
Слайд 36Торможение клеток Пуркинье может осуществляться разными способами; один из них – сигналы
Торможение клеток Пуркинье может осуществляться разными способами; один из них – сигналы
Слайд 37При повторном торможении клетка Пуркинье все легче выключает «тормозную завесу» (модифи-кация «входных»
При повторном торможении клетка Пуркинье все легче выключает «тормозную завесу» (модифи-кация «входных»
Слайд 38Асс.
лобн.
кора
Премоторная кора
Черная
субстанция
Субталами-
ческие ядра
Асс.
лобн.
кора
Премоторная кора
Черная
субстанция
Субталами-
ческие ядра
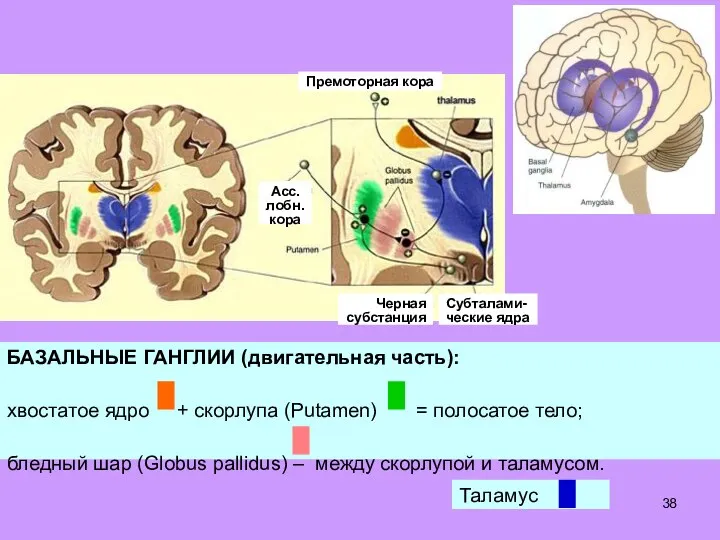
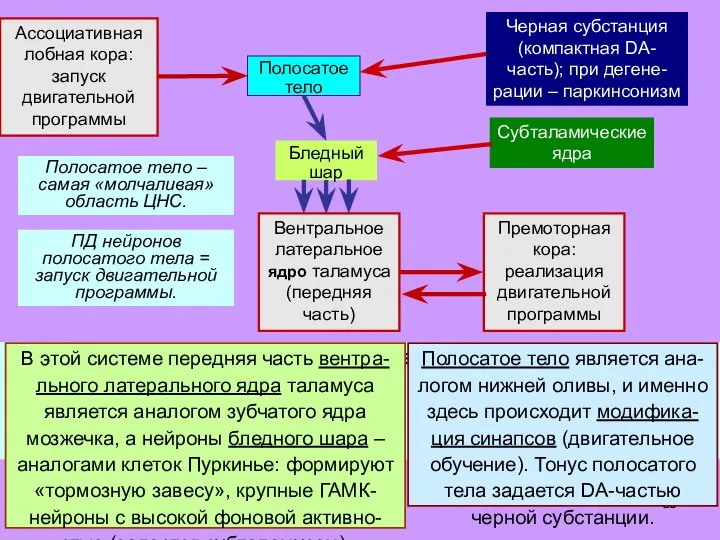
Слайд 39В этой системе передняя часть вентра-льного латерального ядра таламуса является аналогом зубчатого
В этой системе передняя часть вентра-льного латерального ядра таламуса является аналогом зубчатого
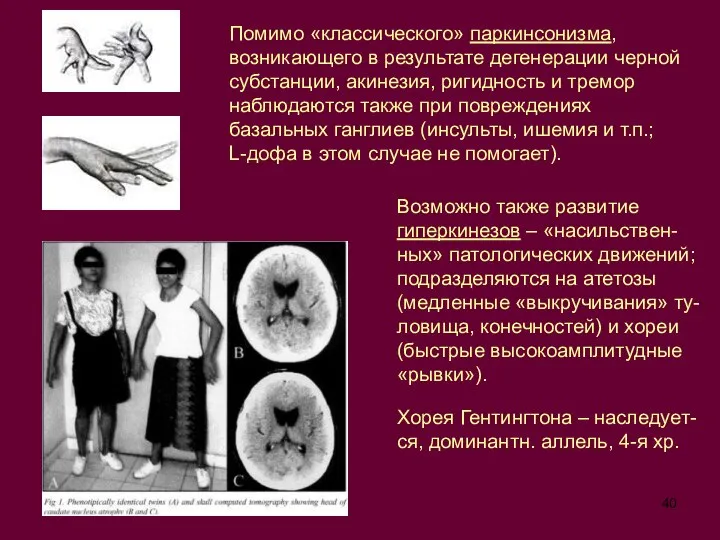
Слайд 40Помимо «классического» паркинсонизма, возникающего в результате дегенерации черной субстанции, акинезия, ригидность и
Помимо «классического» паркинсонизма, возникающего в результате дегенерации черной субстанции, акинезия, ригидность и
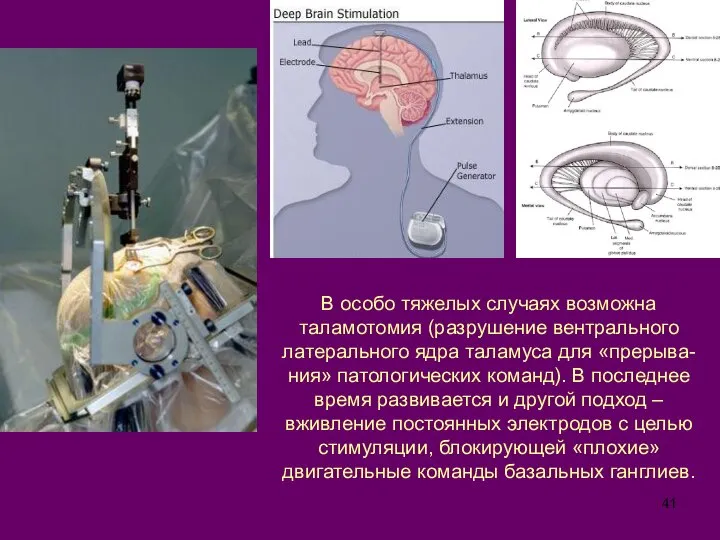
Слайд 41В особо тяжелых случаях возможна таламотомия (разрушение вентрального латерального ядра таламуса для
В особо тяжелых случаях возможна таламотомия (разрушение вентрального латерального ядра таламуса для
Слайд 42Черная субстанция: положи-тельные эмоции, связанные с движениями.
Черная субстанция: положи-тельные эмоции, связанные с движениями.

 Насекомые вредители на станции юных натуралистов г. Тайшета
Насекомые вредители на станции юных натуралистов г. Тайшета Рыбы и насекомые пресных водоемов
Рыбы и насекомые пресных водоемов Грибы, цветы и ягоды леса
Грибы, цветы и ягоды леса Чей хвостик
Чей хвостик Семейство крестоцветных
Семейство крестоцветных Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба
Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба Неклеточные формы жизни
Неклеточные формы жизни Раселение живых организмов
Раселение живых организмов Эпителий
Эпителий Презентация на тему Строение и химический состав клетки
Презентация на тему Строение и химический состав клетки  MOLEKUlyarnaya_biologia_LPZ_1
MOLEKUlyarnaya_biologia_LPZ_1 Фотодыхание
Фотодыхание Вирусология - наука о вирусах
Вирусология - наука о вирусах Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Чабрец (богородская трава, тимьян ползучий)
Чабрец (богородская трава, тимьян ползучий) Составление генетических паспортов и установление родственных связей
Составление генетических паспортов и установление родственных связей От клюва до хвоста
От клюва до хвоста Модификационная изменчивость
Модификационная изменчивость Строение и многообразие грибов
Строение и многообразие грибов Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности
Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности  Что было на Земле, когда она появилась?
Что было на Земле, когда она появилась? Биохимия. Разделы биохимии. Становление биохимии как науки
Биохимия. Разделы биохимии. Становление биохимии как науки История собаководства
История собаководства Морфология бактерий
Морфология бактерий Жизненная Ёмкость Лёгких
Жизненная Ёмкость Лёгких Ткани животных
Ткани животных 2. Урок - .ДНК
2. Урок - .ДНК Лишайники
Лишайники