- Эволюционная морфология растений

Содержание
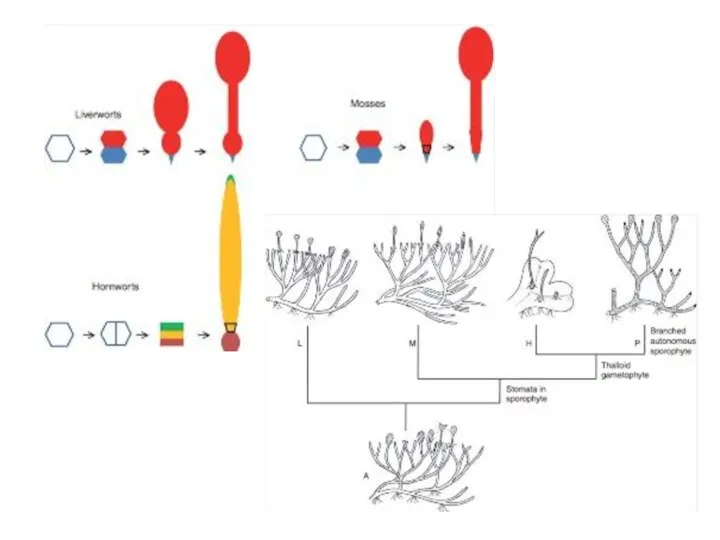
- 2. Ландшафт среднего раннего Девона. Раннедевонские ( 408 - 387 млн лет назад) Cooksonia, Zosterophyllum, Aglaophyton. Среднедевонский
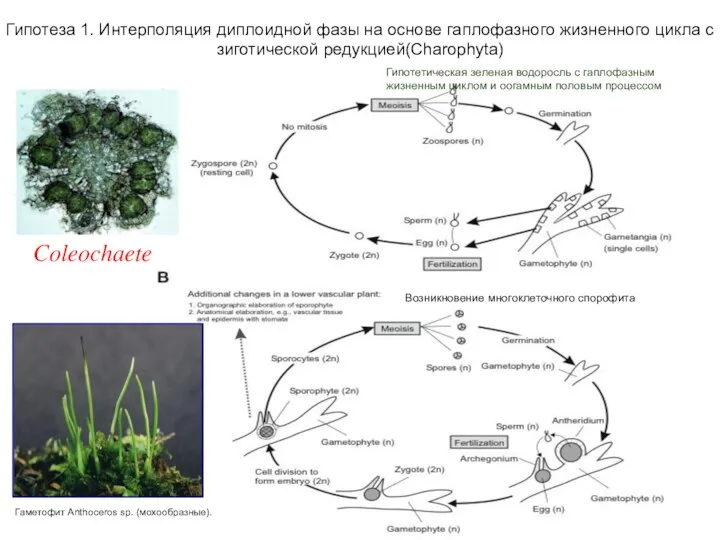
- 3. Гипотеза 1. Интерполяция диплоидной фазы на основе гаплофазного жизненного цикла с зиготической редукцией(Charophyta) Гипотетическая зеленая водоросль
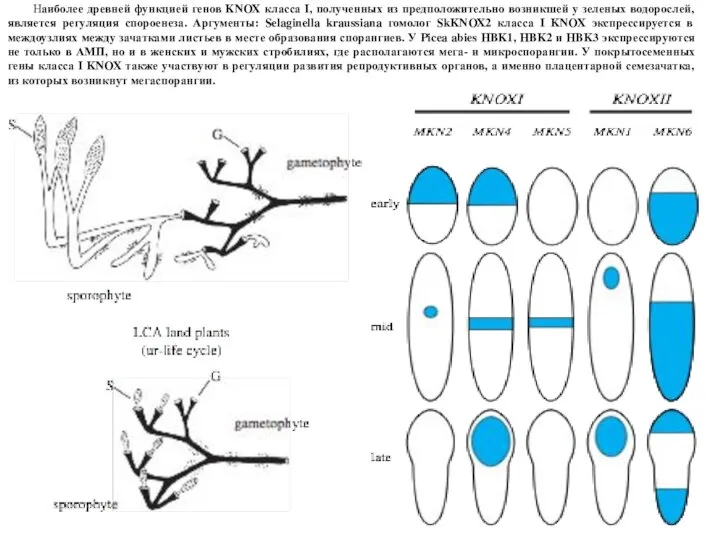
- 4. Наиболее древней функцией генов KNOX класса I, полученных из предположительно возникшей у зеленых водорослей, является регуляция
- 5. Способность к ветвлению - общая черта спорофитов всех высших растений спорофитной линии эволюции. Роль - вегетативное
- 6. Поскольку апикальная меристема побега (АМП) играет исключительную роль в формировании побеговых систем растений, изучение ее функции
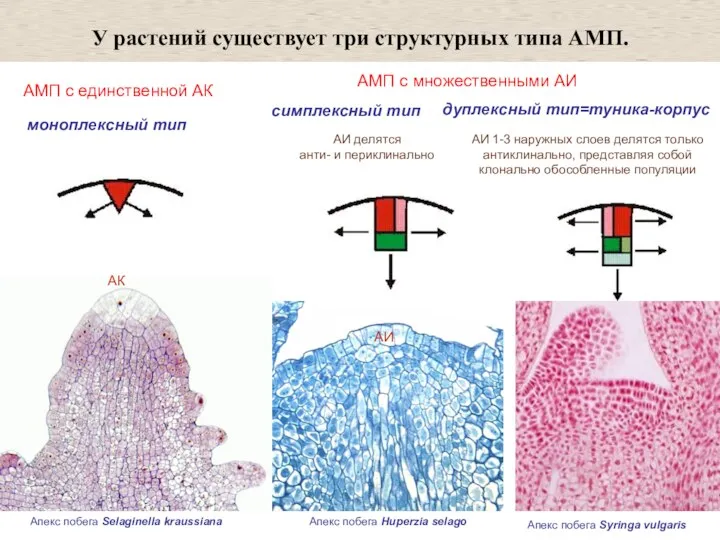
- 7. АМП с единственной АК АМП с множественными АИ У растений существует три структурных типа АМП. АИ
- 8. Phegopteris connectilis Huperzia selago Selaginella kraussiana Pinus sylvestris CZ, ЦЗ central zone PZ, ПЗ периферическая зона
- 9. Matteuccia struthiopteris 10 µm АК и ее ближайшие производные в моноплексной АМП характеризуются нетипичной для клеток
- 10. P. aquilinum, Фрагмент антиклинальной стенки клетки чашевидной зоны Selaginella kraussiana Matteuccia struthiopteris, фрагмент антиклинальной стенки ПИ
- 11. Evkaikina, Romanova, Voitsekhovskaja Frontiers in PlantScience, 2014 В АМП с несколькими АИ производные разных инициалей могут
- 12. ИСХОДНЫЙ ТИП Моноплексаная АМП предполагаемого предка - харовой водоросли По Graham et al, 2000 Вопрос об
- 13. Если первые наземные растения имели симплексную АМП, моноплексные АМП Selaginellaceae и Monilophytes - примеры независимого возникновения,
- 14. Таксономическая приуроченность разных типов АМП и их связь с морфологией не очевидны. АМП с единственной и
- 15. Клеточные аспекты заложения листа коррелируют с типом АМП, но не с морфологической природой листа Monilophytes Huperzia
- 16. Оценить эволюционный уровень разных структурных типов АМП невозможно вне сравнения механизмов регуляции
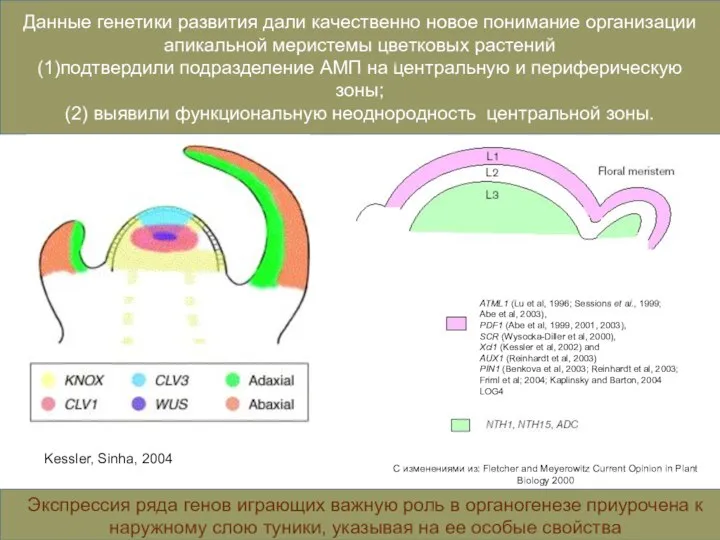
- 17. Kessler, Sinha, 2004 Данные генетики развития дали качественно новое понимание организации апикальной меристемы цветковых растений (1)подтвердили
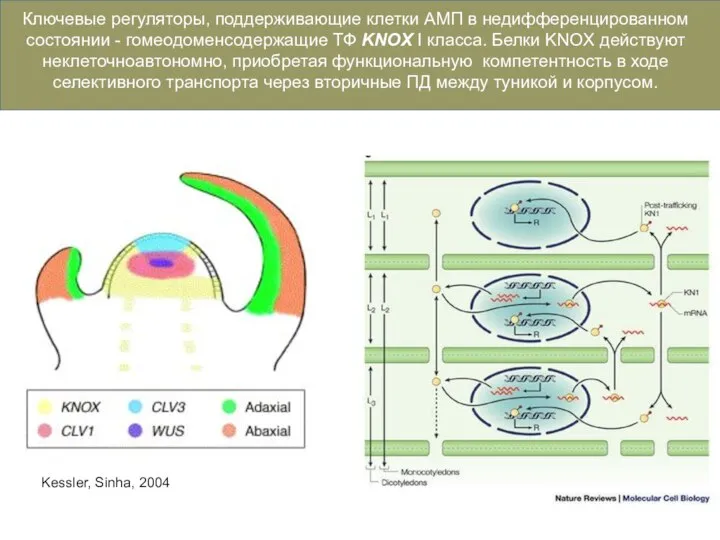
- 18. Kessler, Sinha, 2004 Ключевые регуляторы, поддерживающие клетки АМП в недифференцированном состоянии - гомеодоменсодержащие ТФ KNOX I
- 19. The protein models of native HD (C) and double HD (Q30R and L53Q) mutant (D) were
- 20. Результаты изучения АМП в эволюционном контексте значительно скромнее. Гипотезы о механизмах функционирования АМП у высших споровых
- 21. (Harrison et al, 2005) CrKNOX1 mRNA SkKNOX1 mRNA Zamia floridans (from Bharathan et al, 2002 (Sano
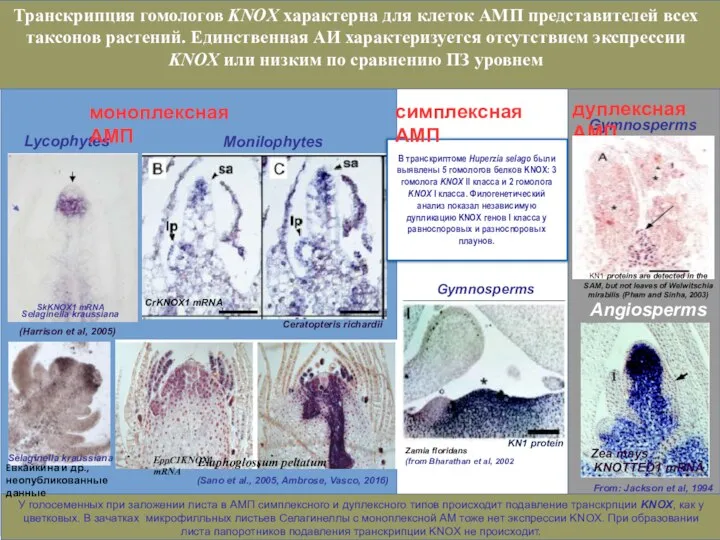
- 22. Отсутствуют данные о генетической регуляции в меристеме симплексного типа плауноообразных. Нами был секвенирован транскриптом верхушек побегов
- 23. Evkaikina et al, unpublished Последовательности гомологов обоих классов у Lycopodiophyta являются сестринскими по отношению к последовательностям
- 24. Филогенетический анализ показал независимую дупликацию KNOX генов I класса у равноспоровых и разноспоровых плаунов. Evkaikina et
- 25. Апекс побега Huperzia selago симплексный тип Логично предположить, что дупликация KNOX генов I класса у Lycopodiales
- 26. Апекс побега Selaginella kraussiana Апекс побега Huperzia selago моноплексный тип симплексный тип Однако ни в транскриптоме
- 27. From Frank et al, 2015 СЕКВЕНИРОВАНИЕ И АНАЛИЗ ТРАНСКРИПТОМОВ КЛЕТОК РАЗНЫХ ЗОН МОНОПЛЕКСНОЙ АМП У ПЛАУНОВ
- 28. АК И ПЗ ПРЕДСТАВЛЯЮТ СОБОЙ РАЗЛИЧАЮЩИЕСЯ ТРАНСКРИПЦИОННЫЕ ДОМЕНЫ. ПЗ МОНОПЛЕКСНОЙ АМП ТРАНСКРИПЦИОННО СХОДЕН С ПЗ ДУПЛЕКСНОЙ
- 29. ГОМОЛОГИ СLV3-LIKE (CLE) ГЕНОВ. monoplex SAM simplex SAM duplex SAM Expression of CLE209 and CLE210 in
- 30. ГОМОЛОГИ WUSCHEL-LIKE HOMEOBOX (WOX) ГЕНОВ monoplex SAM simplex SAM duplex SAM Quantitative real-time PCR of P.
- 31. Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений Листья всех растений имеют
- 32. Huperzia selago ЦЗ ПЗ ПЗ АИ Один из критериев энационного листа - возникновение ниже апикальной меристемы.
- 33. Вопреки классической точке зрения о принципиально разном способе закладке микро- и макрофилльных листьев, они закладываются сходным
- 34. Гаплостела Риниевые Актиностела Псилотовые, плауны Эвстела Голосеменные, Двудольные Атактостела Однодольные Диктиостела Папоротники Плектостела Плауны Артростела Хвощи
- 35. Актиностела Sphenophyllum Актиностела со “смешанной сердцевиной” Lepidodendron Эвстела каменноугольного семенного папоротника Протостела аневрофита Протостела Девонского семенного
- 36. Переключение между программами “меристематичности” и органогенеза определяется антагонистическим взаимодействием между ТФ KNOX и двумя группами “листовых”
- 37. SkARP1 SkKNOX1 SkARP and SkKNOX in Selaginella kraussiana ARP proteins in Osmunda regalis co-localized with Class
- 38. monoplex SAM simplex SAM duplex SAM PHAB and PHAV From Floyd, Bowman, 2006 Pseudotsuga menziensii Ginkgo
- 39. Selaginella moellendorfiii SmC3HDZ32 mRNA SmC3HDZ31 mRNA Psilotum nudum PnC3HDZ2 mRNA From: Vasco et al, 2016 PnC3HDZ3
- 40. Lycophytes Gymnosperms Monilophytes From Finet et al, 2016 PmeYABC Экспрессия гомологов “листовых” YABBY у голосеменных с
- 41. Филогения белков YABBY у высших растений. В транскриптоме Huperzia selago впервые для несеменных растений нами был
- 42. Имеющиеся на настоящий момент молекулярно-генетические данные показали, что функциональная регуляция АМП в разных таксонах имеет специфические
- 43. Колокализация гомологов KNOX и ARP в АМП плаунов и папоротников указывает на то, что их исходная
- 44. экспрессия гомологов HD-Zip III в зачатках листьев и спорангиев плаунов и Monilophytes рассматривается как подтверждение гипотезы
- 45. KNOX/ARP и KNOX/YABBY взаимодействия были задействованы независимо у плаунообразных для поэтапного возникновения выростов-микрофиллов и у остальных
- 46. From: A. Tomescu Trends in Plant Science. 2009 различия в экспрессии KNOX у плаунообразных, папоротникообразных и
- 47. Наличие в транскриптоме Huperzia гомологов ТФ YABBY, а у Selaginella ARP, позволяет предположить что у их
- 48. 1, неравная дихотомия (перевершинивание. Возможно результат усложнения механизмов поддержания меристем. 2, дифференцировка на побеги с ограниченным
- 49. 5, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов адаксиальности” (напр homeodomain-zip gene
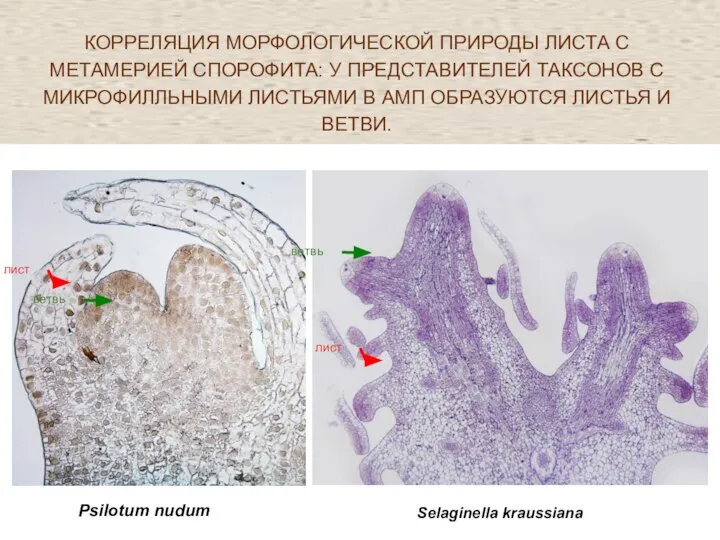
- 50. КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА: У ПРЕДСТАВИТЕЛЕЙ ТАКСОНОВ С МИКРОФИЛЛЬНЫМИ ЛИСТЬЯМИ В АМП ОБРАЗУЮТСЯ
- 51. КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА Phegopteris connectilis лист лист корень почка почка корень корень
- 52. Биполярный рост и “геммаксилярная” метамерия скоррелированы с развитием зародыша внутри семени
- 53. 5, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов адаксиальности” (напр homeodomain-zip gene
- 54. Элементарный метамер семенных растений - лист и пазушая почка ОРГАНОГЕНЕЗ Наименьшая повторяющаяся структурная единица, образующаяся из
- 61. Скачать презентацию

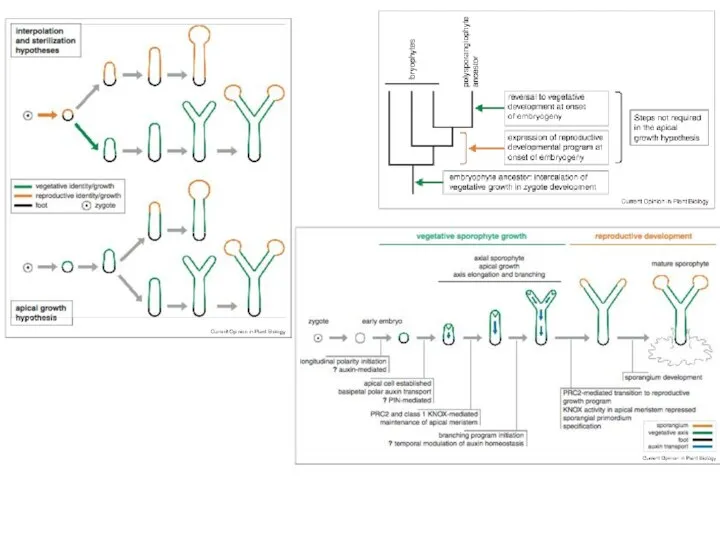
Слайд 3Гипотеза 1. Интерполяция диплоидной фазы на основе гаплофазного жизненного цикла с зиготической
Гипотеза 1. Интерполяция диплоидной фазы на основе гаплофазного жизненного цикла с зиготической
Слайд 4Наиболее древней функцией генов KNOX класса I, полученных из предположительно возникшей у
Наиболее древней функцией генов KNOX класса I, полученных из предположительно возникшей у
Слайд 5 Способность к ветвлению - общая черта спорофитов всех высших растений спорофитной линии
Способность к ветвлению - общая черта спорофитов всех высших растений спорофитной линии

Слайд 6Поскольку апикальная меристема побега (АМП) играет исключительную роль в формировании побеговых систем
Поскольку апикальная меристема побега (АМП) играет исключительную роль в формировании побеговых систем
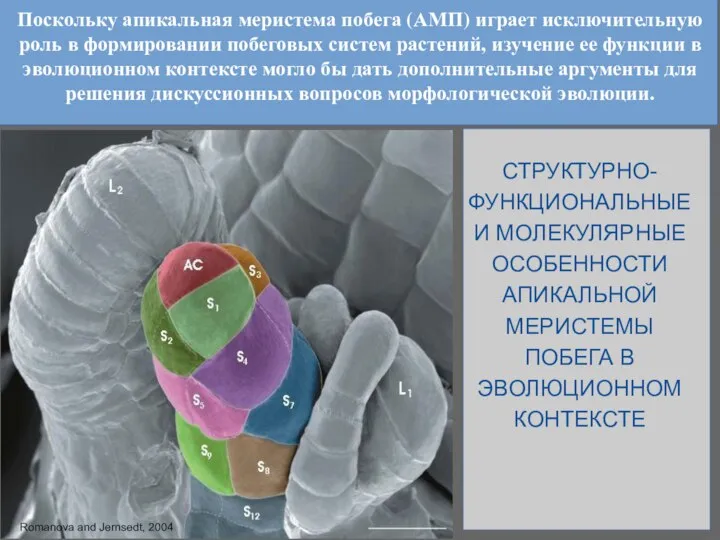
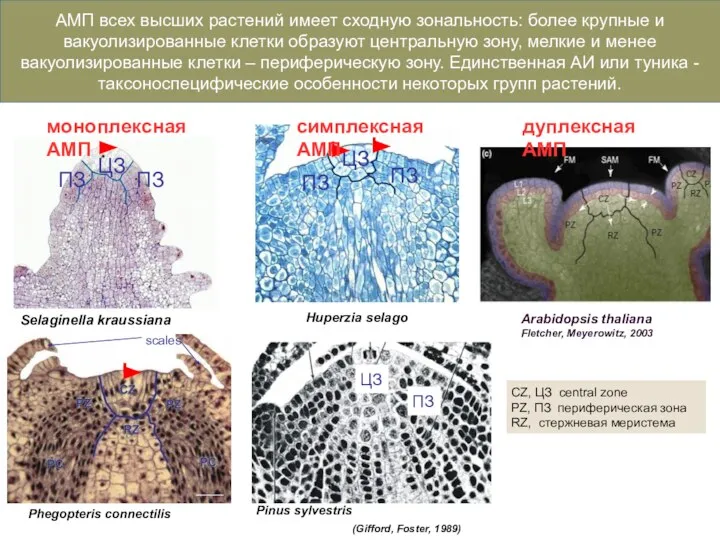
Слайд 7АМП с единственной АК
АМП с множественными АИ
У растений существует три структурных типа
АМП с единственной АК
АМП с множественными АИ
У растений существует три структурных типа
Слайд 8Phegopteris connectilis
Huperzia selago
Selaginella kraussiana
Pinus sylvestris
CZ, ЦЗ central zone
PZ, ПЗ периферическая зона
RZ, стержневая
Phegopteris connectilis
Huperzia selago
Selaginella kraussiana
Pinus sylvestris
CZ, ЦЗ central zone
PZ, ПЗ периферическая зона
RZ, стержневая
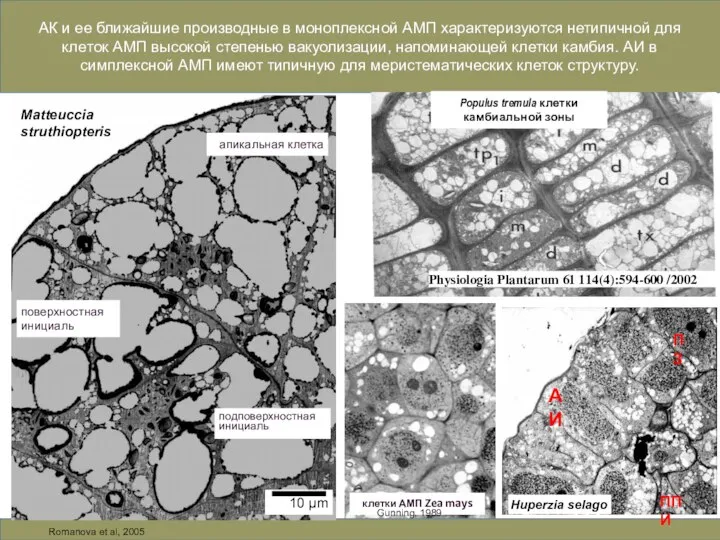
Слайд 9Matteuccia
struthiopteris
10 µm
АК и ее ближайшие производные в моноплексной АМП характеризуются нетипичной для
Matteuccia
struthiopteris
10 µm
АК и ее ближайшие производные в моноплексной АМП характеризуются нетипичной для
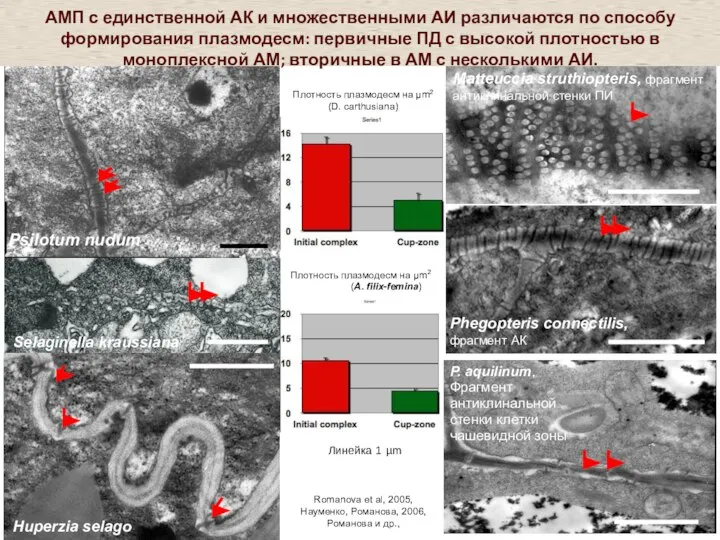
Слайд 10P. aquilinum,
Фрагмент антиклинальной стенки клетки чашевидной зоны
Selaginella kraussiana
Matteuccia struthiopteris, фрагмент антиклинальной
P. aquilinum,
Фрагмент антиклинальной стенки клетки чашевидной зоны
Selaginella kraussiana
Matteuccia struthiopteris, фрагмент антиклинальной
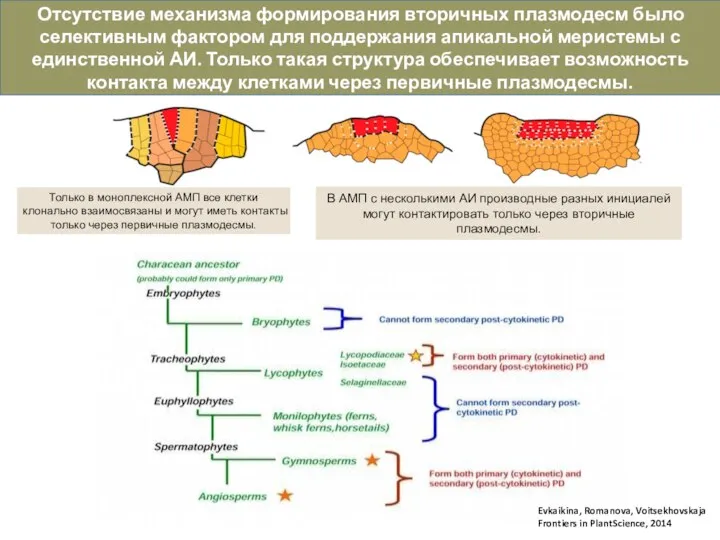
Слайд 11Evkaikina, Romanova, Voitsekhovskaja
Frontiers in PlantScience, 2014
В АМП с несколькими АИ производные
Evkaikina, Romanova, Voitsekhovskaja
Frontiers in PlantScience, 2014
В АМП с несколькими АИ производные
Слайд 12ИСХОДНЫЙ ТИП
Моноплексаная АМП предполагаемого предка - харовой водоросли
По Graham et al,
ИСХОДНЫЙ ТИП
Моноплексаная АМП предполагаемого предка - харовой водоросли
По Graham et al,
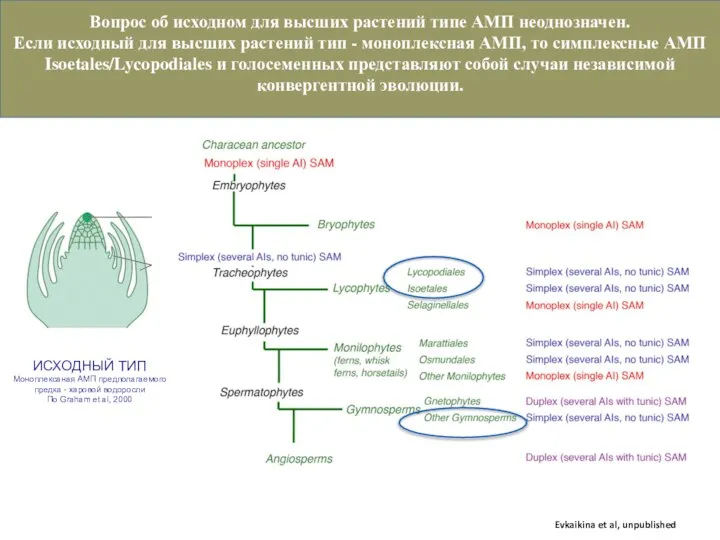
Слайд 13Если первые наземные растения имели симплексную АМП, моноплексные АМП Selaginellaceae и Monilophytes
Если первые наземные растения имели симплексную АМП, моноплексные АМП Selaginellaceae и Monilophytes
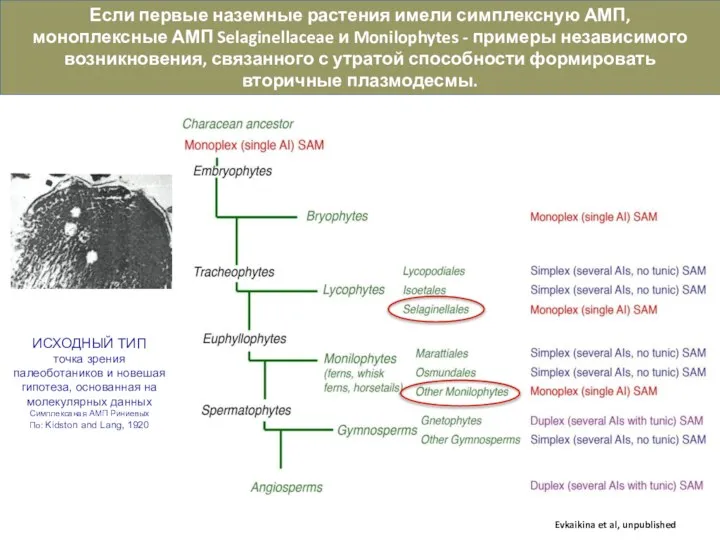
Слайд 14Таксономическая приуроченность разных типов АМП и их связь с морфологией не очевидны.
Таксономическая приуроченность разных типов АМП и их связь с морфологией не очевидны.
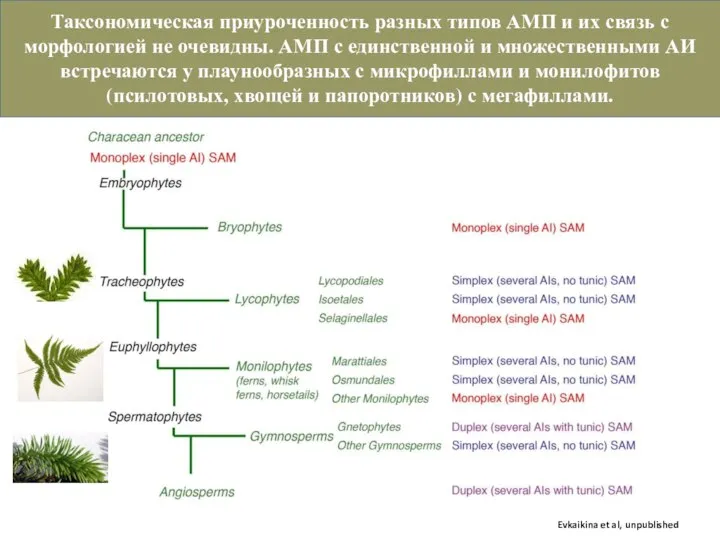
Слайд 15Клеточные аспекты заложения листа коррелируют с типом АМП, но не с морфологической
Клеточные аспекты заложения листа коррелируют с типом АМП, но не с морфологической
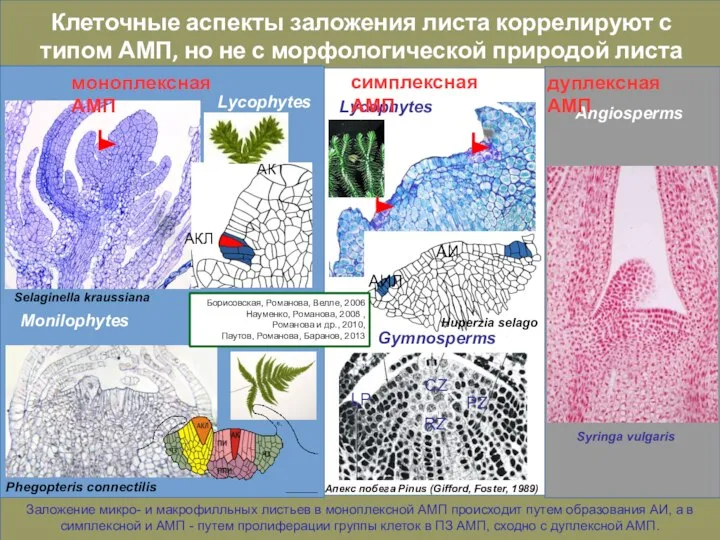
Слайд 16Оценить эволюционный уровень разных структурных типов АМП невозможно вне сравнения механизмов регуляции
Оценить эволюционный уровень разных структурных типов АМП невозможно вне сравнения механизмов регуляции

Слайд 17Kessler, Sinha, 2004
Данные генетики развития дали качественно новое понимание организации апикальной меристемы
Kessler, Sinha, 2004
Данные генетики развития дали качественно новое понимание организации апикальной меристемы
Слайд 18Kessler, Sinha, 2004
Ключевые регуляторы, поддерживающие клетки АМП в недифференцированном состоянии - гомеодоменсодержащие
Kessler, Sinha, 2004
Ключевые регуляторы, поддерживающие клетки АМП в недифференцированном состоянии - гомеодоменсодержащие
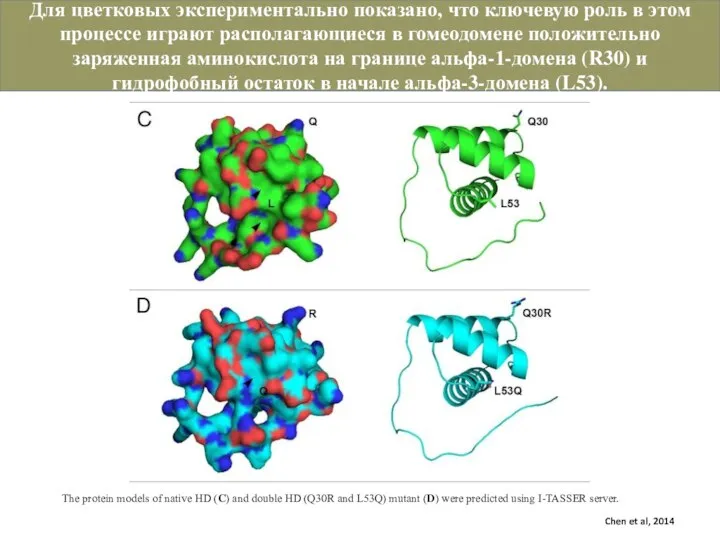
Слайд 19The protein models of native HD (C) and double HD (Q30R and
The protein models of native HD (C) and double HD (Q30R and
Слайд 20Результаты изучения АМП в эволюционном контексте значительно скромнее.
Гипотезы о механизмах функционирования
Результаты изучения АМП в эволюционном контексте значительно скромнее.
Гипотезы о механизмах функционирования

Слайд 21(Harrison et al, 2005)
CrKNOX1 mRNA
SkKNOX1 mRNA
Zamia floridans
(from Bharathan et
(Harrison et al, 2005)
CrKNOX1 mRNA
SkKNOX1 mRNA
Zamia floridans
(from Bharathan et
Слайд 22Отсутствуют данные о генетической регуляции в меристеме симплексного типа плауноообразных.
Нами был
Отсутствуют данные о генетической регуляции в меристеме симплексного типа плауноообразных.
Нами был

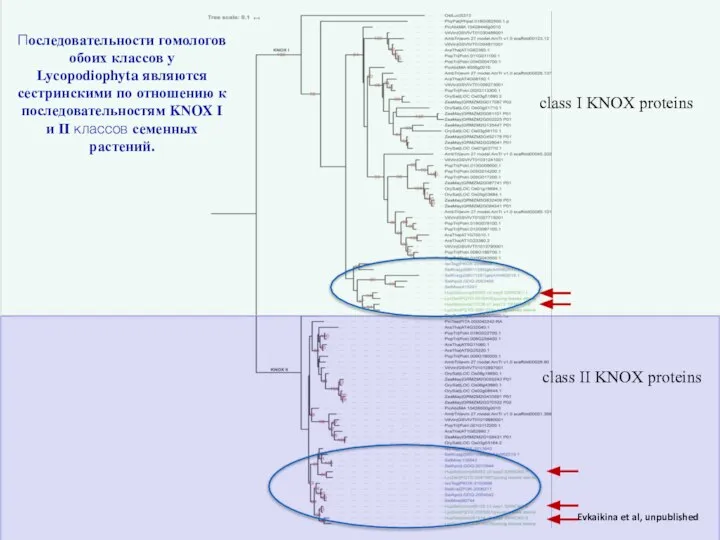
Слайд 23Evkaikina et al, unpublished
Последовательности гомологов обоих классов у Lycopodiophyta являются сестринскими по
Evkaikina et al, unpublished
Последовательности гомологов обоих классов у Lycopodiophyta являются сестринскими по
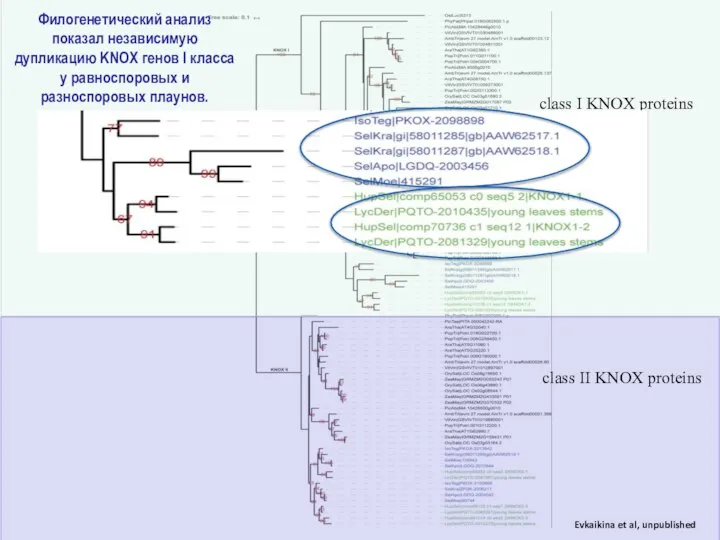
Слайд 24Филогенетический анализ показал независимую дупликацию KNOX генов I класса у равноспоровых и
Филогенетический анализ показал независимую дупликацию KNOX генов I класса у равноспоровых и
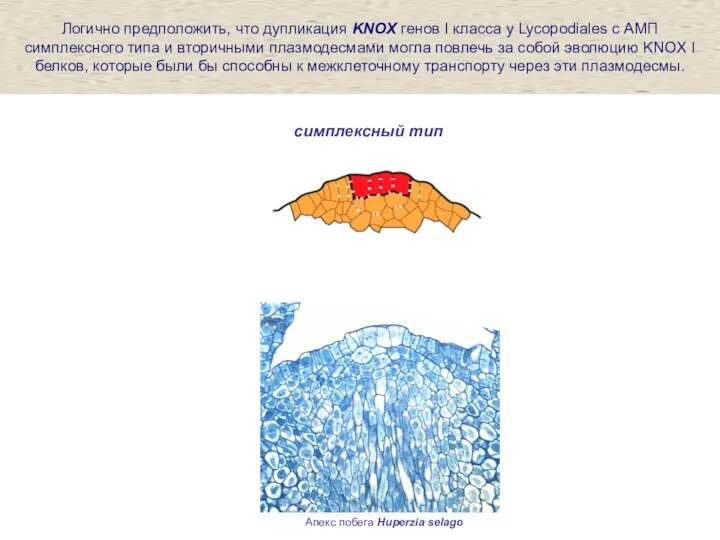
Слайд 25Апекс побега Huperzia selago
симплексный тип
Логично предположить, что дупликация KNOX генов I класса
Апекс побега Huperzia selago
симплексный тип
Логично предположить, что дупликация KNOX генов I класса
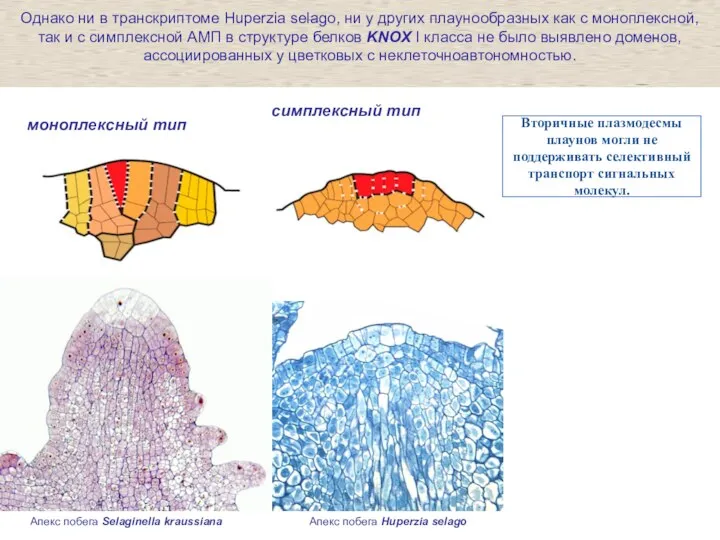
Слайд 26Апекс побега Selaginella kraussiana
Апекс побега Huperzia selago
моноплексный тип
симплексный тип
Однако ни в
Апекс побега Selaginella kraussiana
Апекс побега Huperzia selago
моноплексный тип
симплексный тип
Однако ни в
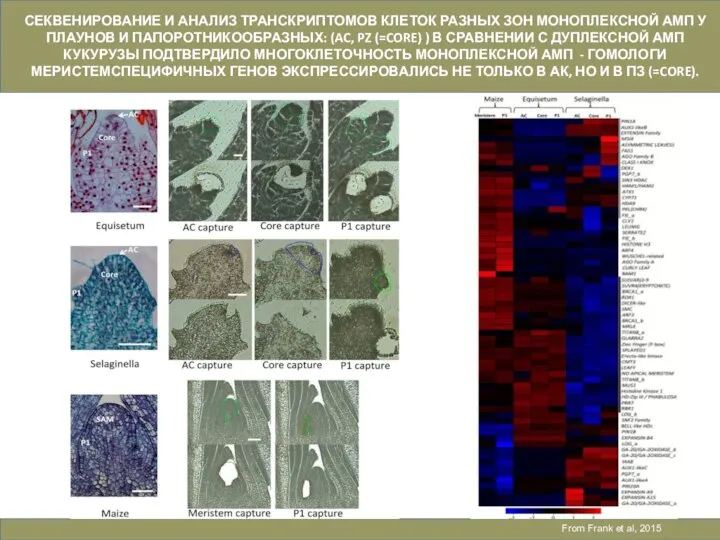
Слайд 27From Frank et al, 2015
СЕКВЕНИРОВАНИЕ И АНАЛИЗ ТРАНСКРИПТОМОВ КЛЕТОК РАЗНЫХ ЗОН МОНОПЛЕКСНОЙ
From Frank et al, 2015
СЕКВЕНИРОВАНИЕ И АНАЛИЗ ТРАНСКРИПТОМОВ КЛЕТОК РАЗНЫХ ЗОН МОНОПЛЕКСНОЙ
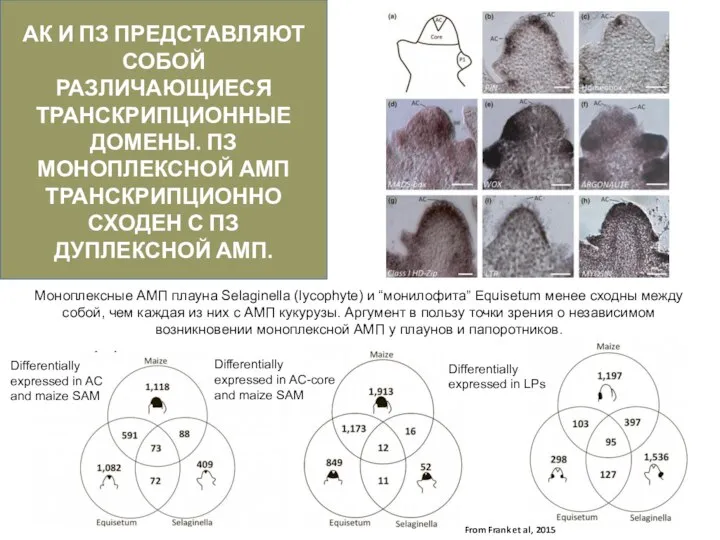
Слайд 28АК И ПЗ ПРЕДСТАВЛЯЮТ СОБОЙ РАЗЛИЧАЮЩИЕСЯ ТРАНСКРИПЦИОННЫЕ ДОМЕНЫ. ПЗ МОНОПЛЕКСНОЙ АМП ТРАНСКРИПЦИОННО
АК И ПЗ ПРЕДСТАВЛЯЮТ СОБОЙ РАЗЛИЧАЮЩИЕСЯ ТРАНСКРИПЦИОННЫЕ ДОМЕНЫ. ПЗ МОНОПЛЕКСНОЙ АМП ТРАНСКРИПЦИОННО
Слайд 29ГОМОЛОГИ СLV3-LIKE (CLE) ГЕНОВ.
monoplex SAM
simplex SAM
duplex SAM
Expression of CLE209 and CLE210 in
ГОМОЛОГИ СLV3-LIKE (CLE) ГЕНОВ.
monoplex SAM
simplex SAM
duplex SAM
Expression of CLE209 and CLE210 in

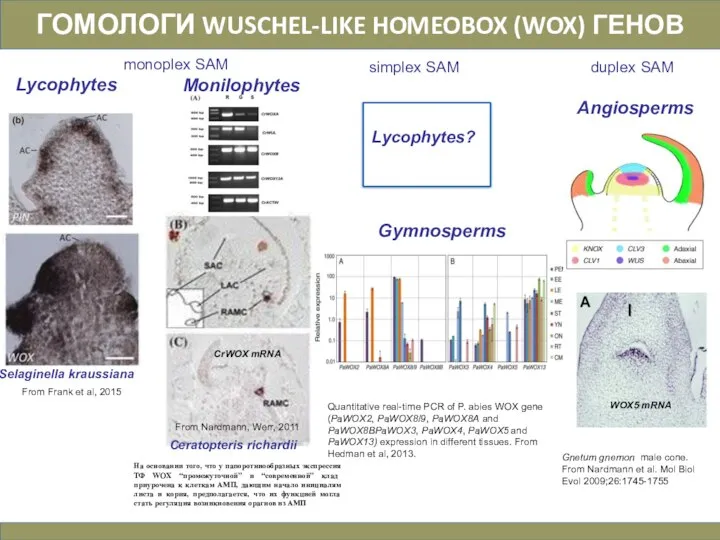
Слайд 30ГОМОЛОГИ WUSCHEL-LIKE HOMEOBOX (WOX) ГЕНОВ
monoplex SAM
simplex SAM
duplex SAM
Quantitative real-time PCR of P.
ГОМОЛОГИ WUSCHEL-LIKE HOMEOBOX (WOX) ГЕНОВ
monoplex SAM
simplex SAM
duplex SAM
Quantitative real-time PCR of P.
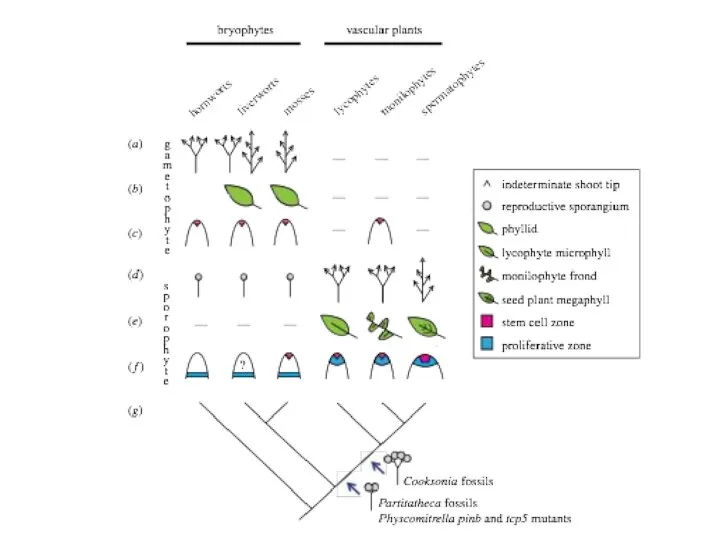
Слайд 31Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений
Листья
Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений
Листья
Слайд 32Huperzia selago
ЦЗ
ПЗ
ПЗ
АИ
Один из критериев энационного листа - возникновение ниже апикальной меристемы.
Huperzia selago
ЦЗ
ПЗ
ПЗ
АИ
Один из критериев энационного листа - возникновение ниже апикальной меристемы.

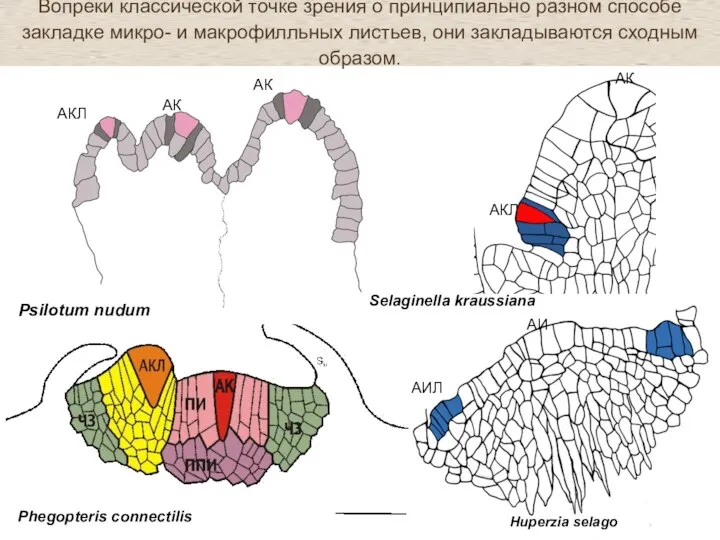
Слайд 33Вопреки классической точке зрения о принципиально разном способе закладке микро- и макрофилльных
Вопреки классической точке зрения о принципиально разном способе закладке микро- и макрофилльных
Слайд 34Гаплостела Риниевые
Актиностела Псилотовые, плауны
Эвстела Голосеменные, Двудольные
Атактостела
Однодольные
Диктиостела
Папоротники
Плектостела Плауны
Артростела
Хвощи
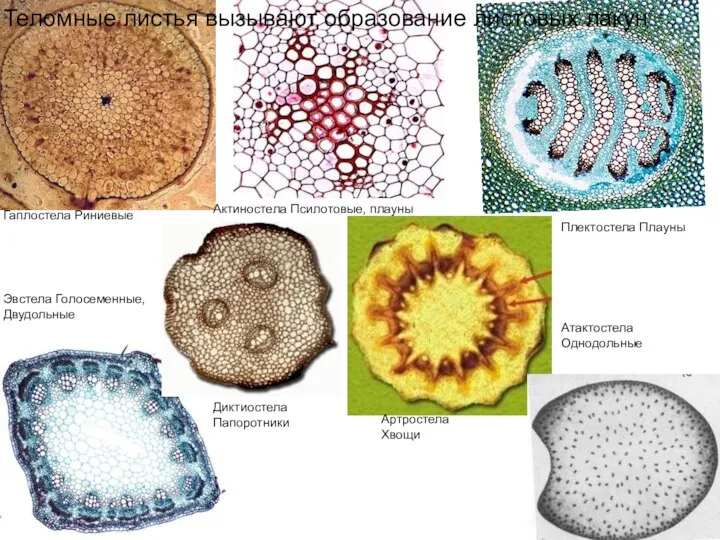
Теломные листья вызывают образование
Гаплостела Риниевые
Актиностела Псилотовые, плауны
Эвстела Голосеменные, Двудольные
Атактостела
Однодольные
Диктиостела
Папоротники
Плектостела Плауны
Артростела
Хвощи
Теломные листья вызывают образование
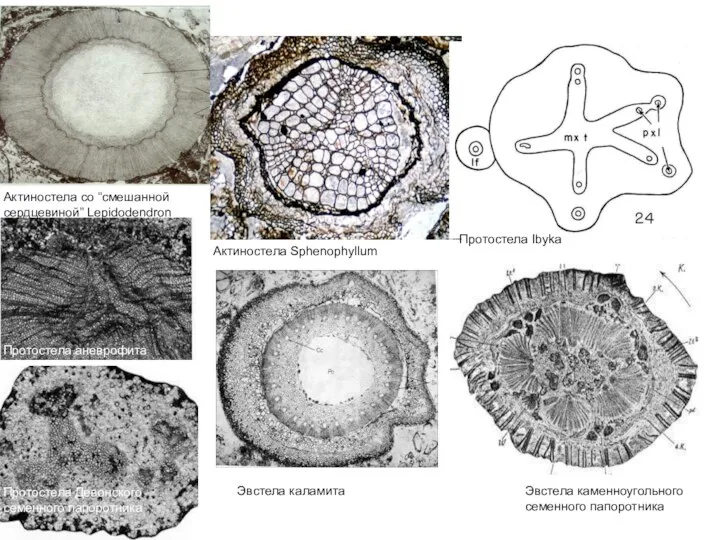
Слайд 35Актиностела Sphenophyllum
Актиностела со “смешанной сердцевиной” Lepidodendron
Эвстела каменноугольного семенного папоротника
Протостела аневрофита
Протостела Девонского семенного
Актиностела Sphenophyllum
Актиностела со “смешанной сердцевиной” Lepidodendron
Эвстела каменноугольного семенного папоротника
Протостела аневрофита
Протостела Девонского семенного
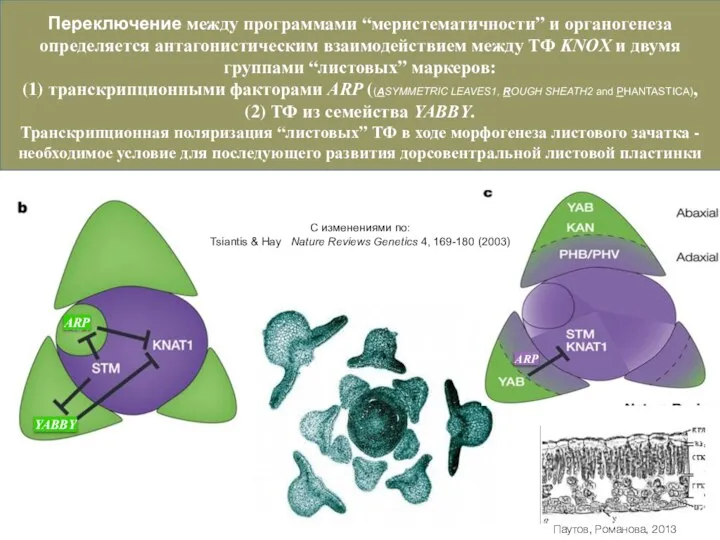
Слайд 36Переключение между программами “меристематичности” и органогенеза определяется антагонистическим взаимодействием между ТФ KNOX
Переключение между программами “меристематичности” и органогенеза определяется антагонистическим взаимодействием между ТФ KNOX
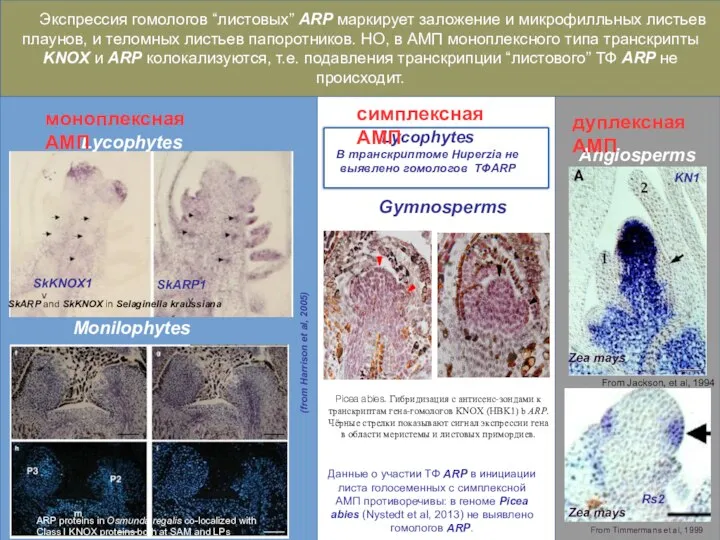
Слайд 37SkARP1
SkKNOX1
SkARP and SkKNOX in Selaginella kraussiana
ARP proteins in Osmunda regalis co-localized with
SkARP1
SkKNOX1
SkARP and SkKNOX in Selaginella kraussiana
ARP proteins in Osmunda regalis co-localized with
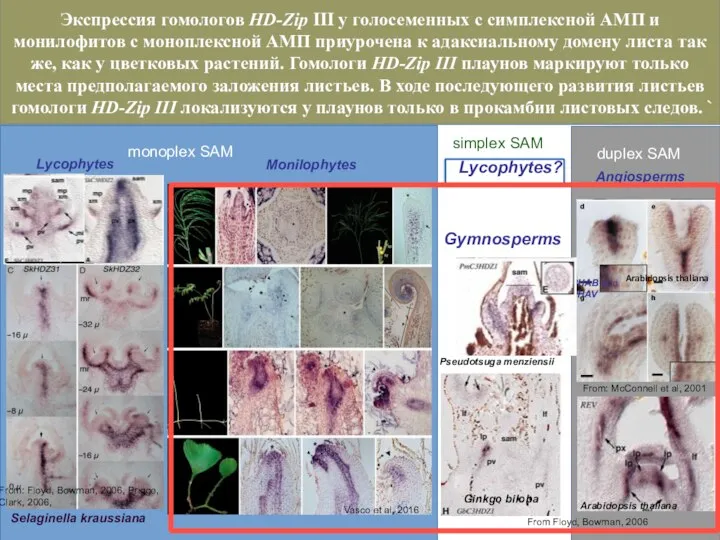
Слайд 38monoplex SAM
simplex SAM
duplex SAM
PHAB and PHAV
From Floyd, Bowman, 2006
Pseudotsuga menziensii
Ginkgo biloba
Selaginella kraussiana
From:
monoplex SAM
simplex SAM
duplex SAM
PHAB and PHAV
From Floyd, Bowman, 2006
Pseudotsuga menziensii
Ginkgo biloba
Selaginella kraussiana
From:
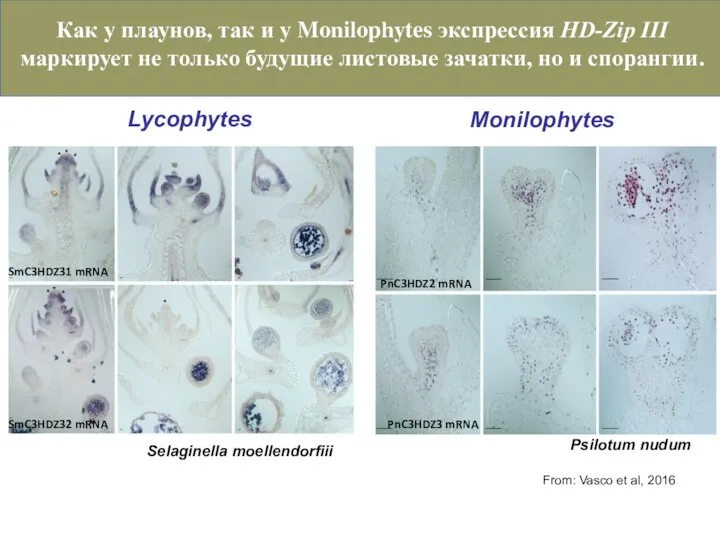
Слайд 39Selaginella moellendorfiii
SmC3HDZ32 mRNA
SmC3HDZ31 mRNA
Psilotum nudum
PnC3HDZ2 mRNA
From: Vasco et al, 2016
PnC3HDZ3 mRNA
Lycophytes
Monilophytes
Как у
Selaginella moellendorfiii
SmC3HDZ32 mRNA
SmC3HDZ31 mRNA
Psilotum nudum
PnC3HDZ2 mRNA
From: Vasco et al, 2016
PnC3HDZ3 mRNA
Lycophytes
Monilophytes
Как у
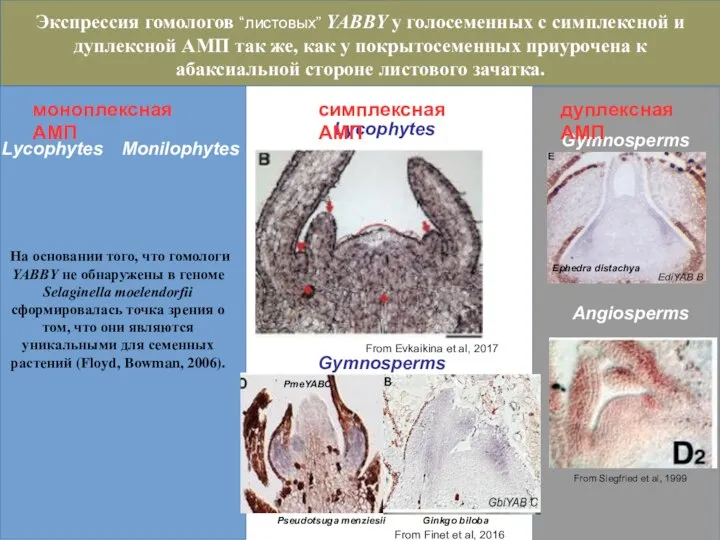
Слайд 40Lycophytes
Gymnosperms
Monilophytes
From Finet et al, 2016
PmeYABC
Экспрессия гомологов “листовых” YABBY у голосеменных с симплексной
Lycophytes
Gymnosperms
Monilophytes
From Finet et al, 2016
PmeYABC
Экспрессия гомологов “листовых” YABBY у голосеменных с симплексной
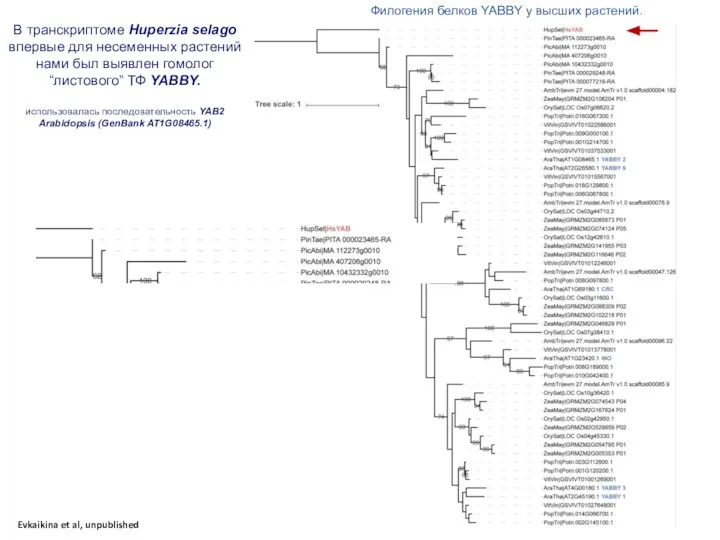
Слайд 41Филогения белков YABBY у высших растений.
В транскриптоме Huperzia selago впервые для несеменных
Филогения белков YABBY у высших растений.
В транскриптоме Huperzia selago впервые для несеменных
Слайд 42Имеющиеся на настоящий момент молекулярно-генетические данные показали, что функциональная регуляция АМП в

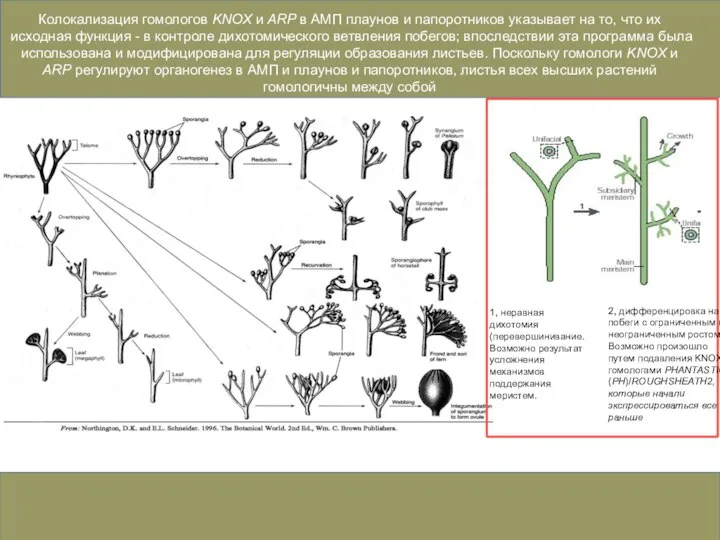
Слайд 43Колокализация гомологов KNOX и ARP в АМП плаунов и папоротников указывает на
Колокализация гомологов KNOX и ARP в АМП плаунов и папоротников указывает на
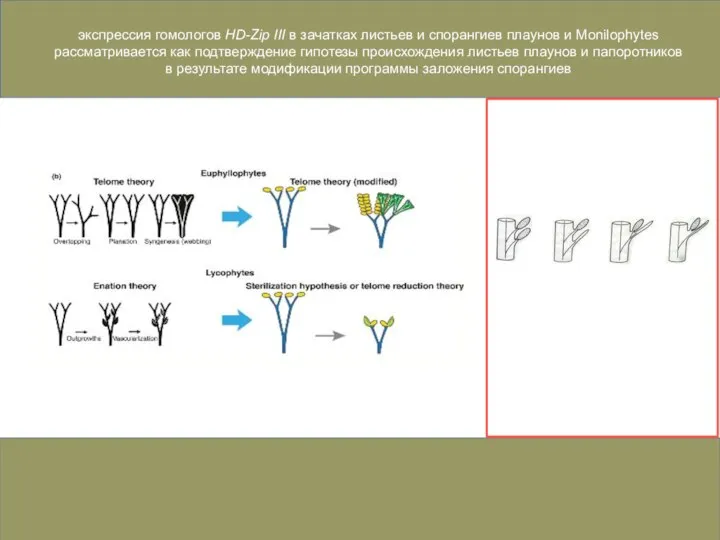
Слайд 44экспрессия гомологов HD-Zip III в зачатках листьев и спорангиев плаунов и Monilophytes
экспрессия гомологов HD-Zip III в зачатках листьев и спорангиев плаунов и Monilophytes
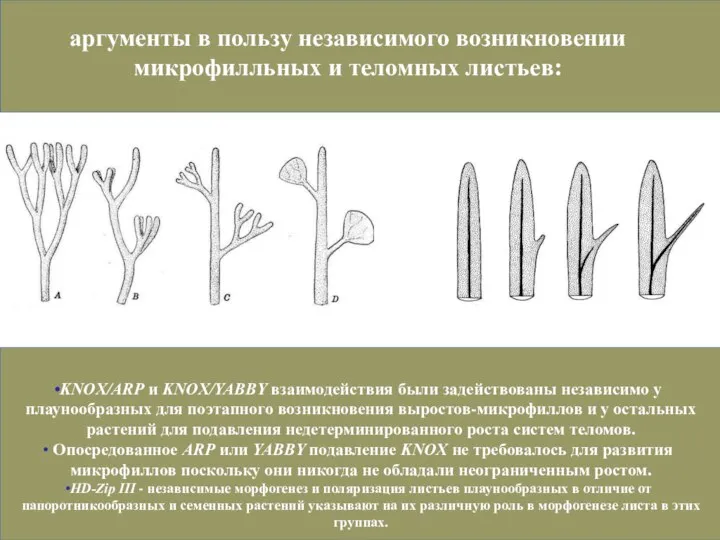
Слайд 45KNOX/ARP и KNOX/YABBY взаимодействия были задействованы независимо у плаунообразных для поэтапного возникновения
Слайд 46From: A. Tomescu Trends in Plant Science. 2009
различия в экспрессии KNOX у
From: A. Tomescu Trends in Plant Science. 2009
различия в экспрессии KNOX у

Слайд 47Наличие в транскриптоме Huperzia гомологов ТФ YABBY, а у Selaginella ARP, позволяет
Наличие в транскриптоме Huperzia гомологов ТФ YABBY, а у Selaginella ARP, позволяет
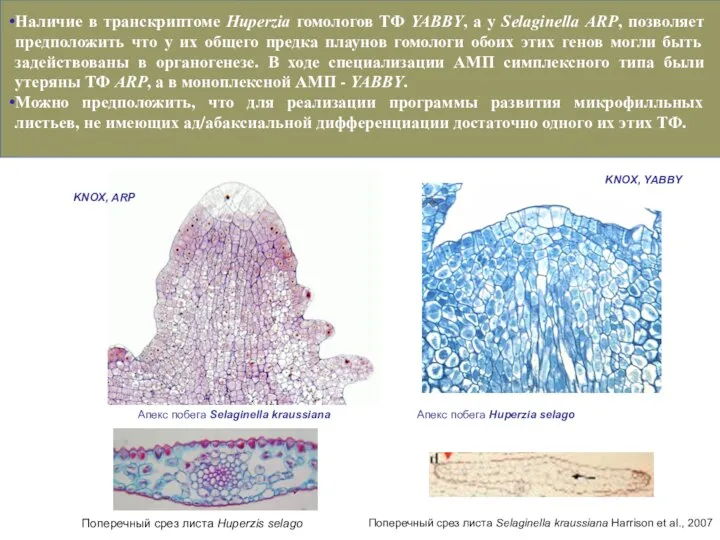
Слайд 481, неравная дихотомия (перевершинивание. Возможно результат усложнения механизмов поддержания меристем.
2, дифференцировка на
1, неравная дихотомия (перевершинивание. Возможно результат усложнения механизмов поддержания меристем.
2, дифференцировка на
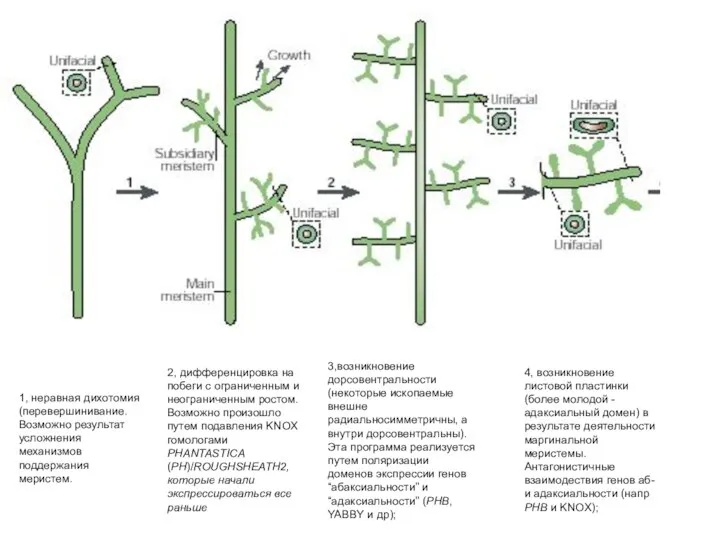
Слайд 495, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов
5, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов
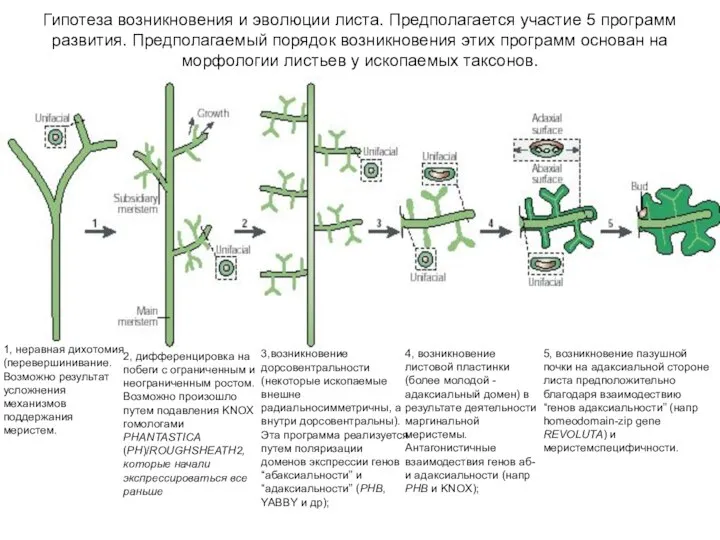
Слайд 50КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА: У ПРЕДСТАВИТЕЛЕЙ ТАКСОНОВ С МИКРОФИЛЛЬНЫМИ
КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА: У ПРЕДСТАВИТЕЛЕЙ ТАКСОНОВ С МИКРОФИЛЛЬНЫМИ
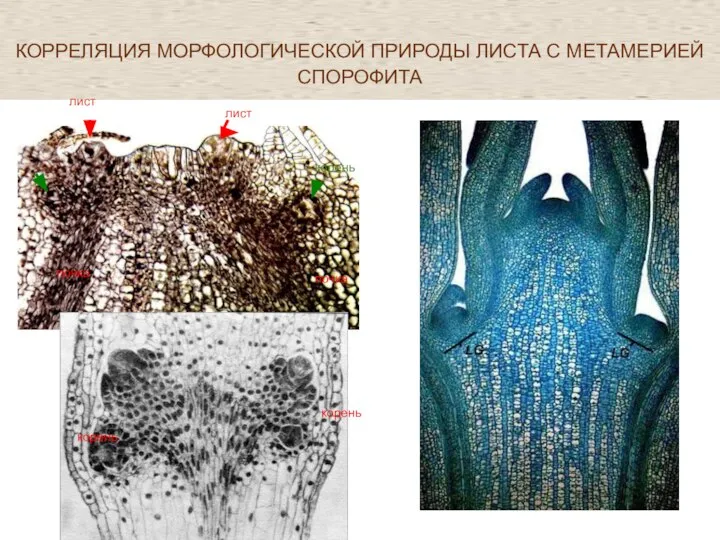
Слайд 51КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА
Phegopteris connectilis
лист
лист
корень
почка
почка
корень
корень
КОРРЕЛЯЦИЯ МОРФОЛОГИЧЕСКОЙ ПРИРОДЫ ЛИСТА С МЕТАМЕРИЕЙ СПОРОФИТА
Phegopteris connectilis
лист
лист
корень
почка
почка
корень
корень
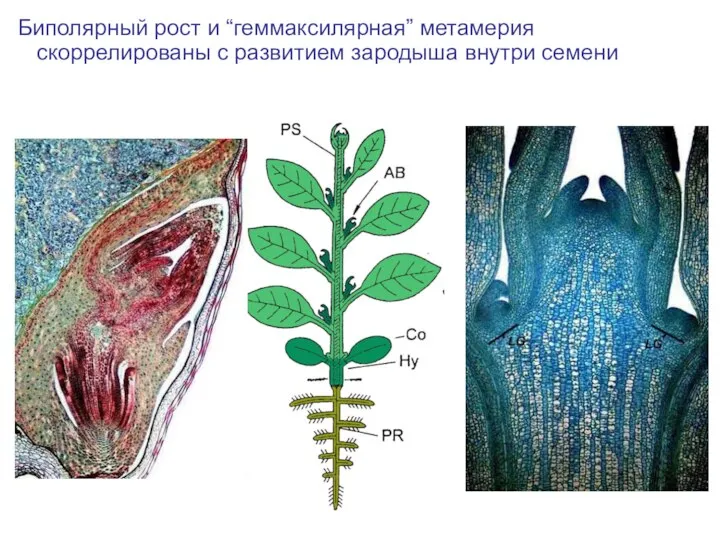
Слайд 52Биполярный рост и “геммаксилярная” метамерия скоррелированы с развитием зародыша внутри семени
Биполярный рост и “геммаксилярная” метамерия скоррелированы с развитием зародыша внутри семени
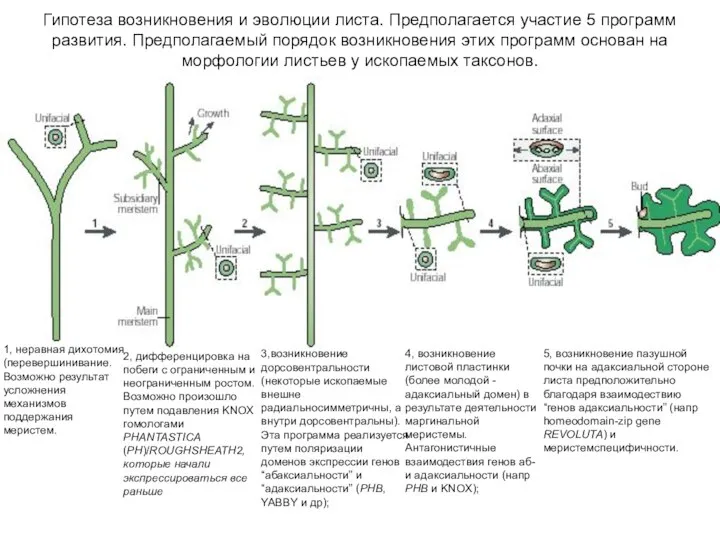
Слайд 535, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов
5, возникновение пазушной почки на адаксиальной стороне листа предположительно благодаря взаимодествию “генов
Слайд 54Элементарный метамер семенных растений - лист и пазушая почка
ОРГАНОГЕНЕЗ Наименьшая повторяющаяся структурная
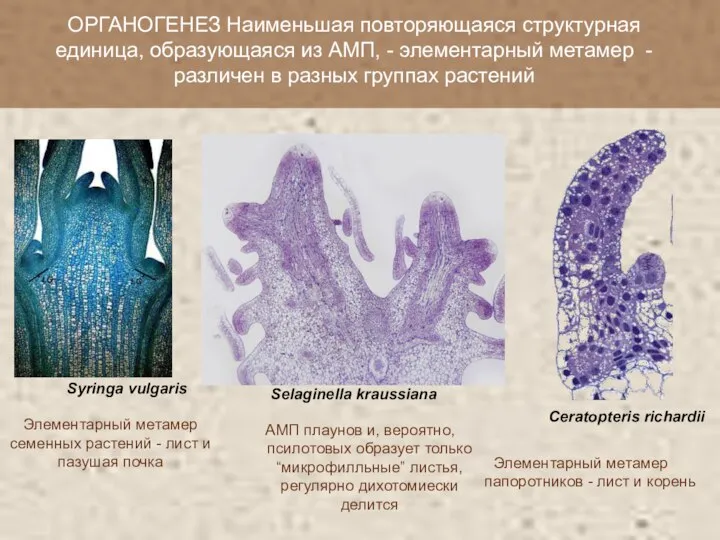
Элементарный метамер семенных растений - лист и пазушая почка
ОРГАНОГЕНЕЗ Наименьшая повторяющаяся структурная

 Клён
Клён Царство грибов
Царство грибов Презентация на тему Абиотические факторы среды и их влияние на живые организмы
Презентация на тему Абиотические факторы среды и их влияние на живые организмы  Клеточная инженерия
Клеточная инженерия Презентация на тему ЖИЗНЕННЫЕ ФОРМЫ РАСТЕНИЙ
Презентация на тему ЖИЗНЕННЫЕ ФОРМЫ РАСТЕНИЙ  Презентация на тему Приспособленность организмов – результат эволюции
Презентация на тему Приспособленность организмов – результат эволюции  Крушина ольховидная. Семейство Крушиновые
Крушина ольховидная. Семейство Крушиновые Пищеварительная система
Пищеварительная система Бионика. Технический взгляд на природу
Бионика. Технический взгляд на природу Презентация на тему Селекция (11 класс)
Презентация на тему Селекция (11 класс)  Копытные. Дикий северный олень
Копытные. Дикий северный олень Видообразование
Видообразование «ПЕРЕЛЁТНЫЕ ПТИЦЫ»
«ПЕРЕЛЁТНЫЕ ПТИЦЫ» Биология в естествознании
Биология в естествознании Виды изменчивости. Модификационная изменчивость
Виды изменчивости. Модификационная изменчивость Демэкология экология популяций
Демэкология экология популяций Роль кожи в терморегуляции организма
Роль кожи в терморегуляции организма Карл Лінней (1707-1778)
Карл Лінней (1707-1778) Спинной мозг
Спинной мозг Подражание Форма научения
Подражание Форма научения Coraux d’Hawaïï
Coraux d’Hawaïï Жизненные циклы паразитов
Жизненные циклы паразитов Приспособительное поведение
Приспособительное поведение Обеспечение клеток энергией. Лекция №7
Обеспечение клеток энергией. Лекция №7 Ассортимент плодовых и ягодных культур для Удмуртии. Современные перспективные сорта
Ассортимент плодовых и ягодных культур для Удмуртии. Современные перспективные сорта Основной обмен. Минимальная скорость расходования энергии, необходимая для поддержания жизни. Факторы среды и рост
Основной обмен. Минимальная скорость расходования энергии, необходимая для поддержания жизни. Факторы среды и рост Мышечные ткани
Мышечные ткани Экология паразитов
Экология паразитов