- Ферменты

Содержание
- 2. ФЕРМЕНТЫ – БИОЛОГИЧЕСКИЕ КАТАЛИЗАТОРЫ БЕЛКОВОЙ ПРИРОДЫ. Термин «фермент» введен в начале 17 века Ван Гельмонтом. Синонимом
- 3. Fermentum – закваска En zyme – в дрожжах
- 4. Начало XVII века – Ван Гельмонт вводит название «фермент», как вещество влияющее на спиртовое брожение Конец
- 5. конец XIX века – М.М. Манассеина и братья Бухнер доказали несостоятельность деления ферментов на «организованные» и
- 6. 1955г. – Мур и Стейн полностью расшифровали, а в 1969г. В лаборатории Мерифилда искусственно синтезировали фермент
- 7. Основная биологическая роль ферментов – это обеспечение протекания химических реакций в живом организме. Вместе с тем
- 8. высокая скорость катализа обладают высокой специфичностью катализируют в мягких условиях (t=37о, рН-нейтральное, нормальное атмосферное давление) регулируемость
- 9. Тем не менее, ферменты подчиняются общим законам катализа: т.е.: Катализируют только энергетически возможные реакции. Они никогда
- 10. ХИМИЧЕСКАЯ ПРИРОДА ФЕРМЕНТОВ И ИХ СТРОЕНИЕ Все ферменты являются белками. Выделяют ферменты простые и сложные. Простые
- 11. Сложный фермент = Белковая часть + Небелковая (холофермент) (апофермент) часть (кофермент) Если кофермент очень прочно связан
- 12. До 1/3 всех ферментов нуждается в присутствии иона металла, который либо активирует фермент, либо обеспечивает его
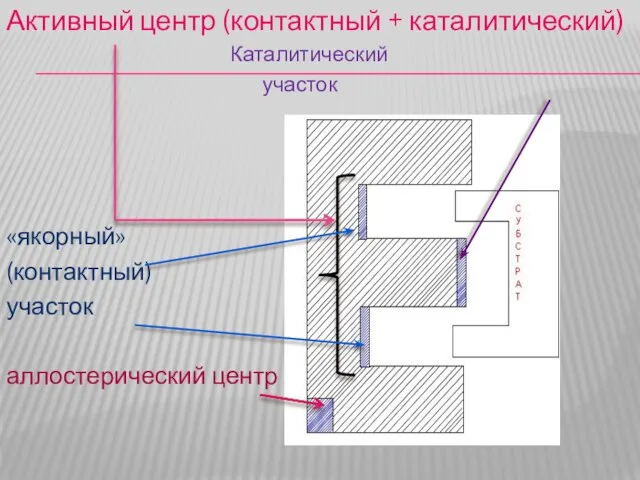
- 13. СТРОЕНИЕ МОЛЕКУЛЫ ФЕРМЕНТА Любой фермент имеет в своем составе активный центр. В состав активного центра входят
- 14. Активный центр (контактный + каталитический) Каталитический участок «якорный» (контактный) участок аллостерический центр
- 15. В состав активного центра сложного фермента входит кофермент или простетическая группа, в простом ферменте эту функцию
- 16. Аминокислоты, образующие активный центр в простом ферменте при образовании первичной структуры полипептидной цепи не обязательно находятся
- 17. Пример: активный центр пищеварительной гидролазы химотрипсина образуют серин - 195, гистидин – 57, аспарагиновая кислота –
- 18. АЛЛОСТЕРИЧЕСКИЙ ЦЕНТР Выполняет регуляторную функцию, путем взаимодействия с аллостерическим эффектором, веществом комплиментарным по строению с аллостерическим
- 19. Эффектор присоединяется в аллостерическом центре Изменяется конформация аллостерического центра Изменяется конформация фермента Изменяется конформация активного центра
- 20. ЭТАПЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА Диффузия фермента и субстрата; Образование фермент-субстратного комплекса за счет комплиментарности активного центра фермента
- 21. 4. Образование новых химических связей в молекулах, превращаемых под действием фермента и появление комплекса фермент +
- 22. E + S ES ES* EP E+P
- 23. МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ В основе действия любого фермента лежит снижение энергии активации. Данный эффект может быть
- 24. 2. Деформация субстрата. Заключается в принудительном увеличении межатомных связей субстрата, что позволяет легче их атаковать реагентом.
- 25. 4. Ковалентный катализ. Заключается в образовании с каталитическими группами фермента промежуточных веществ, которые затем легко распадаются
- 26. СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ В зависимости от степени сродства фермента к субстрату выделяют следующие виды специфичности: 1. Стереохимическая
- 27. 2. Абсолютная субстратная специфичность. Фермент катализирует превращение только одного субстрата. Пример: уреаза катализирует превращение только мочевины.
- 28. 4. Относительная групповая специфичность. Для фермента первостепенное значение имеет характер химической связи в субстрате. Пример: протеазы
- 29. ТЕОРИИ ОБЪЯСНЯЮШИЕ СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ Теория Фишера – «ключ-замок». Активный центр фермента жесткая структура. Субстрат подходит к
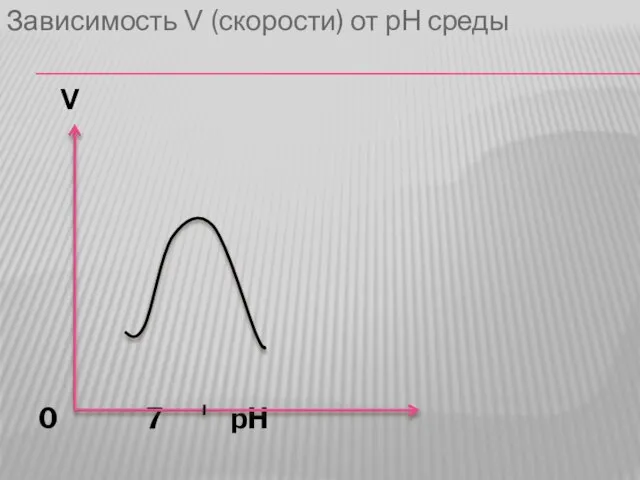
- 30. КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ 1. Зависимость скорости ферментативной реакции от pH. График имеет колоколообразный характер, с оптимумом
- 31. Зависимость V (скорости) от рН среды V 0 7 pH
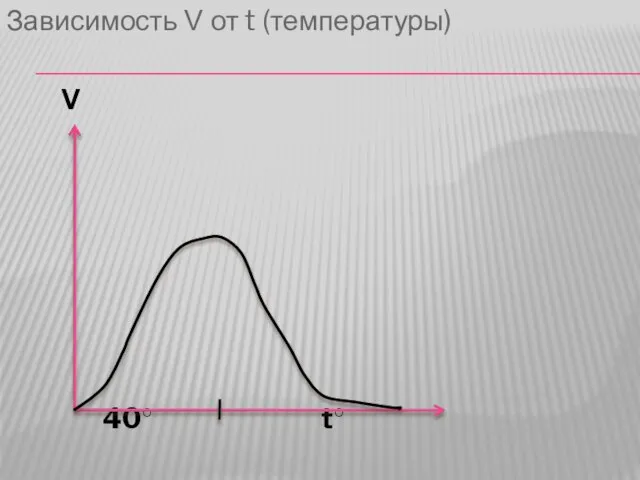
- 32. 2. Зависимость скорости ферментативной реакции от температуры. График имеет колоколообразный характер. При увеличении t в соответствии
- 33. Однако при дальнейшем увеличении температуры происходит денатурация ферментов и резкое падение скорости реакции, поэтому температуру тела
- 34. Зависимость V от t (температуры) V 40○ t○
- 35. Не все ферменты имеют оптимум при 40 °С. Например каталаза имеет оптимум при 0°С. 3. Зависимость
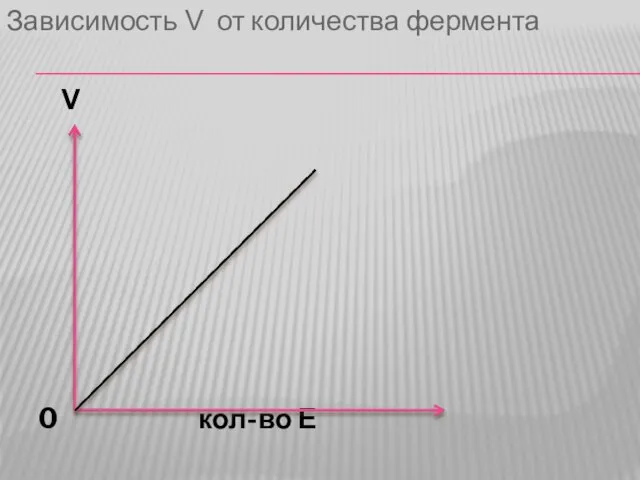
- 36. Зависимость V от количества фермента V 0 кол-во Е
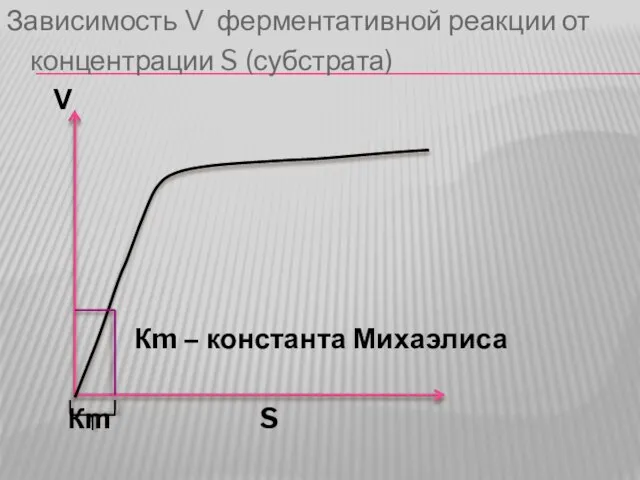
- 37. 4. Зависимость скорости ферментативной реакции от концентрации субстрата. Графическое изображение данной зависимости носит название «кривой Михаэлиса»
- 38. Зависимость V ферментативной реакции от концентрации S (субстрата) V Кm – константа Михаэлиса Кm S
- 39. Достижение максимальной скорости реакции зависит от сродства фермента к субстрату. Для того, чтобы численно отобразить степень
- 40. Константа Михаэлиса – это такое количество субстрата, при котором достигается половина максимальной скорости реакции. Т.о., чем
- 41. Классификация ферментов Оксидоредуктазы – обеспечивают перенос электронов и протонов. Участие в окислительно – восстановительных реакциях. Трансферазы
- 42. ХАРАКТЕРИСТИКА КЛАССОВ ФЕРМЕНТОВ I класс. Оксидоредуктазы катализируют окисление-дегидрирование в аэробных и анаэробных условиях. Редуктазы – анаэробные
- 43. Коферменты оксидоредуктаз (1) Никотинамидные (НАД, НАДФ) Флавиновые (ФМН, ФАД) Металлопорфириновые (гемы a, b, c, d) Хинонкоферменты
- 44. II класс. Трансферазы – перенос атомных групп и молекулярных остатков. Подразделяется на 8 подклассов.
- 45. Группы: амино-, альдегидная, сульфо–, кетонные остатки, глюко–, фосфо–, одноуглеродые фрагменты (CH3-, СН2-) Пример: гексокиназа. D –
- 46. Коферменты трансфераз (2) Пиридоксиновые (ПАЛФ, ПАМФ) Пантотеновые (КоА, дефосфо – КоА, 4 - фосфопантотенат) Нуклеотидные (УДФ
- 47. III класс. Гидролазы – ферменты, которые осуществляют разрыв химической связи с присоединением молекулы воды. В основном
- 48. IV класс. Лиазы – осуществляют негидролитические и неокислительные реакции распада и обратные им реакции синтеза (синтазы).
- 49. Коферменты лиаз (4) Пиридоксиновые (ПАЛФ) Пантотеновые (КоА, дефосфо - КоА) Тиаминовые (ТДФ) Кобамидные (дезоксиаденозилкобаламин)
- 50. V класс. Изомеразы – осуществляют внутримолекулярные превращения, изменения пространственного положения, перенос Н, перемещение двойных связей. Класс
- 51. Коферменты изомераз (5) Пиридоксиновые (ПАЛФ) Кобамидные (дезоксиаденозилкобаламин) Фосфаты моносахаридов (глюкозо – 1, 6 – дифосфат, 2,
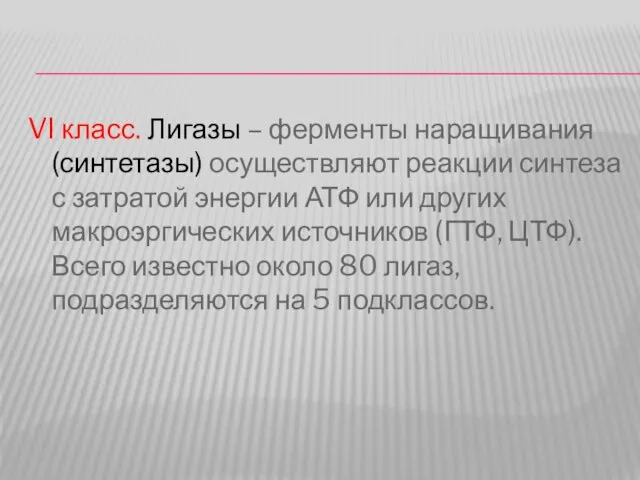
- 52. VI класс. Лигазы – ферменты наращивания (синтетазы) осуществляют реакции синтеза с затратой энергии АТФ или других
- 53. Коферменты лигаз (6) Нуклеотидные (УДФ – глюкоза, ЦДФ – холин и др.) Биотиновые (карбоксибиотин) Фолиевые (5,
- 54. НОМЕНКЛАТУРА ФЕРМЕНТОВ Для ряда давно известных ферментов оставлены тривиальные названия: пепсин, трипсин, амилаза. Кроме того все
- 55. Рабочее название: Лактат + дегидрогенизация + аза лактатдегидрогеназа Систематическое название: L – лактат: НАД+ - оксидоредуктаза
- 57. Скачать презентацию

Слайд 3Fermentum – закваска
En zyme – в дрожжах
En zyme – в дрожжах

Слайд 4Начало XVII века – Ван Гельмонт вводит название «фермент», как вещество влияющее
Начало XVII века – Ван Гельмонт вводит название «фермент», как вещество влияющее

Слайд 5конец XIX века – М.М. Манассеина и братья Бухнер доказали несостоятельность деления
конец XIX века – М.М. Манассеина и братья Бухнер доказали несостоятельность деления

Слайд 61955г. – Мур и Стейн полностью расшифровали, а в 1969г. В лаборатории
1955г. – Мур и Стейн полностью расшифровали, а в 1969г. В лаборатории

Слайд 7Основная биологическая роль ферментов – это обеспечение протекания химических реакций в живом
Основная биологическая роль ферментов – это обеспечение протекания химических реакций в живом

Слайд 8высокая скорость катализа
обладают высокой специфичностью
катализируют в мягких условиях (t=37о, рН-нейтральное, нормальное атмосферное
высокая скорость катализа
обладают высокой специфичностью
катализируют в мягких условиях (t=37о, рН-нейтральное, нормальное атмосферное

Слайд 9Тем не менее, ферменты подчиняются общим законам катализа: т.е.:
Катализируют только энергетически возможные
Тем не менее, ферменты подчиняются общим законам катализа: т.е.:
Катализируют только энергетически возможные

Слайд 10ХИМИЧЕСКАЯ ПРИРОДА ФЕРМЕНТОВ И ИХ СТРОЕНИЕ
Все ферменты являются белками. Выделяют ферменты простые
ХИМИЧЕСКАЯ ПРИРОДА ФЕРМЕНТОВ И ИХ СТРОЕНИЕ
Все ферменты являются белками. Выделяют ферменты простые

Слайд 11Сложный фермент = Белковая часть + Небелковая
(холофермент) (апофермент) часть
(кофермент)
Если кофермент очень
Сложный фермент = Белковая часть + Небелковая
(холофермент) (апофермент) часть
(кофермент)
Если кофермент очень

Слайд 12До 1/3 всех ферментов нуждается в присутствии иона металла, который либо активирует
До 1/3 всех ферментов нуждается в присутствии иона металла, который либо активирует

Слайд 13СТРОЕНИЕ МОЛЕКУЛЫ ФЕРМЕНТА
Любой фермент имеет в своем составе активный центр. В состав
СТРОЕНИЕ МОЛЕКУЛЫ ФЕРМЕНТА
Любой фермент имеет в своем составе активный центр. В состав

Слайд 14Активный центр (контактный + каталитический)
Каталитический
участок
«якорный»
(контактный)
участок
аллостерический центр
Активный центр (контактный + каталитический)
Каталитический
участок
«якорный»
(контактный)
участок
аллостерический центр
Слайд 15В состав активного центра сложного фермента входит кофермент или простетическая группа, в
В состав активного центра сложного фермента входит кофермент или простетическая группа, в

Слайд 16Аминокислоты, образующие активный центр в простом ферменте при образовании первичной структуры полипептидной
Аминокислоты, образующие активный центр в простом ферменте при образовании первичной структуры полипептидной

Слайд 17Пример: активный центр пищеварительной гидролазы химотрипсина образуют серин - 195, гистидин –
Пример: активный центр пищеварительной гидролазы химотрипсина образуют серин - 195, гистидин –

Слайд 18АЛЛОСТЕРИЧЕСКИЙ ЦЕНТР
Выполняет регуляторную функцию, путем взаимодействия с аллостерическим эффектором, веществом комплиментарным по
АЛЛОСТЕРИЧЕСКИЙ ЦЕНТР
Выполняет регуляторную функцию, путем взаимодействия с аллостерическим эффектором, веществом комплиментарным по

Слайд 19Эффектор присоединяется в аллостерическом центре
Изменяется конформация аллостерического центра
Изменяется конформация фермента
Изменяется конформация активного
Эффектор присоединяется в аллостерическом центре
Изменяется конформация аллостерического центра
Изменяется конформация фермента
Изменяется конформация активного

Слайд 20ЭТАПЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Диффузия фермента и субстрата;
Образование фермент-субстратного комплекса за счет комплиментарности активного
ЭТАПЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
Диффузия фермента и субстрата;
Образование фермент-субстратного комплекса за счет комплиментарности активного

Слайд 214. Образование новых химических связей в молекулах, превращаемых под действием фермента и
4. Образование новых химических связей в молекулах, превращаемых под действием фермента и

Слайд 22E + S ES ES* EP E+P

Слайд 23МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ
В основе действия любого фермента лежит снижение энергии активации. Данный
МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ
В основе действия любого фермента лежит снижение энергии активации. Данный

Слайд 242. Деформация субстрата. Заключается в принудительном увеличении межатомных связей субстрата, что позволяет
2. Деформация субстрата. Заключается в принудительном увеличении межатомных связей субстрата, что позволяет

Слайд 254. Ковалентный катализ. Заключается в образовании с каталитическими группами фермента промежуточных веществ,
4. Ковалентный катализ. Заключается в образовании с каталитическими группами фермента промежуточных веществ,

Слайд 26СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ
В зависимости от степени сродства фермента к субстрату выделяют следующие виды
СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ
В зависимости от степени сродства фермента к субстрату выделяют следующие виды

Слайд 272. Абсолютная субстратная специфичность. Фермент катализирует превращение только одного субстрата. Пример: уреаза
2. Абсолютная субстратная специфичность. Фермент катализирует превращение только одного субстрата. Пример: уреаза

Слайд 284. Относительная групповая специфичность. Для фермента первостепенное значение имеет характер химической связи
4. Относительная групповая специфичность. Для фермента первостепенное значение имеет характер химической связи

Слайд 29ТЕОРИИ ОБЪЯСНЯЮШИЕ СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ
Теория Фишера – «ключ-замок». Активный центр фермента жесткая структура.
ТЕОРИИ ОБЪЯСНЯЮШИЕ СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ
Теория Фишера – «ключ-замок». Активный центр фермента жесткая структура.

Слайд 30КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
1. Зависимость скорости ферментативной реакции от pH. График имеет колоколообразный
КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
1. Зависимость скорости ферментативной реакции от pH. График имеет колоколообразный

Слайд 31Зависимость V (скорости) от рН среды
V
0 7 pH
Зависимость V (скорости) от рН среды
V
0 7 pH
Слайд 322. Зависимость скорости ферментативной реакции от температуры. График имеет колоколообразный характер. При
2. Зависимость скорости ферментативной реакции от температуры. График имеет колоколообразный характер. При

Слайд 33Однако при дальнейшем увеличении температуры происходит денатурация ферментов и резкое падение скорости
Однако при дальнейшем увеличении температуры происходит денатурация ферментов и резкое падение скорости

Слайд 34Зависимость V от t (температуры)
V
40○ t○
Зависимость V от t (температуры)
V
40○ t○
Слайд 35Не все ферменты имеют оптимум при 40 °С. Например каталаза имеет оптимум
Не все ферменты имеют оптимум при 40 °С. Например каталаза имеет оптимум

Слайд 36Зависимость V от количества фермента
V
0 кол-во Е
Зависимость V от количества фермента
V
0 кол-во Е
Слайд 374. Зависимость скорости ферментативной реакции от концентрации субстрата. Графическое изображение данной зависимости
4. Зависимость скорости ферментативной реакции от концентрации субстрата. Графическое изображение данной зависимости

Слайд 38Зависимость V ферментативной реакции от
концентрации S (субстрата)
V
Кm – константа Михаэлиса
Зависимость V ферментативной реакции от
концентрации S (субстрата)
V
Кm – константа Михаэлиса
Слайд 39Достижение максимальной скорости реакции зависит от сродства фермента к субстрату. Для того,
Достижение максимальной скорости реакции зависит от сродства фермента к субстрату. Для того,

Слайд 40Константа Михаэлиса – это такое количество субстрата, при котором достигается половина максимальной
Константа Михаэлиса – это такое количество субстрата, при котором достигается половина максимальной

Слайд 41Классификация ферментов
Оксидоредуктазы – обеспечивают перенос электронов и протонов. Участие в окислительно –
Классификация ферментов
Оксидоредуктазы – обеспечивают перенос электронов и протонов. Участие в окислительно –

Слайд 42ХАРАКТЕРИСТИКА КЛАССОВ ФЕРМЕНТОВ
I класс. Оксидоредуктазы катализируют окисление-дегидрирование в аэробных и анаэробных условиях.
Редуктазы
ХАРАКТЕРИСТИКА КЛАССОВ ФЕРМЕНТОВ
I класс. Оксидоредуктазы катализируют окисление-дегидрирование в аэробных и анаэробных условиях.
Редуктазы

Слайд 43Коферменты оксидоредуктаз (1)
Никотинамидные (НАД, НАДФ)
Флавиновые (ФМН, ФАД)
Металлопорфириновые (гемы a, b, c, d)
Хинонкоферменты
Коферменты оксидоредуктаз (1)
Никотинамидные (НАД, НАДФ)
Флавиновые (ФМН, ФАД)
Металлопорфириновые (гемы a, b, c, d)
Хинонкоферменты

Слайд 44II класс. Трансферазы – перенос атомных групп и молекулярных остатков. Подразделяется на
II класс. Трансферазы – перенос атомных групп и молекулярных остатков. Подразделяется на

Слайд 45Группы:
амино-, альдегидная,
сульфо–, кетонные остатки,
глюко–, фосфо–,
одноуглеродые фрагменты (CH3-, СН2-)
Пример: гексокиназа.
D – глю
Группы:
амино-, альдегидная,
сульфо–, кетонные остатки,
глюко–, фосфо–,
одноуглеродые фрагменты (CH3-, СН2-)
Пример: гексокиназа.
D – глю

Слайд 46Коферменты трансфераз (2)
Пиридоксиновые (ПАЛФ, ПАМФ)
Пантотеновые (КоА, дефосфо – КоА, 4 - фосфопантотенат)
Нуклеотидные
Коферменты трансфераз (2)
Пиридоксиновые (ПАЛФ, ПАМФ)
Пантотеновые (КоА, дефосфо – КоА, 4 - фосфопантотенат)
Нуклеотидные

Слайд 47III класс. Гидролазы – ферменты, которые осуществляют разрыв химической связи с присоединением
III класс. Гидролазы – ферменты, которые осуществляют разрыв химической связи с присоединением

Слайд 48IV класс. Лиазы – осуществляют негидролитические и неокислительные реакции распада и обратные
IV класс. Лиазы – осуществляют негидролитические и неокислительные реакции распада и обратные

Слайд 49Коферменты лиаз (4)
Пиридоксиновые (ПАЛФ)
Пантотеновые (КоА, дефосфо - КоА)
Тиаминовые (ТДФ)
Кобамидные (дезоксиаденозилкобаламин)
Коферменты лиаз (4)
Пиридоксиновые (ПАЛФ)
Пантотеновые (КоА, дефосфо - КоА)
Тиаминовые (ТДФ)
Кобамидные (дезоксиаденозилкобаламин)

Слайд 50V класс. Изомеразы – осуществляют внутримолекулярные превращения, изменения пространственного положения, перенос Н,
V класс. Изомеразы – осуществляют внутримолекулярные превращения, изменения пространственного положения, перенос Н,

Слайд 51Коферменты изомераз (5)
Пиридоксиновые (ПАЛФ)
Кобамидные (дезоксиаденозилкобаламин)
Фосфаты моносахаридов (глюкозо – 1, 6 – дифосфат,
Коферменты изомераз (5)
Пиридоксиновые (ПАЛФ)
Кобамидные (дезоксиаденозилкобаламин)
Фосфаты моносахаридов (глюкозо – 1, 6 – дифосфат,

Слайд 52VI класс. Лигазы – ферменты наращивания (синтетазы) осуществляют реакции синтеза с затратой
VI класс. Лигазы – ферменты наращивания (синтетазы) осуществляют реакции синтеза с затратой
Слайд 53Коферменты лигаз (6)
Нуклеотидные (УДФ – глюкоза, ЦДФ – холин и др.)
Биотиновые (карбоксибиотин)
Фолиевые
Коферменты лигаз (6)
Нуклеотидные (УДФ – глюкоза, ЦДФ – холин и др.)
Биотиновые (карбоксибиотин)
Фолиевые

Слайд 54НОМЕНКЛАТУРА ФЕРМЕНТОВ
Для ряда давно известных ферментов оставлены тривиальные названия: пепсин, трипсин, амилаза.
Кроме
НОМЕНКЛАТУРА ФЕРМЕНТОВ
Для ряда давно известных ферментов оставлены тривиальные названия: пепсин, трипсин, амилаза.
Кроме

Слайд 55Рабочее название:
Лактат + дегидрогенизация + аза
лактатдегидрогеназа
Систематическое название:
L – лактат: НАД+ - оксидоредуктаза
субстрат
Рабочее название:
Лактат + дегидрогенизация + аза
лактатдегидрогеназа
Систематическое название:
L – лактат: НАД+ - оксидоредуктаза
субстрат

 Волшебный витамин или химические элементы в живой клетке
Волшебный витамин или химические элементы в живой клетке Типчак. Распространение и экология
Типчак. Распространение и экология Вода-это жизнь
Вода-это жизнь Плауны. Хвощи. Папоротниковидные
Плауны. Хвощи. Папоротниковидные Мышцы головы и шеи
Мышцы головы и шеи Пищеварение у жвачных животных
Пищеварение у жвачных животных Проект. Микроскоп
Проект. Микроскоп Реакции клеточного метаболизма
Реакции клеточного метаболизма Клетки и состоящие из них ткани организма человека
Клетки и состоящие из них ткани организма человека Обмен веществ
Обмен веществ Викторина Отряды птиц
Викторина Отряды птиц Растения тоже воевали. 75-летию Победы посвящается
Растения тоже воевали. 75-летию Победы посвящается Нервная ткань
Нервная ткань Молекулярные узлы
Молекулярные узлы Ядро. Ядрышко. Кариотипирование
Ядро. Ядрышко. Кариотипирование Пророщення квасолі
Пророщення квасолі Растения степей
Растения степей Урожай на подоконнике
Урожай на подоконнике Викторина «МЛЕКОПИТАЮЩИЕ»
Викторина «МЛЕКОПИТАЮЩИЕ» Пути приспособления
Пути приспособления Скаты. Общие характеристики
Скаты. Общие характеристики Микропрепараты к 1 семестру
Микропрепараты к 1 семестру Міграції птахів, сезонні перельоти
Міграції птахів, сезонні перельоти Мышцы, типы мышц, их строение и значение. Работа мышц
Мышцы, типы мышц, их строение и значение. Работа мышц Презентация на тему Органы чувств
Презентация на тему Органы чувств  Лекция 7. Плазмодесмы. И вместе, и по отдельности…
Лекция 7. Плазмодесмы. И вместе, и по отдельности… Тип моллюски, или мягкотелые
Тип моллюски, или мягкотелые 1060021619
1060021619