- Генетика різноманітності антиген-специфічних рецепторів

Содержание
- 2. Теорії утворення різноманітності антитіл. Селекційно-клональна теорія Бернета. Теорія “Зародкового різноманіття”. Теорія “Соматичного різноманіття”. Теорія В. Дрейєра
- 3. В наш час вважають, що синтез імуноґлобулінів кодують певні генетичні сеґменти, які об’єднуються разом ще в
- 4. Методи дослідження генів імуноглобулінів і антиген-специфічних рецепторів. Отримання мРНК важких і легких ланцюгів імуноґлобулінів. Отримання кДНК
- 5. Дослідження С. Тонегави. Досліджували та отримали мРНК важких і легких ланцюгів імуноґлобулінів та встановили приблизну кількість
- 6. ОРГАНІЗАЦІЯ ГЕНІВ ІМУНОҐЛОБУЛІНІВ Важкі й легкі ланцюги імуноглобулінів кодуються трьома локусами генів: Двома локусами - κ
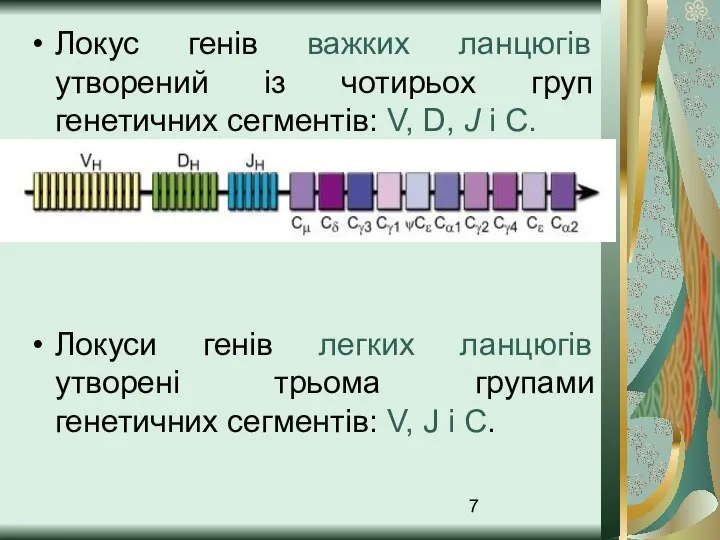
- 7. Локус генів важких ланцюгів утворений із чотирьох груп генетичних сегментів: V, D, J і C. Локуси
- 8. Організація генів імуноглобулінів людини.
- 9. Схема утворення легких ланцюгів імуноглобулінів із зародкової ДНК реаранжуванням генів.
- 10. Схема утворення легких (а) і важких (б) ланцюгів імуноглобулінів із зародкової ДНК реаранжуванням генів.
- 11. Переключення генів важких ланцюгів. В ембріональних клітинах J-сегменти містяться лише перед Сμ-генним сегментом. Тому внаслідок рекомбінації
- 12. Переключення синтезу ІgМ та ІgD. Комплекс, що складається з об'єднаних генних сегментів VDJ, може експресуватися з
- 13. Під час транскрипції утворюється первинна РНК, яка відповідає VDJСμСδ-гену, що включає екзони, здатні кодувати константну частину
- 14. Після експресії ІgМ та ІgD В-клітина може переключитися на синтез інших класів імуноглобулінів залежно від типу
- 16. Скачать презентацию
Слайд 2Теорії утворення різноманітності антитіл.
Селекційно-клональна теорія Бернета.
Теорія “Зародкового різноманіття”.
Теорія “Соматичного різноманіття”.
Теорія В. Дрейєра
Теорії утворення різноманітності антитіл.
Селекційно-клональна теорія Бернета.
Теорія “Зародкового різноманіття”.
Теорія “Соматичного різноманіття”.
Теорія В. Дрейєра

Слайд 3В наш час вважають, що синтез імуноґлобулінів кодують певні генетичні сеґменти, які
В наш час вважають, що синтез імуноґлобулінів кодують певні генетичні сеґменти, які

Слайд 4Методи дослідження генів імуноглобулінів і антиген-специфічних рецепторів.
Отримання мРНК важких і легких ланцюгів
Методи дослідження генів імуноглобулінів і антиген-специфічних рецепторів.
Отримання мРНК важких і легких ланцюгів

Слайд 5Дослідження С. Тонегави.
Досліджували та отримали мРНК важких і легких ланцюгів імуноґлобулінів та
Дослідження С. Тонегави.
Досліджували та отримали мРНК важких і легких ланцюгів імуноґлобулінів та

Слайд 6ОРГАНІЗАЦІЯ ГЕНІВ ІМУНОҐЛОБУЛІНІВ
Важкі й легкі ланцюги імуноглобулінів кодуються трьома локусами генів:
ОРГАНІЗАЦІЯ ГЕНІВ ІМУНОҐЛОБУЛІНІВ
Важкі й легкі ланцюги імуноглобулінів кодуються трьома локусами генів:

Слайд 7Локус генів важких ланцюгів утворений із чотирьох груп генетичних сегментів: V, D,
Локус генів важких ланцюгів утворений із чотирьох груп генетичних сегментів: V, D,
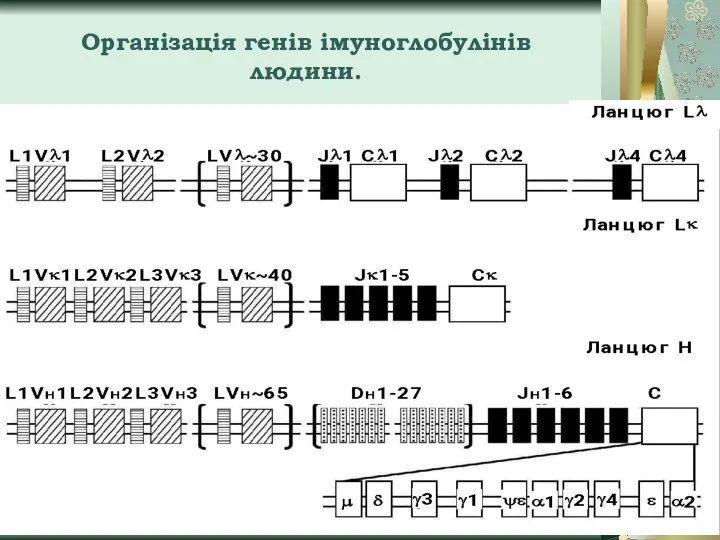
Слайд 8Організація генів імуноглобулінів людини.
Організація генів імуноглобулінів людини.
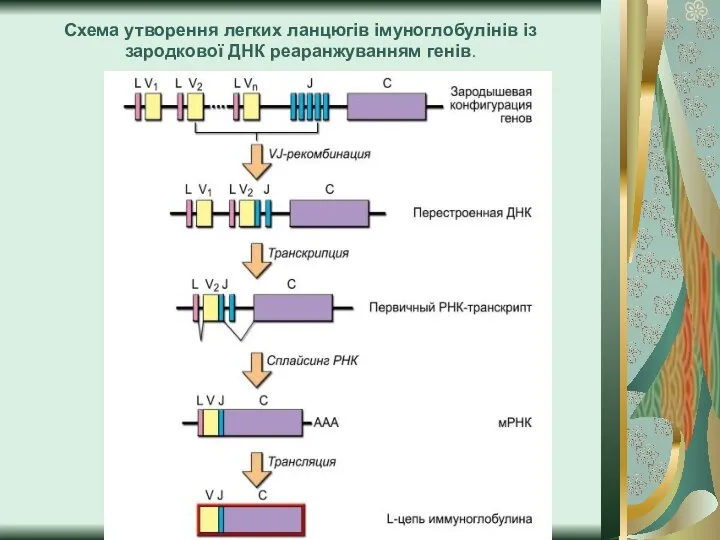
Слайд 9Схема утворення легких ланцюгів імуноглобулінів із зародкової ДНК реаранжуванням генів.
Схема утворення легких ланцюгів імуноглобулінів із зародкової ДНК реаранжуванням генів.
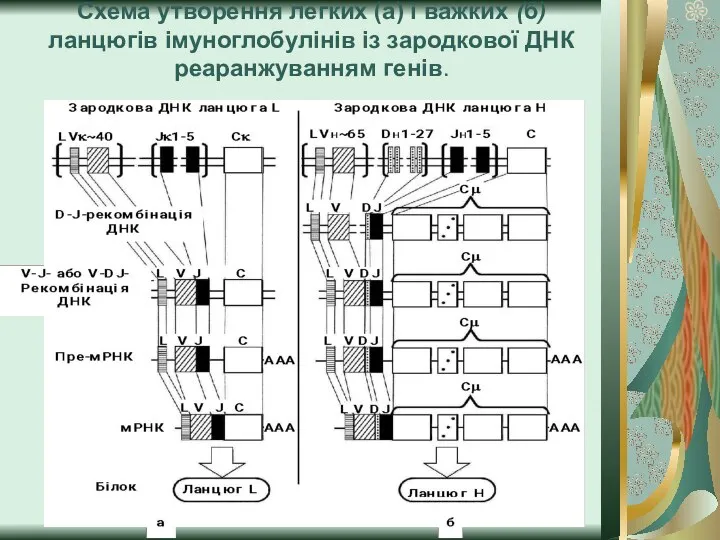
Слайд 10Схема утворення легких (а) і важких (б) ланцюгів імуноглобулінів із зародкової ДНК
Схема утворення легких (а) і важких (б) ланцюгів імуноглобулінів із зародкової ДНК
Слайд 11Переключення генів важких ланцюгів.
В ембріональних клітинах J-сегменти містяться лише перед Сμ-генним
Переключення генів важких ланцюгів.
В ембріональних клітинах J-сегменти містяться лише перед Сμ-генним

Слайд 12Переключення синтезу ІgМ та ІgD.
Комплекс, що складається з об'єднаних генних сегментів
Переключення синтезу ІgМ та ІgD.
Комплекс, що складається з об'єднаних генних сегментів

Слайд 13Під час транскрипції утворюється первинна РНК, яка відповідає VDJСμСδ-гену, що включає екзони,
Під час транскрипції утворюється первинна РНК, яка відповідає VDJСμСδ-гену, що включає екзони,

Слайд 14Після експресії ІgМ та ІgD В-клітина може переключитися на синтез інших класів

 Росянка. Очаровательный убийца
Росянка. Очаровательный убийца Биологические парки Татарстана
Биологические парки Татарстана Әр түрлі жастағы егеуқұйрықтардың тамақтану рационына асқабақ пен топинамбур талшығын қосу арқылы
Әр түрлі жастағы егеуқұйрықтардың тамақтану рационына асқабақ пен топинамбур талшығын қосу арқылы Регенера́ция (восстановление)
Регенера́ция (восстановление) Низшие. Плесневые грибы
Низшие. Плесневые грибы Жизнь на Земле
Жизнь на Земле Пищеварительная система
Пищеварительная система Презентация на тему РЕЙТИНГ САМЫХ ОПАСНЫХ ЖИВОТНЫХ В МИРЕ
Презентация на тему РЕЙТИНГ САМЫХ ОПАСНЫХ ЖИВОТНЫХ В МИРЕ  Надкласс рыбы
Надкласс рыбы Основные процессы в корнях
Основные процессы в корнях Першо-звіри. Історія появи тварин
Першо-звіри. Історія появи тварин Характеристика и многообразие рыб
Характеристика и многообразие рыб Тип хордовые. Ланцетник
Тип хордовые. Ланцетник Знатоки природы
Знатоки природы Пищеварение
Пищеварение Основы цитологии
Основы цитологии Формы естественного отбора
Формы естественного отбора Vylučovanie - močová sústava človeka
Vylučovanie - močová sústava človeka Годовой жизненный цикл и сезонные явления в жизни птиц
Годовой жизненный цикл и сезонные явления в жизни птиц Горох и его разновидности
Горох и его разновидности Строение животной клетки
Строение животной клетки СКЕЛЕТНЫЕ МЫШЦЫ
СКЕЛЕТНЫЕ МЫШЦЫ Стеблевые суккуленты
Стеблевые суккуленты Презентация на тему История развития жизни
Презентация на тему История развития жизни  Гетерохронность развития
Гетерохронность развития Боль в спине. Истории пациентов (собак)
Боль в спине. Истории пациентов (собак) Сравнительно анатомические доказательства инволюции
Сравнительно анатомические доказательства инволюции Тип Моллюски или Мягкотелые
Тип Моллюски или Мягкотелые