Эволюция головного мозга у позвоночных. Смена типов головного мозга. Формирование первичной и вторичной коры
- Эволюция головного мозга у позвоночных. Смена типов головного мозга. Формирование первичной и вторичной коры

Содержание
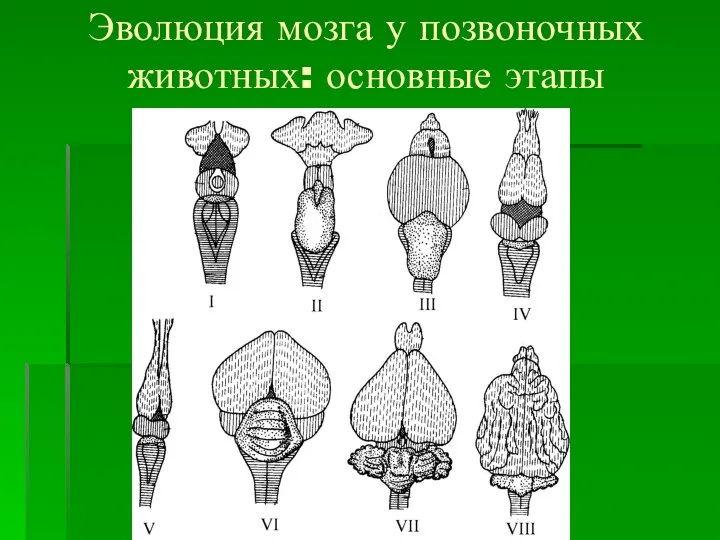
- 2. Эволюция мозга у позвоночных животных: основные этапы
- 3. Этап 1. Образование центральной нервной системы в виде нервной трубки впервые появляется у животных представителей типа
- 4. Этап 2. У позвоночных животных нервная трубка подразделяется на головной и спинной мозг. В эмбриональной стадии
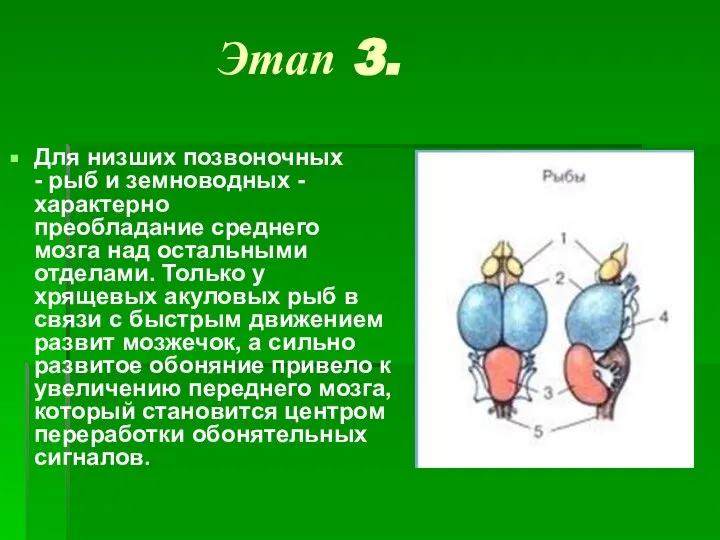
- 5. Этап 3. Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными
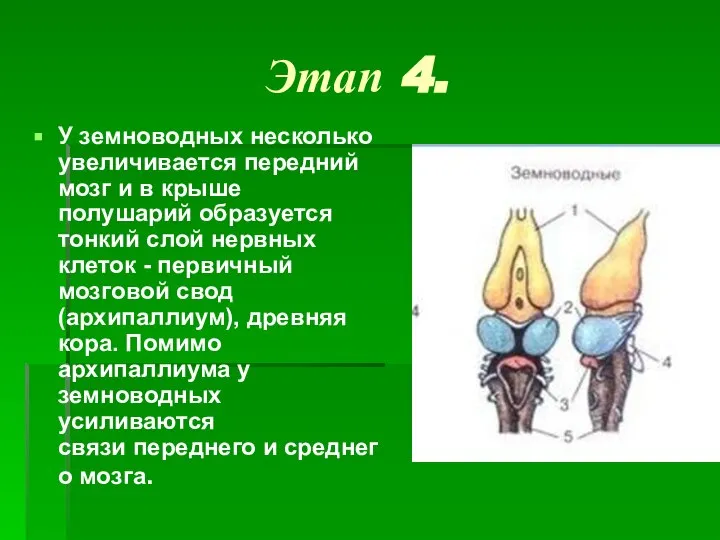
- 6. Этап 4. У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных
- 7. Этап 5. У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток - полосатых тел
- 8. В строении головного мозга птиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора,
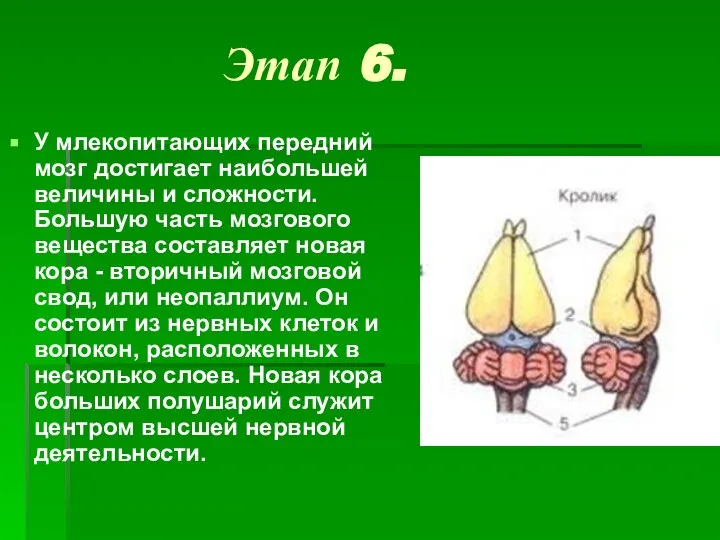
- 9. Этап 6. У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового вещества составляет
- 10. Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают
- 11. Этап 7. Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа. Мозг как
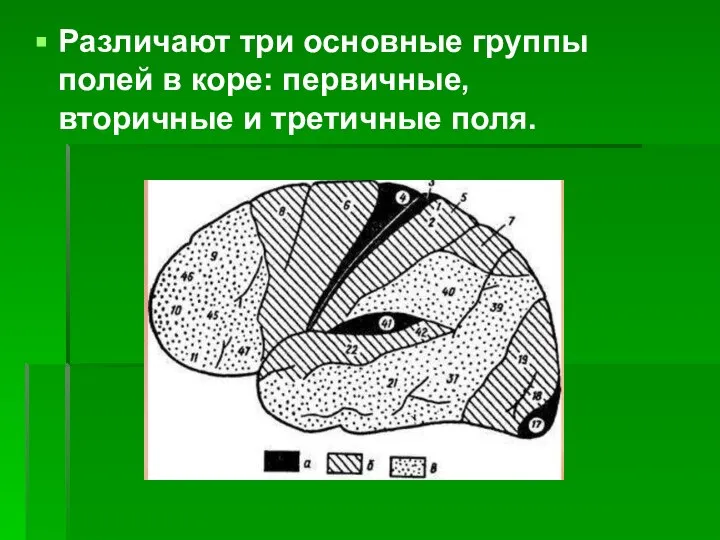
- 12. Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
- 13. Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в
- 14. Величина зоны представительства того или иного рецепторного участка в первичной коре зависит от функциональной значимости этого
- 15. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные
- 16. Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть
- 17. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь
- 18. При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и
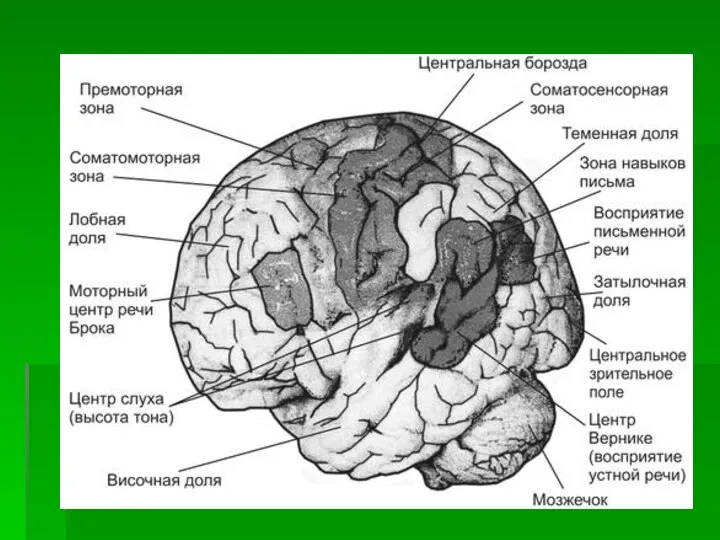
- 20. Различия в функциональной специфике первичных, вторичных и третичных полей обусловливают и различия в их способности компенсировать
- 21. Функции пострадавших третичных полей компенсируются относительно легко за счет полимодальности, позволяющей опираться на мощную систему ассоциаций,
- 23. Скачать презентацию
Слайд 3Этап 1.
Образование центральной нервной системы в виде нервной трубки впервые появляется у животных представителей
Этап 1.
Образование центральной нервной системы в виде нервной трубки впервые появляется у животных представителей

Слайд 4Этап 2.
У позвоночных животных нервная трубка подразделяется на головной и спинной мозг. В эмбриональной
Этап 2.
У позвоночных животных нервная трубка подразделяется на головной и спинной мозг. В эмбриональной

Слайд 5Этап 3.
Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами.
Этап 3.
Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами.
Слайд 6Этап 4.
У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных
Этап 4.
У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных
Слайд 7Этап 5.
У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток - полосатых тел -
Этап 5.
У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток - полосатых тел -

Слайд 8В строении головного мозга птиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо
В строении головного мозга птиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо

Слайд 9Этап 6.
У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового
Этап 6.
У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового
Слайд 10Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под
Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под

Слайд 11Этап 7.
Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа.
Этап 7.
Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа.

Слайд 12Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Слайд 13Первичные поля связаны с органами чувств и органами движения на периферии, они
Первичные поля связаны с органами чувств и органами движения на периферии, они

Слайд 14Величина зоны представительства того или иного рецепторного участка в первичной коре зависит
Величина зоны представительства того или иного рецепторного участка в первичной коре зависит

Слайд 15Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными
Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными

Слайд 16Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия

Слайд 17Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее
Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее

Слайд 18При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит

Слайд 20Различия в функциональной специфике первичных, вторичных и третичных полей обусловливают и различия
Различия в функциональной специфике первичных, вторичных и третичных полей обусловливают и различия

Слайд 21Функции пострадавших третичных полей компенсируются относительно легко за счет полимодальности, позволяющей опираться
Функции пострадавших третичных полей компенсируются относительно легко за счет полимодальности, позволяющей опираться

 Экологические сообщества
Экологические сообщества Многообразие животного мира нашей планеты
Многообразие животного мира нашей планеты Насекомые. Старшая группа (5 – 6 лет)
Насекомые. Старшая группа (5 – 6 лет) кишечнополостные
кишечнополостные Вітамін В12
Вітамін В12 Осень, Осень, в гости просим!
Осень, Осень, в гости просим! Кто такие неандертальцы?
Кто такие неандертальцы? Роль химии в ветеринарии
Роль химии в ветеринарии Экологические факторы среды (5 класс)
Экологические факторы среды (5 класс) Проверка антибактериальных свойств лука и чеснока на бактериях слизистой оболочки ротовой полости
Проверка антибактериальных свойств лука и чеснока на бактериях слизистой оболочки ротовой полости Строение и функции листа
Строение и функции листа Аксолотль
Аксолотль Химический состав клетки
Химический состав клетки Презентация на тему Организменная среда обитания
Презентация на тему Организменная среда обитания  Энциклопедия леса. Деревья
Энциклопедия леса. Деревья Презентация на тему АДАПТАЦИЯ ОРГАНИЗМОВ К УСЛОВИЯМ ОБИТАНИЯ КАК РЕЗУЛЬТАТ ДЕЙСТВИЯ ЕСТЕСТВЕННОГО ОТБОРА
Презентация на тему АДАПТАЦИЯ ОРГАНИЗМОВ К УСЛОВИЯМ ОБИТАНИЯ КАК РЕЗУЛЬТАТ ДЕЙСТВИЯ ЕСТЕСТВЕННОГО ОТБОРА  Походная еда
Походная еда Белки острой фазы
Белки острой фазы Презентация на тему Патогенные бактерии
Презентация на тему Патогенные бактерии  Презентация на тему СТРОЕНИЕ СЕМЯН
Презентация на тему СТРОЕНИЕ СЕМЯН  Растительный покров земли
Растительный покров земли Кости и топография костей черепа. Виды соединения костей черепа
Кости и топография костей черепа. Виды соединения костей черепа Происхождение жизни на Земле
Происхождение жизни на Земле Общая характеристика физических способностей
Общая характеристика физических способностей Лента времени зарубежных социальных психологов. Ричард Нисбетт
Лента времени зарубежных социальных психологов. Ричард Нисбетт Презентация на тему Многообразие и значение земноводных
Презентация на тему Многообразие и значение земноводных  Бактерии: 1 Цианобактерии
Бактерии: 1 Цианобактерии Презентация на тему Эритроциты
Презентация на тему Эритроциты