- Физиология промежуточного мозга, коры больших полушарий

Содержание
- 2. Таламус. Функциональная характеристика и особенности ядерных групп.
- 4. Таламус - участвует в реализации сложных поведенческих реакций (инстинктов, влечений, эмоций), в регуляции и определении функционального
- 5. - происходит обработка практически всех сигналов, идущих в кору большого мозга от нижележащих отделов ЦНС. -
- 6. Функционально ядерные группы таламуса делятся на: специфические; ассоциативные ; неспецифические.
- 7. Функциональная классификация ядерных групп Специфические ядра (проекционные, релейные). Главные проекционные ядра, располагаются в боковой группе –медиальное
- 8. Особенности специфических ядер Имеют локальную проекцию в строго определенный участок коры (соматотопическая локализация); Моносинаптически связаны аксосоматическими
- 9. Особенности специфических ядер Основную массу клеток составляют «релейные» нейроны, которые имеют мало дендритов, небольшое тело, длинный
- 10. Функции специфических ядер Задние вентральные - проецируется соматосенсорная система (т.е. тактильная, болевая, температурная, проприоцептивная чувствит-ть). Латеральное
- 11. Ассоциативные ядра - медиодорсальные, латеральные, дорсальные и подушка таламуса. Особенности ассоциативных ядер Эфферентные волокна направляются к
- 12. Особенности ассоциативных ядер Основная афферентация поступает, главным образом, не из периферических отделов сенсорных систем, а от
- 13. Функции ассоциативных ядер Принимают участие в наиболее сложной обработке сенсорного сигнала, в оценке его биологической значимости
- 14. Неспецифические ядра таламуса - парацентральное ядро, вентральное переднее, ретикулярное ядро и др. Особенности неспецифических ядер -
- 15. - Афферентация поступает, в основном, из РФ мозга, гипоталамуса, лимбической системы, базальных ядер, специфических ядер таламуса;
- 16. Функции неспецифических ядер Оказывают облегчающее действие на корковые нейроны; Повышают возбудимость корковых нейронов; Улучшают способность корковых
- 17. Гипоталамус, структура, функции. Характеристика основных ядерных групп. Роль гипоталамуса в регуляции вегетативных функций организма и формировании
- 18. ГИПОТАЛАМУС
- 19. Структуры, расположенные ниже таламуса, объединяются под названием «гипоталамус». К нему относятся серый бугор, гипофиз, зрительный перекрест
- 20. Имеет мощные афферентные связи с РФ, лимбической системой, базальными ганглиями, таламусом, корой большого мозга. Получает информацию
- 21. Гипоталамус - высший подкорковый центр интеграции вегетативных, эмоциональных, моторных компонентов сложных реакций адаптивного поведения организма и
- 22. Особенности нейронных групп гипоталамуса Обладают высокой чувствительностью (химической и физико-химической) к параметрам крови и их изменениям;
- 23. Трофотропная система Передние отделы гипоталамуса (преоптическая зона) – парасимпатические эффекты Вызывает анаболические процессы Обеспечивает нутритивные функции
- 24. Эрготропная система Задние отделы (мамиллярные тела) – симпатические эффекты Способствуют приспособлению к меняющимся условиям внешней среды
- 25. Также в гипоталамусе располагаются мотивационные центры: терморегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения
- 27. Лимбическая система мозга. Структуры, входящие в ее состав, их функции (Дж. Пейпец). Интегративная роль лимбичекой системы
- 28. Лимбическая система - Корковые области (гиппокамп, парагиппокампова извилина, поясная извилина и филогенетически старые структуры обонятельного мозга
- 31. Функции лимбической системы Регуляция висцеральных функций (висцеральный мозг) Формирование эмоций Участвует в процессах памяти и обучения
- 32. Для ЛС характерно наличие как простых двухсторонних связей и так сложных путей, образующих большое количество замкнутых
- 33. - круг Пейпеса (гиппокамп—сосцевидные тела— передние ядра таламуса–- кора поясной извилины —парагиппокампова извилина — гиппокамп) –
- 36. Функциональная характеристика базальных ганглиев. Их структурно-функциональная характеристика и роль в формировании мышечного тонуса и двигательных программ.
- 37. БАЗАЛЬНЫЕ ГАНГЛИИ Являются важным подкорковым связующим звеном между ассоциативными и двигательными областями коры головного мозга. Их
- 40. Базальные ганглии – хвостатые ядра, скорлупа (неостриатум), бледный шар (палеостриатум) и ограда Хвостатые ядра Играют роль
- 41. У человека поражение хвостатых ядер приводит к мнестическим расстройствам, изменению условных рефлексов (нарушение выработки, исчезновение), расстройствам
- 42. Бледный шар Полное двухстороннее разрушение бледного шара приведет к глубоким изменениям ВНД, исчезновению ранее выработанных условных
- 43. Функции базальных ядер Участвуют в поддержании мышечного тонуса, позы и движений. Формируют программы инициации движения и
- 44. Функции базальных ядер Помогают планировать и осуществлять сложные двигательные программы, регулируя относительную интенсивность и направление отдельных
- 48. Т.о. базальные ядра участвуют в реализации моторных программ, эмоций, мотиваций, высших психических функций.
- 49. Роль различных отделов коры в регуляции физиологических функций. Афферентные, эфферентные и ассоциативные области. Топическая локализация функций
- 50. Кора имеет ряд морфофункциональных особенностей: - многослойность расположения нейронов; - модульный принцип организации; - цитоархитектоническое распределение
- 51. - экранность (экранный принцип организации коры, который заключается в том, что рецептор проецирует свой сигнал не
- 53. Колонковая организация коры Наряду с горизонтальной организацией по слоям в неокортексе имеется четкая вертикальная организация в
- 54. Свойства колонки корковых нейронов Все нейроны колонки реагируют на одну и ту же модальность однотипных сенсорных
- 55. . Модульный принцип структурно-функциональной организации коры головного мозга В одном нейронном модуле осуществляется локальная переработка информации
- 56. К. Бродман (1910) составил цитоархитектоническую карту коры, в которой выделено 11 областей, включающих 52 поля. В
- 58. Корковые моторные области Первичная двигательная моторная кора –прецентральная извилина (поле 4 по Бродману) Прилегающая к ней
- 59. Моторные двигательные области М1 и М2 (поле 4 по Бродману) Имеет двухсторонние внутрикорковые связи со всеми
- 61. Сенсорные (проекционные, афферентные) зоны) 1,2,3 поля – постцентральная извилина –соматосенсорная чувствительность 17,18,19 поля – затылочная область
- 62. Сенсорные (проекционные, афферентные) зоны) Функции - осуществляют высший уровень сенсорного анализа; получают афферентацию от специфических релейных
- 63. Зрительная сенсорная система. Первая зрительная область (17 поле): при локальном разрушении этого поля выпадают соответствующие участки
- 64. Слуховая сенсорная система Первичная слуховая зона (41, 42 поля): заканчиваются аксоны релейных нейронов МКТ таламуса, несущие
- 65. Соматосенсорные проекционные области - анализирует сигналы кожной, мышечной и висцеральной чувствительности. Первая (S1) соматосенсорная зона -
- 68. Обонятельная сенсорная система Проецируется в области переднего конца гиппокампальной извилины (поле 34), гетеротипическая. Раздражение приводит к
- 69. Ассоциативные зоны Основные: теменная (поля 5, 7, 39, 40) и лобная (8, 9, 10, 11, 12)
- 70. Основные ассоциативные области Теменная (поля 5, 7, 39, 40) - обеспечивает воссоздание целостностных образов предметов и
- 71. Основные ассоциативные области Лобная (поля 8, 9, 10, 11, 12) - Участвует в реализации наиболее сложных
- 73. Электроэнцефалография- ЭЭГ заключается в регистрации ритмических изменений потенциалов определенных областей коры большого мозга между двумя активными
- 74. Ритмы ЭЭГ отражают возбудительные и тормозные постсинаптические процессы тел и дендритов нейронов различных генераторных систем (корковых,
- 77. Пластичность нервной системы - способность к адекватным перестройкам функциональной организации мозга в ответ на значимые изменения
- 78. Уровни пластичности Макроуровень Связан с изменением сетевой структуры мозга, обеспечивающей сообщение между полушариями и между различными
- 79. Причины, вызывающие перестройку функциональной организации мозга Повреждение самого мозга (инсульт моторной коры, в результате которого мышцы
- 80. Факторы, определяющие восстановительные способности мозга Возраст пациента Длительность воздействия повреждающего объекта. Локализация повреждения мозга. Обширность поражения.
- 81. Кортико - висцеральные взаимоотношения (К.М. Быков) Естественное функциональное взаимодействие между корой больших полушарий головного мозга и
- 83. Скачать презентацию

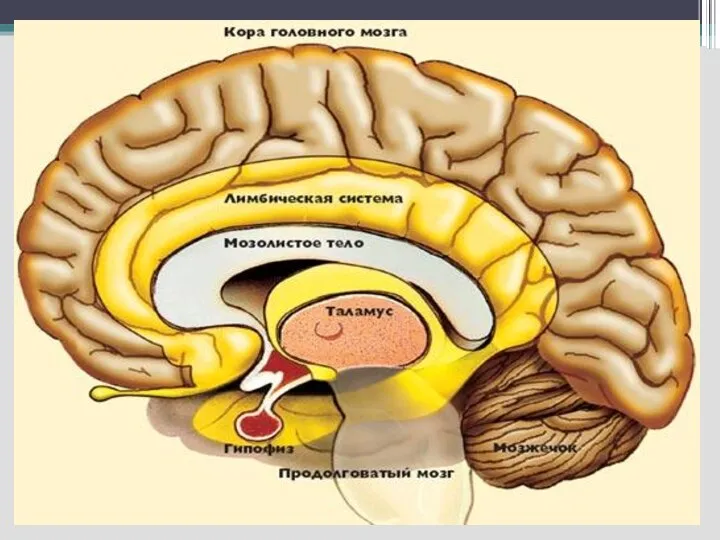
Слайд 4 Таламус
- участвует в реализации сложных поведенческих реакций (инстинктов, влечений, эмоций),
Таламус
- участвует в реализации сложных поведенческих реакций (инстинктов, влечений, эмоций),

Слайд 5 - происходит обработка практически всех сигналов, идущих в кору большого мозга
- происходит обработка практически всех сигналов, идущих в кору большого мозга

Слайд 6 Функционально ядерные группы таламуса делятся на:
специфические;
ассоциативные ;
неспецифические.
Функционально ядерные группы таламуса делятся на:
специфические;
ассоциативные ;
неспецифические.

Слайд 7Функциональная классификация ядерных групп
Специфические ядра (проекционные, релейные). Главные проекционные ядра, располагаются в
Функциональная классификация ядерных групп
Специфические ядра (проекционные, релейные). Главные проекционные ядра, располагаются в

Слайд 8Особенности специфических ядер
Имеют локальную проекцию в строго определенный участок коры (соматотопическая локализация);
Моносинаптически
Особенности специфических ядер
Имеют локальную проекцию в строго определенный участок коры (соматотопическая локализация);
Моносинаптически

Слайд 9Особенности специфических ядер
Основную массу клеток составляют «релейные» нейроны, которые имеют мало дендритов,
Особенности специфических ядер
Основную массу клеток составляют «релейные» нейроны, которые имеют мало дендритов,

Слайд 10Функции специфических ядер
Задние вентральные - проецируется соматосенсорная система (т.е. тактильная, болевая, температурная,
Функции специфических ядер
Задние вентральные - проецируется соматосенсорная система (т.е. тактильная, болевая, температурная,

Слайд 11 Ассоциативные ядра - медиодорсальные, латеральные, дорсальные и подушка таламуса.
Особенности ассоциативных
Ассоциативные ядра - медиодорсальные, латеральные, дорсальные и подушка таламуса.
Особенности ассоциативных

Слайд 12Особенности ассоциативных ядер
Основная афферентация поступает, главным образом, не из периферических отделов сенсорных
Особенности ассоциативных ядер
Основная афферентация поступает, главным образом, не из периферических отделов сенсорных

Слайд 13Функции ассоциативных ядер
Принимают участие в наиболее сложной обработке сенсорного сигнала, в оценке
Функции ассоциативных ядер
Принимают участие в наиболее сложной обработке сенсорного сигнала, в оценке

Слайд 14 Неспецифические ядра таламуса - парацентральное ядро, вентральное переднее, ретикулярное ядро и
Неспецифические ядра таламуса - парацентральное ядро, вентральное переднее, ретикулярное ядро и

Слайд 15- Афферентация поступает, в основном, из РФ мозга, гипоталамуса, лимбической системы, базальных
- Афферентация поступает, в основном, из РФ мозга, гипоталамуса, лимбической системы, базальных

Слайд 16Функции неспецифических ядер
Оказывают облегчающее действие на корковые нейроны;
Повышают возбудимость корковых нейронов;
Улучшают способность
Функции неспецифических ядер
Оказывают облегчающее действие на корковые нейроны;
Повышают возбудимость корковых нейронов;
Улучшают способность

Слайд 17Гипоталамус, структура, функции. Характеристика основных ядерных групп. Роль гипоталамуса в регуляции вегетативных
Гипоталамус, структура, функции. Характеристика основных ядерных групп. Роль гипоталамуса в регуляции вегетативных

Слайд 18ГИПОТАЛАМУС
ГИПОТАЛАМУС
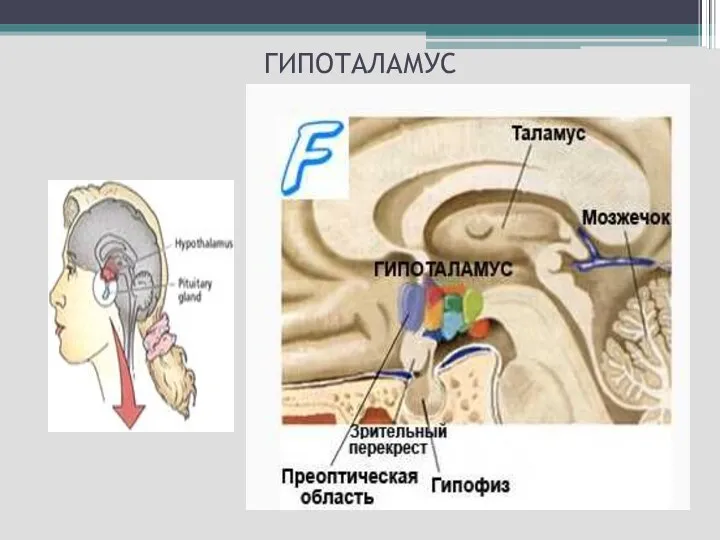
Слайд 19 Структуры, расположенные ниже таламуса, объединяются под названием «гипоталамус». К нему относятся
Структуры, расположенные ниже таламуса, объединяются под названием «гипоталамус». К нему относятся

Слайд 20Имеет мощные афферентные связи с РФ, лимбической системой, базальными ганглиями, таламусом, корой
Имеет мощные афферентные связи с РФ, лимбической системой, базальными ганглиями, таламусом, корой

Слайд 21 Гипоталамус - высший подкорковый центр интеграции вегетативных, эмоциональных, моторных компонентов сложных
Гипоталамус - высший подкорковый центр интеграции вегетативных, эмоциональных, моторных компонентов сложных

Слайд 22Особенности нейронных групп гипоталамуса
Обладают высокой чувствительностью (химической и физико-химической) к параметрам крови
Особенности нейронных групп гипоталамуса
Обладают высокой чувствительностью (химической и физико-химической) к параметрам крови

Слайд 23Трофотропная система
Передние отделы гипоталамуса (преоптическая зона) – парасимпатические эффекты
Вызывает анаболические процессы
Обеспечивает нутритивные
Трофотропная система
Передние отделы гипоталамуса (преоптическая зона) – парасимпатические эффекты
Вызывает анаболические процессы
Обеспечивает нутритивные

Слайд 24Эрготропная система
Задние отделы (мамиллярные тела) – симпатические эффекты
Способствуют приспособлению к меняющимся условиям
Эрготропная система
Задние отделы (мамиллярные тела) – симпатические эффекты
Способствуют приспособлению к меняющимся условиям

Слайд 25 Также в гипоталамусе располагаются мотивационные центры: терморегуляции, голода и насыщения, жажды
Также в гипоталамусе располагаются мотивационные центры: терморегуляции, голода и насыщения, жажды

Слайд 27Лимбическая система мозга. Структуры, входящие в ее состав, их функции (Дж. Пейпец).
Лимбическая система мозга. Структуры, входящие в ее состав, их функции (Дж. Пейпец).

Слайд 28Лимбическая система
- Корковые области (гиппокамп, парагиппокампова извилина, поясная извилина и филогенетически старые
Лимбическая система
- Корковые области (гиппокамп, парагиппокампова извилина, поясная извилина и филогенетически старые


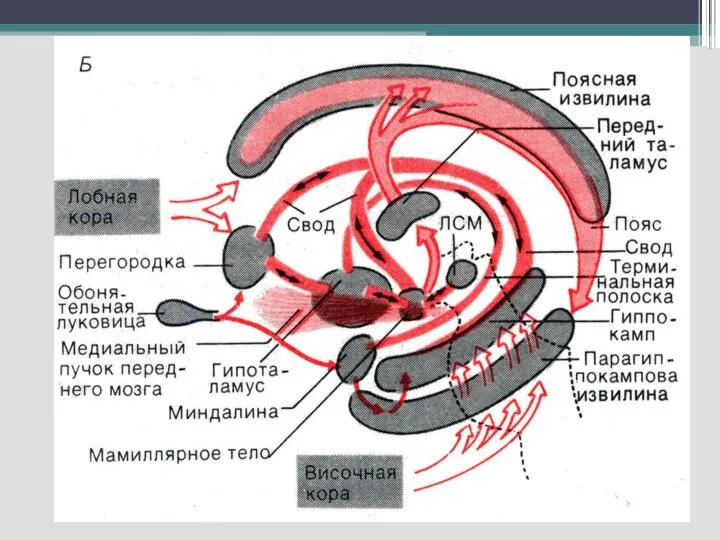
Слайд 31Функции лимбической системы
Регуляция висцеральных функций (висцеральный мозг)
Формирование эмоций
Участвует в процессах памяти
Функции лимбической системы
Регуляция висцеральных функций (висцеральный мозг)
Формирование эмоций
Участвует в процессах памяти

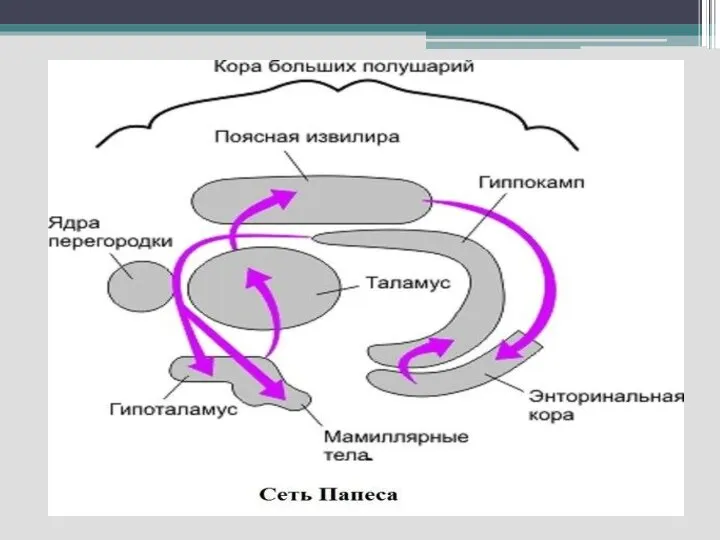
Слайд 32Для ЛС характерно наличие как простых двухсторонних связей и так сложных путей,
Для ЛС характерно наличие как простых двухсторонних связей и так сложных путей,

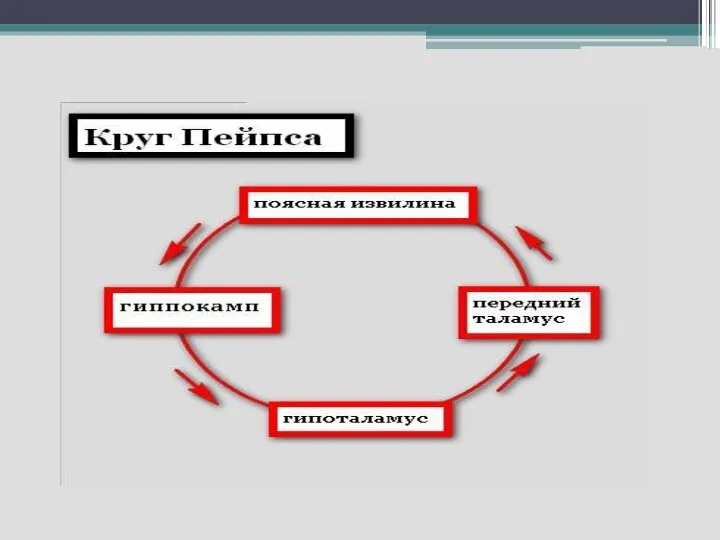
Слайд 33- круг Пейпеса (гиппокамп—сосцевидные тела— передние ядра таламуса–- кора поясной извилины —парагиппокампова
- круг Пейпеса (гиппокамп—сосцевидные тела— передние ядра таламуса–- кора поясной извилины —парагиппокампова

Слайд 36Функциональная характеристика базальных ганглиев. Их структурно-функциональная характеристика и роль в формировании мышечного
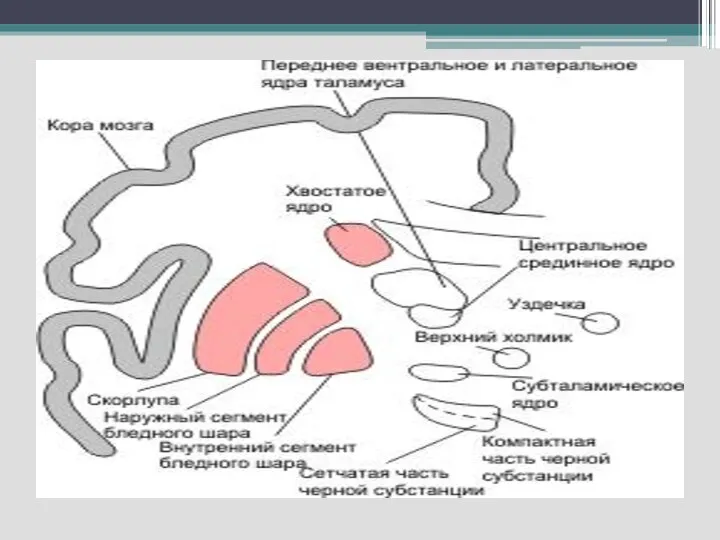
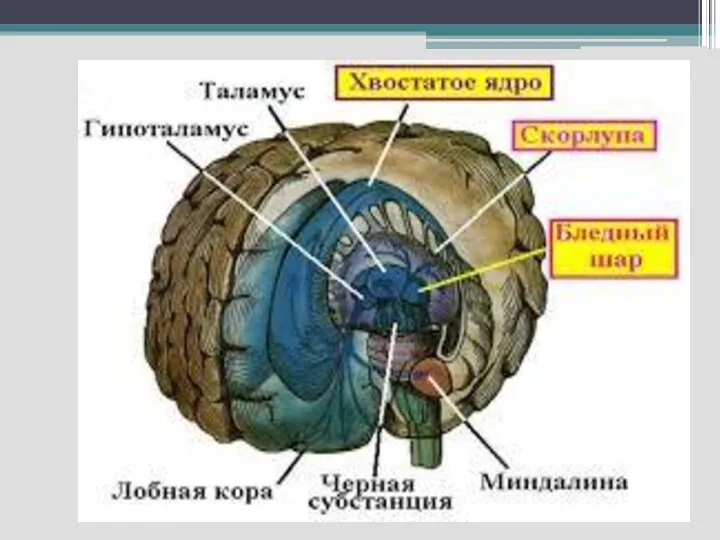
Функциональная характеристика базальных ганглиев. Их структурно-функциональная характеристика и роль в формировании мышечного

Слайд 37БАЗАЛЬНЫЕ ГАНГЛИИ
Являются важным подкорковым связующим звеном между ассоциативными и двигательными областями коры
БАЗАЛЬНЫЕ ГАНГЛИИ
Являются важным подкорковым связующим звеном между ассоциативными и двигательными областями коры

Слайд 40Базальные ганглии – хвостатые ядра, скорлупа (неостриатум), бледный шар (палеостриатум) и ограда
Хвостатые
Базальные ганглии – хвостатые ядра, скорлупа (неостриатум), бледный шар (палеостриатум) и ограда
Хвостатые

Слайд 41У человека поражение хвостатых ядер приводит к мнестическим расстройствам, изменению условных рефлексов
У человека поражение хвостатых ядер приводит к мнестическим расстройствам, изменению условных рефлексов

Слайд 42Бледный шар
Полное двухстороннее разрушение бледного шара приведет к глубоким изменениям ВНД, исчезновению
Бледный шар
Полное двухстороннее разрушение бледного шара приведет к глубоким изменениям ВНД, исчезновению

Слайд 43Функции базальных ядер
Участвуют в поддержании мышечного тонуса, позы и движений.
Формируют программы инициации
Функции базальных ядер
Участвуют в поддержании мышечного тонуса, позы и движений.
Формируют программы инициации

Слайд 44Функции базальных ядер
Помогают планировать и осуществлять сложные двигательные программы, регулируя относительную интенсивность
Функции базальных ядер
Помогают планировать и осуществлять сложные двигательные программы, регулируя относительную интенсивность

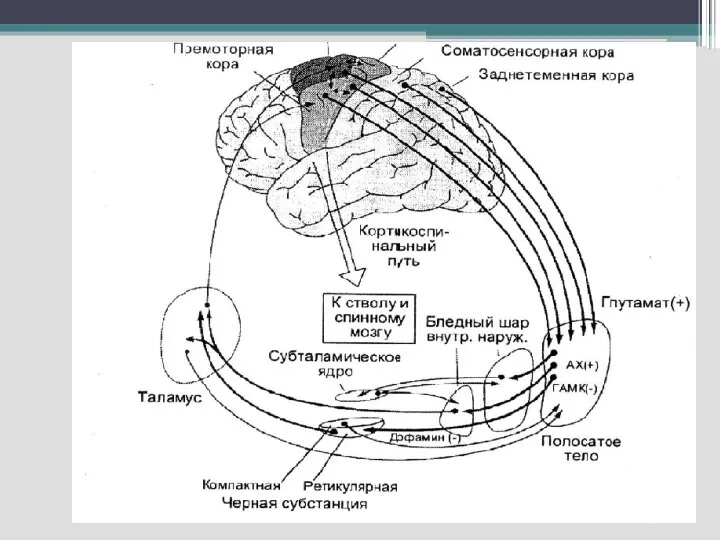

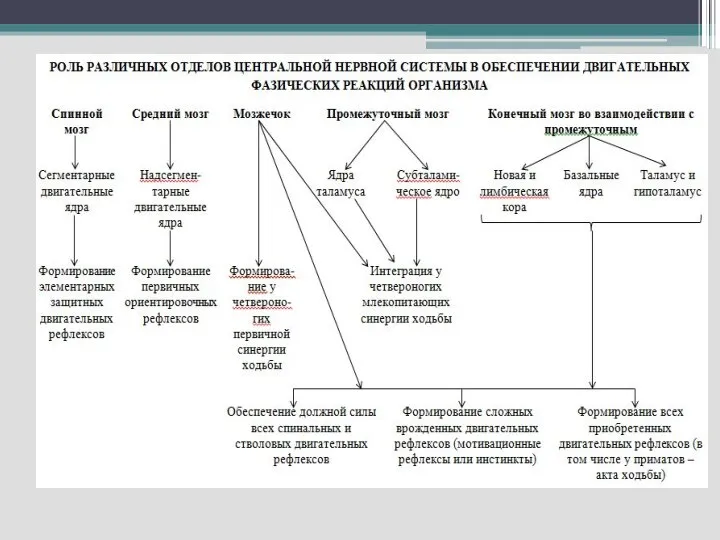
Слайд 48Т.о. базальные ядра участвуют в реализации моторных программ, эмоций, мотиваций, высших психических
Т.о. базальные ядра участвуют в реализации моторных программ, эмоций, мотиваций, высших психических

Слайд 49 Роль различных отделов коры в регуляции физиологических функций. Афферентные, эфферентные и
Роль различных отделов коры в регуляции физиологических функций. Афферентные, эфферентные и

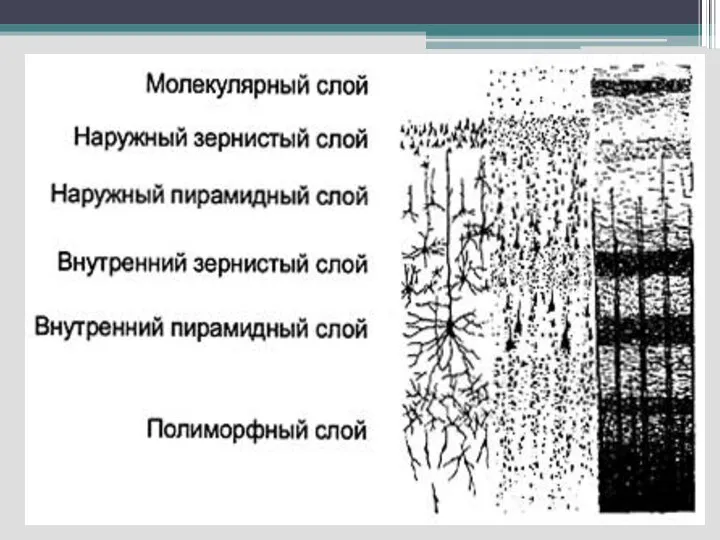
Слайд 50 Кора имеет ряд морфофункциональных особенностей:
- многослойность расположения нейронов;
-
Кора имеет ряд морфофункциональных особенностей:
- многослойность расположения нейронов;
-

Слайд 51 - экранность (экранный принцип организации коры, который заключается в том, что
- экранность (экранный принцип организации коры, который заключается в том, что

Слайд 53Колонковая организация коры
Наряду с горизонтальной организацией по слоям в неокортексе имеется
Колонковая организация коры
Наряду с горизонтальной организацией по слоям в неокортексе имеется

Слайд 54Свойства колонки корковых нейронов
Все нейроны колонки реагируют на одну и ту же
Свойства колонки корковых нейронов
Все нейроны колонки реагируют на одну и ту же

Слайд 55. Модульный принцип структурно-функциональной организации коры головного мозга
В одном нейронном модуле осуществляется
. Модульный принцип структурно-функциональной организации коры головного мозга
В одном нейронном модуле осуществляется

Слайд 56К. Бродман (1910) составил цитоархитектоническую карту коры, в которой выделено 11 областей,
К. Бродман (1910) составил цитоархитектоническую карту коры, в которой выделено 11 областей,

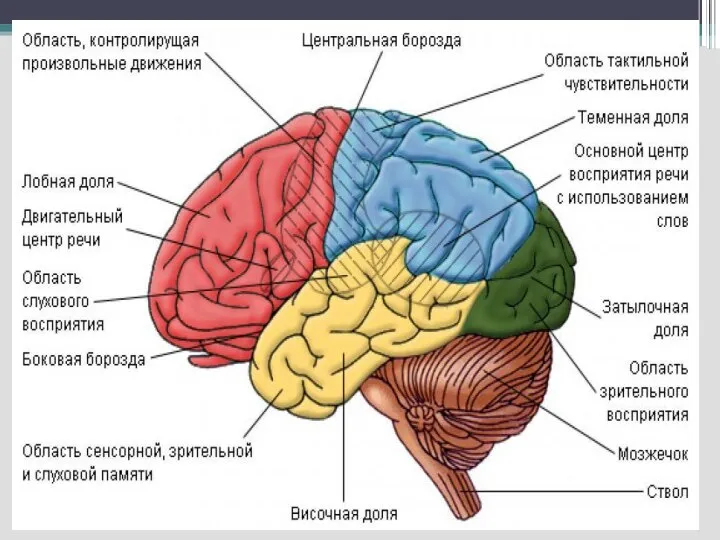
Слайд 58Корковые моторные области
Первичная двигательная моторная кора –прецентральная извилина (поле 4 по Бродману)
Прилегающая
Корковые моторные области
Первичная двигательная моторная кора –прецентральная извилина (поле 4 по Бродману)
Прилегающая

Слайд 59Моторные двигательные области М1 и М2 (поле 4 по Бродману)
Имеет двухсторонние внутрикорковые
Моторные двигательные области М1 и М2 (поле 4 по Бродману)
Имеет двухсторонние внутрикорковые

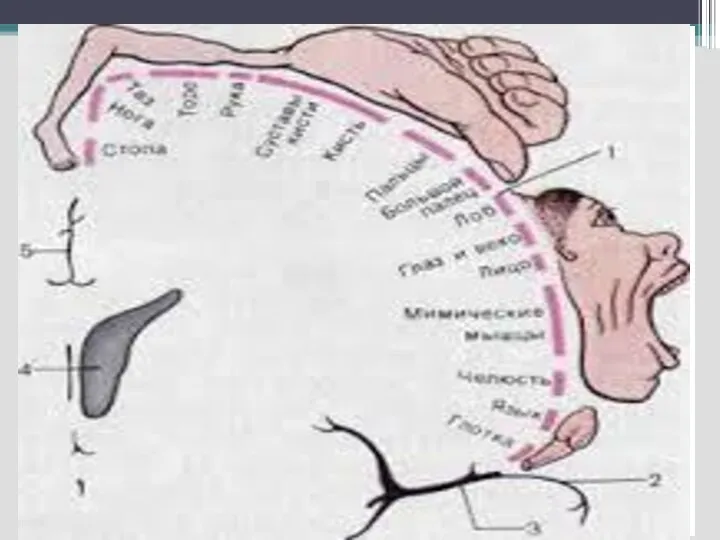
Слайд 61 Сенсорные (проекционные, афферентные) зоны)
1,2,3 поля – постцентральная извилина –соматосенсорная чувствительность
17,18,19 поля
Сенсорные (проекционные, афферентные) зоны)
1,2,3 поля – постцентральная извилина –соматосенсорная чувствительность
17,18,19 поля

Слайд 62 Сенсорные (проекционные, афферентные) зоны)
Функции
- осуществляют высший уровень сенсорного анализа;
получают афферентацию
Сенсорные (проекционные, афферентные) зоны)
Функции
- осуществляют высший уровень сенсорного анализа;
получают афферентацию

Слайд 63 Зрительная сенсорная система.
Первая зрительная область (17 поле): при локальном разрушении
Зрительная сенсорная система.
Первая зрительная область (17 поле): при локальном разрушении

Слайд 64 Слуховая сенсорная система
Первичная слуховая зона (41, 42 поля): заканчиваются аксоны
Слуховая сенсорная система
Первичная слуховая зона (41, 42 поля): заканчиваются аксоны

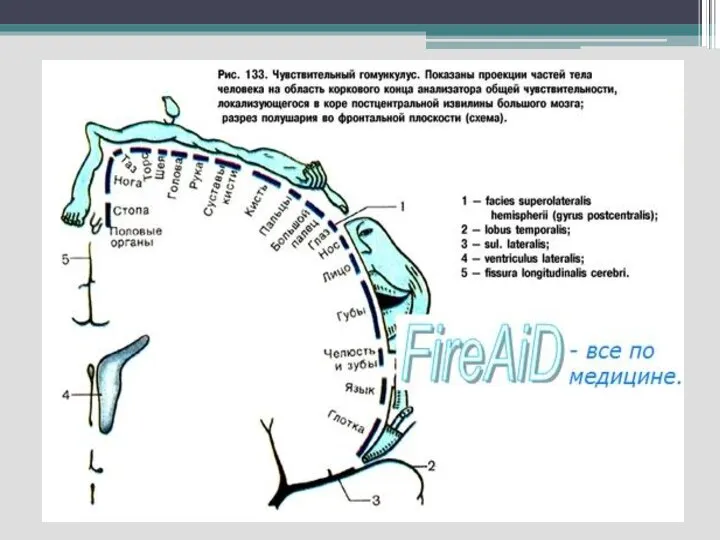
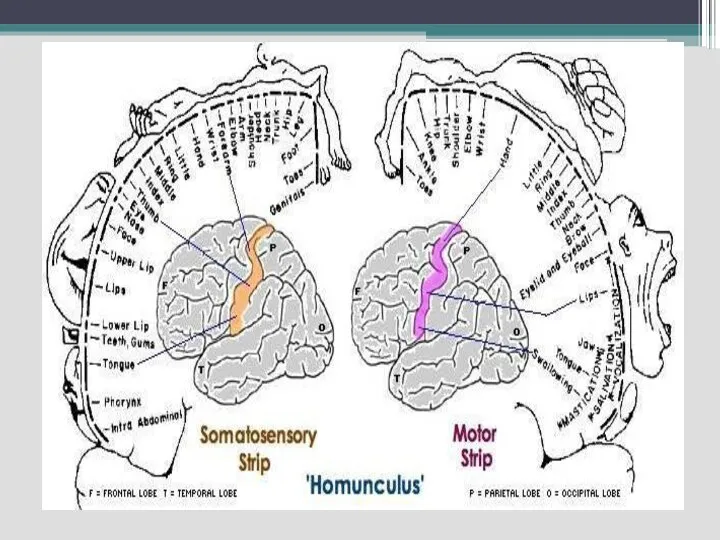
Слайд 65
Соматосенсорные проекционные области
- анализирует сигналы кожной, мышечной и висцеральной
Соматосенсорные проекционные области
- анализирует сигналы кожной, мышечной и висцеральной

Слайд 68
Обонятельная сенсорная система
Проецируется в области переднего конца гиппокампальной извилины (поле
Обонятельная сенсорная система
Проецируется в области переднего конца гиппокампальной извилины (поле

Слайд 69 Ассоциативные зоны
Основные: теменная (поля 5, 7, 39, 40) и лобная
Ассоциативные зоны
Основные: теменная (поля 5, 7, 39, 40) и лобная

Слайд 70Основные ассоциативные области
Теменная (поля 5, 7, 39, 40)
- обеспечивает воссоздание целостностных образов
Основные ассоциативные области
Теменная (поля 5, 7, 39, 40)
- обеспечивает воссоздание целостностных образов

Слайд 71Основные ассоциативные области
Лобная (поля 8, 9, 10, 11, 12)
- Участвует в реализации
Основные ассоциативные области
Лобная (поля 8, 9, 10, 11, 12)
- Участвует в реализации


Слайд 73 Электроэнцефалография-
ЭЭГ заключается в регистрации ритмических изменений потенциалов определенных областей коры
Электроэнцефалография-
ЭЭГ заключается в регистрации ритмических изменений потенциалов определенных областей коры

Слайд 74 Ритмы ЭЭГ отражают возбудительные и тормозные постсинаптические процессы тел и дендритов
Ритмы ЭЭГ отражают возбудительные и тормозные постсинаптические процессы тел и дендритов



Слайд 77Пластичность нервной системы
- способность к адекватным перестройкам функциональной организации мозга в
Пластичность нервной системы
- способность к адекватным перестройкам функциональной организации мозга в

Слайд 78Уровни пластичности
Макроуровень
Связан с изменением сетевой структуры мозга, обеспечивающей сообщение между полушариями и
Уровни пластичности
Макроуровень
Связан с изменением сетевой структуры мозга, обеспечивающей сообщение между полушариями и

Слайд 79Причины, вызывающие перестройку функциональной организации мозга
Повреждение самого мозга (инсульт моторной коры, в
Причины, вызывающие перестройку функциональной организации мозга
Повреждение самого мозга (инсульт моторной коры, в

Слайд 80Факторы, определяющие восстановительные способности мозга
Возраст пациента
Длительность воздействия повреждающего объекта.
Локализация повреждения
Факторы, определяющие восстановительные способности мозга
Возраст пациента
Длительность воздействия повреждающего объекта.
Локализация повреждения

Слайд 81Кортико - висцеральные взаимоотношения (К.М. Быков)
Естественное функциональное взаимодействие между корой больших полушарий
Кортико - висцеральные взаимоотношения (К.М. Быков)
Естественное функциональное взаимодействие между корой больших полушарий

 Устьица и чечевички. Замыкающие клетки. Устьичная щель
Устьица и чечевички. Замыкающие клетки. Устьичная щель Феномен человека в естественнонаучной картине мира
Феномен человека в естественнонаучной картине мира Строение дыхательной системы
Строение дыхательной системы Вегетативная нервная система
Вегетативная нервная система Вирусы – внутриклеточные паразиты
Вирусы – внутриклеточные паразиты Презентация на тему "Жизнь земноводных весной" - презентации по Биологии
Презентация на тему "Жизнь земноводных весной" - презентации по Биологии Лимфатическая система
Лимфатическая система Органоиды клетки
Органоиды клетки Опровержение теории Ч. Дарвина
Опровержение теории Ч. Дарвина prezentatsia_ChMN_1-12p
prezentatsia_ChMN_1-12p Периоды эмбриогенеза
Периоды эмбриогенеза Остеология. Осевой скелет
Остеология. Осевой скелет Аномальный рост
Аномальный рост Внутреннее строение рыбы
Внутреннее строение рыбы Презентация на тему Особенности высшей нервной деятельности человека
Презентация на тему Особенности высшей нервной деятельности человека  Особенности организации плоских червей. Важнейшие ароморфозы
Особенности организации плоских червей. Важнейшие ароморфозы Домашние животные. Формирование, расширение и обобщение знаний детей о домашних животных
Домашние животные. Формирование, расширение и обобщение знаний детей о домашних животных Сарычев Н. Г. Животноводство с основами общей зоогигиены
Сарычев Н. Г. Животноводство с основами общей зоогигиены Зеленая аптека Донбасса. Угадай растение. Викторина
Зеленая аптека Донбасса. Угадай растение. Викторина 1. Введение в медицинскую генетику. Цитологические основы наследственности
1. Введение в медицинскую генетику. Цитологические основы наследственности Обмен серосодержащих аминокислот
Обмен серосодержащих аминокислот Лишайники. Биологический диктант
Лишайники. Биологический диктант Строение тела человека. 2 класс
Строение тела человека. 2 класс Царство животные
Царство животные Антропологическая альтернатива и её принципы: многообразие, целостности, субстанциональности, свободы
Антропологическая альтернатива и её принципы: многообразие, целостности, субстанциональности, свободы Презентация на тему ТИП КОЛЬЧАТЫЕ ЧЕРВИ
Презентация на тему ТИП КОЛЬЧАТЫЕ ЧЕРВИ  Скелет человека
Скелет человека История возникновения ВИЧ. Биологические аспекты ВИЧ. Развитие ВИЧ-инфекции
История возникновения ВИЧ. Биологические аспекты ВИЧ. Развитие ВИЧ-инфекции