- Гормоны белковой природы. Лекция 3

Содержание
- 2. Гормоны – биологически активные вещества, которые синтезируются в эндокринных клетках, транспортируются кровью и действуют на ткани
- 3. Классификация и номенклатура гормонов Все гормоны классифицируют по химическому строению, биологическим функциям и механизму действия. 1.
- 5. Все гормоны действуют ограниченное время, которое зависит от скорости их распада. Самое короткое действие имеют пептиды
- 6. Механизм действия гормонов на клетку. Биологическое действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней. Для
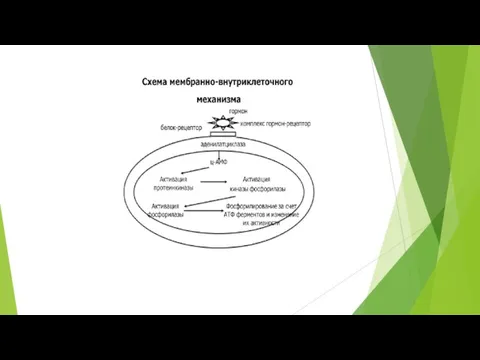
- 7. Активация аденилатциклазы приводит в свою очередь к увеличению образования циклической АМФ, которая служит как бы посредником
- 9. Гормоны поджелудочной железы Поджелудочная железа выполняет в организме две важнейшие функции: экзокринную и эндокринную. Экзокринная функция
- 10. Биосинтез инсулина включает образование двух неактивных предшественников, препроинсулина и проинсулина, которые в результате последовательного протеолиза превращаются
- 11. проинсулин Т1/2 инсулина в плазме крови составляет 3-10 мин, С-пептида - около 30 мин. Схема биосинтеза
- 12. Регуляция синтеза и секреции инсулина. Глюкоза - главный регулятор секреции инсулина, а β-клетки - наиболее важные
- 13. На секрецию инсулина влияют другие гормоны. Адреналин через α2-рецепторы тормозит секрецию инсулина даже на фоне стимуляции
- 14. Биологические функции инсулина Инсулин - главный анаболический гормон. Он участвует в регуляции метаболизма, транспорта глюкозы, аминокислот,
- 15. - заболевание, обусловленное абсолютной или относительной недостаточностью инсулина и нарушением всех видов обмена веществ сахарный диабет
- 16. БИОХИМИЧЕСКИЕ НАРУШЕНИЯ ПРИ САХАРНОМ ДИАБЕТЕ: ГИПЕРГЛЮКОЗЕМИЯ, ГЛЮКОЗУРИЯ; ПОЛИДЕПСИЯ, ПОЛИУРИЯ; КЕТОНЕМИЯ, КЕТОНУРИЯ; ГИПЕРАЗОТЕМИЯ, ГИПЕРАЗОТУРИЯ; МЕТАБОЛИЧЕСКИЙ АЦИДОЗ.
- 17. Глюкагон Глюкагон - одноцепочечный полипептид, состоящий из 29 аминокислотных остатков. Биосинтез глюкагона происходит в α-клетках островков
- 18. Эффекты глюкагона в основном противоположны эффектам инсулина. Основные клетки-мишени глюкагона - печень и жировая ткань. Связываясь
- 20. Синтез и секреция катехоламинов Синтез катехоламинов происходит в цитоплазме и гранулах клеток мозгового слоя надпочечников. В
- 21. Механизм действия и биологические функции катехоламинов Катехоламины действуют на клетки-мишени через рецепторы, локализованные в плазматической мембране.
- 22. Патология мозгового вещества надпочечников Основная патология мозгового вещества надпочечников - феохромоцитома, опухоль, образованная хромаффинными клетками и
- 23. Биосинтез йодтиронинов Йодтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют собой морфологическую и
- 24. БИОСИНТЕЗ ТИРЕОИДНЫХ ГОРМОНОВ Биологическая активность йодтиронинов обусловлена свободной фракцией. Основная биологически активная форма йодтиронинов - Т3;
- 25. Йодирование тиреоглобулина и образование йодтиронинов осуществляется в несколько этапов. Транспорт йода в клетки щитовидной железы. Йод
- 26. Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойодтирозинов (МИТ) и
- 27. йодтиронины УВЕЛИЧИВАЮТ белковый синтез, стимулируют процессы роста и клеточной дифференцировки, ускоряют транскрипцию гена гормона роста. в
- 28. Гипотиреоз -при недостаточности функции щитовидной Гипотериоз приводит к снижению основного обмена, скорости гликолиза, мобилизации гликогена и
- 29. Гормоны гипоталамуса Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС и эндокринные железы.
- 30. 2. Кортиколиберин Кортиколиберин - полипептид, содержащий 41 аминокислотный остаток. Как и другие пептидные гормоны, Кортиколиберин синтезируется
- 31. 5.Соматостатин Соматостатин первично был выделен из гипоталамуса, но впоследствии оказалось, что он синтезируется во многих клетках,
- 32. Гормоны гипофиза Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов и физиологических функций.
- 33. Гормон роста, пролактин Гормон роста синтезируется в соматотропных клетках, наиболее многочисленных в передней доле гипофиза. Содержание
- 34. Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных с ростом и развитием
- 35. У мужчин Пролактин повышает чувствительность клеток Лейдига к лютеинизирующему гормону, поддерживая таким образом необходимый уровень синтеза
- 36. Тиреотропин, лютеинизирующий гормон фолликулостимулирующий гормон Тиреотропин, ЛГ и ФСГ - гликопротеины. Тиреотропин (TIT) синтезируется в тиреотрофных
- 37. В клетках коры надпочечников АКТГ стимулирует гидролиз эфиров холестерола, увеличивает поступление в клетки холестерола в составе
- 38. Оба гормона образуются в гипоталамусе в нейронах разных гипоталамических ядер в форме прогормонов, из которых в
- 39. Нарушения функций гипоталамо-гипофизарной системы Гипофункция может быта следствием уменьшения или полного подавления продукции тройных гормонов (пангипопитуитаризм)
- 41. Скачать презентацию
Слайд 2Гормоны – биологически активные вещества, которые синтезируются в эндокринных клетках, транспортируются кровью
Гормоны – биологически активные вещества, которые синтезируются в эндокринных клетках, транспортируются кровью

Слайд 3Классификация и номенклатура гормонов
Все гормоны классифицируют по химическому строению, биологическим функциям и
Классификация и номенклатура гормонов
Все гормоны классифицируют по химическому строению, биологическим функциям и


Слайд 5Все гормоны действуют ограниченное время, которое зависит от скорости их распада.
Самое короткое
Все гормоны действуют ограниченное время, которое зависит от скорости их распада.
Самое короткое

Слайд 6Механизм действия гормонов на клетку.
Биологическое действие гормонов проявляется через их взаимодействие с
Механизм действия гормонов на клетку.
Биологическое действие гормонов проявляется через их взаимодействие с

Слайд 7Активация аденилатциклазы приводит в свою очередь к увеличению образования циклической АМФ, которая
Активация аденилатциклазы приводит в свою очередь к увеличению образования циклической АМФ, которая

Слайд 9Гормоны поджелудочной железы
Поджелудочная железа выполняет в организме две важнейшие функции: экзокринную
Гормоны поджелудочной железы
Поджелудочная железа выполняет в организме две важнейшие функции: экзокринную

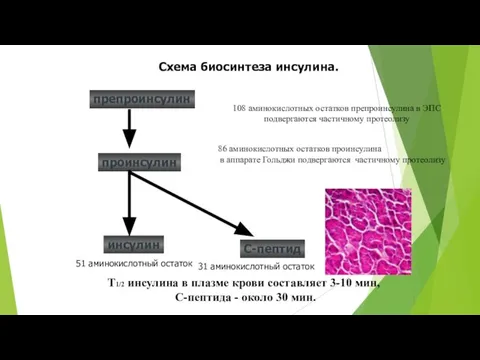
Слайд 10Биосинтез инсулина включает образование двух неактивных предшественников, препроинсулина и проинсулина, которые в результате
Биосинтез инсулина включает образование двух неактивных предшественников, препроинсулина и проинсулина, которые в результате

Слайд 11проинсулин
Т1/2 инсулина в плазме крови составляет 3-10 мин,
С-пептида - около 30
проинсулин
Т1/2 инсулина в плазме крови составляет 3-10 мин,
С-пептида - около 30
Слайд 12Регуляция синтеза и секреции инсулина. Глюкоза - главный регулятор секреции инсулина, а β-клетки
Регуляция синтеза и секреции инсулина. Глюкоза - главный регулятор секреции инсулина, а β-клетки

Слайд 13На секрецию инсулина влияют другие гормоны. Адреналин через α2-рецепторы тормозит секрецию инсулина
На секрецию инсулина влияют другие гормоны. Адреналин через α2-рецепторы тормозит секрецию инсулина

Слайд 14Биологические функции инсулина
Инсулин - главный анаболический гормон. Он участвует в регуляции метаболизма, транспорта
Биологические функции инсулина
Инсулин - главный анаболический гормон. Он участвует в регуляции метаболизма, транспорта

Слайд 15- заболевание, обусловленное абсолютной или относительной недостаточностью инсулина и нарушением всех видов
- заболевание, обусловленное абсолютной или относительной недостаточностью инсулина и нарушением всех видов

Слайд 16БИОХИМИЧЕСКИЕ НАРУШЕНИЯ ПРИ САХАРНОМ ДИАБЕТЕ:
ГИПЕРГЛЮКОЗЕМИЯ, ГЛЮКОЗУРИЯ;
ПОЛИДЕПСИЯ, ПОЛИУРИЯ;
КЕТОНЕМИЯ, КЕТОНУРИЯ;
ГИПЕРАЗОТЕМИЯ, ГИПЕРАЗОТУРИЯ;
МЕТАБОЛИЧЕСКИЙ АЦИДОЗ.
БИОХИМИЧЕСКИЕ НАРУШЕНИЯ ПРИ САХАРНОМ ДИАБЕТЕ:
ГИПЕРГЛЮКОЗЕМИЯ, ГЛЮКОЗУРИЯ;
ПОЛИДЕПСИЯ, ПОЛИУРИЯ;
КЕТОНЕМИЯ, КЕТОНУРИЯ;
ГИПЕРАЗОТЕМИЯ, ГИПЕРАЗОТУРИЯ;
МЕТАБОЛИЧЕСКИЙ АЦИДОЗ.

Слайд 17Глюкагон
Глюкагон - одноцепочечный полипептид, состоящий из 29 аминокислотных остатков. Биосинтез глюкагона происходит
Глюкагон
Глюкагон - одноцепочечный полипептид, состоящий из 29 аминокислотных остатков. Биосинтез глюкагона происходит

Слайд 18Эффекты глюкагона в основном противоположны эффектам инсулина. Основные клетки-мишени глюкагона - печень и
Эффекты глюкагона в основном противоположны эффектам инсулина. Основные клетки-мишени глюкагона - печень и


Слайд 20 Синтез и секреция катехоламинов
Синтез катехоламинов происходит в цитоплазме и гранулах клеток
Синтез и секреция катехоламинов
Синтез катехоламинов происходит в цитоплазме и гранулах клеток

Слайд 21Механизм действия и биологические функции катехоламинов
Катехоламины действуют на клетки-мишени через рецепторы, локализованные
Механизм действия и биологические функции катехоламинов
Катехоламины действуют на клетки-мишени через рецепторы, локализованные

Слайд 22Патология мозгового вещества надпочечников
Основная патология мозгового вещества надпочечников - феохромоцитома, опухоль, образованная
Патология мозгового вещества надпочечников
Основная патология мозгового вещества надпочечников - феохромоцитома, опухоль, образованная

Слайд 23Биосинтез йодтиронинов
Йодтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют
Биосинтез йодтиронинов
Йодтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют

Слайд 24БИОСИНТЕЗ ТИРЕОИДНЫХ ГОРМОНОВ
Биологическая активность йодтиронинов обусловлена свободной фракцией.
Основная биологически активная форма
БИОСИНТЕЗ ТИРЕОИДНЫХ ГОРМОНОВ
Биологическая активность йодтиронинов обусловлена свободной фракцией.
Основная биологически активная форма

Слайд 25Йодирование тиреоглобулина и образование йодтиронинов осуществляется в несколько этапов.
Транспорт йода в клетки
Йодирование тиреоглобулина и образование йодтиронинов осуществляется в несколько этапов.
Транспорт йода в клетки

Слайд 26Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием
Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием

Слайд 27йодтиронины УВЕЛИЧИВАЮТ
белковый синтез,
стимулируют процессы роста и клеточной дифференцировки,
ускоряют транскрипцию
йодтиронины УВЕЛИЧИВАЮТ
белковый синтез,
стимулируют процессы роста и клеточной дифференцировки,
ускоряют транскрипцию

Слайд 28Гипотиреоз -при недостаточности функции щитовидной
Гипотериоз приводит к снижению основного обмена, скорости
Гипотиреоз -при недостаточности функции щитовидной
Гипотериоз приводит к снижению основного обмена, скорости

Слайд 29Гормоны гипоталамуса
Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС
Гормоны гипоталамуса
Гипоталамус занимает важнейшее место в иерархической системе, объединяя высшие отделы ЦНС

Слайд 302. Кортиколиберин
Кортиколиберин - полипептид, содержащий 41 аминокислотный остаток. Как и другие пептидные
2. Кортиколиберин
Кортиколиберин - полипептид, содержащий 41 аминокислотный остаток. Как и другие пептидные

Слайд 315.Соматостатин
Соматостатин первично был выделен из гипоталамуса, но впоследствии оказалось, что он синтезируется
5.Соматостатин
Соматостатин первично был выделен из гипоталамуса, но впоследствии оказалось, что он синтезируется

Слайд 32Гормоны гипофиза
Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов
Гормоны гипофиза
Гипофиз секретирует большое количество гормонов, участвующих в регуляции различных биохимических процессов

Слайд 33 Гормон роста, пролактин
Гормон роста синтезируется в соматотропных клетках, наиболее многочисленных в
Гормон роста, пролактин
Гормон роста синтезируется в соматотропных клетках, наиболее многочисленных в

Слайд 34Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных
Основное действие гормона роста направлено на регуляцию обмена белков и процессов, связанных

Слайд 35У мужчин Пролактин повышает чувствительность клеток Лейдига к лютеинизирующему гормону, поддерживая таким
У мужчин Пролактин повышает чувствительность клеток Лейдига к лютеинизирующему гормону, поддерживая таким

Слайд 36Тиреотропин, лютеинизирующий гормон фолликулостимулирующий гормон
Тиреотропин, ЛГ и ФСГ - гликопротеины. Тиреотропин (TIT)
Тиреотропин, лютеинизирующий гормон фолликулостимулирующий гормон
Тиреотропин, ЛГ и ФСГ - гликопротеины. Тиреотропин (TIT)

Слайд 37В клетках коры надпочечников АКТГ стимулирует гидролиз эфиров холестерола, увеличивает поступление в
В клетках коры надпочечников АКТГ стимулирует гидролиз эфиров холестерола, увеличивает поступление в

Слайд 38Оба гормона образуются в гипоталамусе в нейронах разных гипоталамических ядер в форме
Оба гормона образуются в гипоталамусе в нейронах разных гипоталамических ядер в форме

Слайд 39Нарушения функций гипоталамо-гипофизарной системы
Гипофункция может быта следствием уменьшения или полного подавления продукции тройных
Нарушения функций гипоталамо-гипофизарной системы
Гипофункция может быта следствием уменьшения или полного подавления продукции тройных

 Растительный и животный мир Башкортостана
Растительный и животный мир Башкортостана Моллюски в циклах развития паразитов
Моллюски в циклах развития паразитов Строение эукариотических клеток
Строение эукариотических клеток Пищеварение в желудке и 12-перстной кишке
Пищеварение в желудке и 12-перстной кишке Презентация на тему История красной книги
Презентация на тему История красной книги  Причины акселерации
Причины акселерации Рыба язь
Рыба язь Коровы
Коровы Презентация на тему Основные направления эволюции
Презентация на тему Основные направления эволюции  Влияние поллютантов на возникновение мутаций у бактерий
Влияние поллютантов на возникновение мутаций у бактерий Регуляция метаболизма у бактерий
Регуляция метаболизма у бактерий Характер строения и окраска ствола и ветвей
Характер строения и окраска ствола и ветвей Безусловные рефлексы
Безусловные рефлексы Эволюция органического мира
Эволюция органического мира Фармакология процессов обмена веществ и системы крови
Фармакология процессов обмена веществ и системы крови Организм человека, как единая биологическая система
Организм человека, как единая биологическая система Наследственность и изменчивость. Роль наследственности и изменчивости в эволюции
Наследственность и изменчивость. Роль наследственности и изменчивости в эволюции Устройство микроскопа
Устройство микроскопа Строение и функции головного мозга
Строение и функции головного мозга Ландыш майский
Ландыш майский Пуповина (пупочный канатик)
Пуповина (пупочный канатик) Особенности внутреннего строения рыб (часть 2)
Особенности внутреннего строения рыб (часть 2) Современные методы палеонтологических исследований
Современные методы палеонтологических исследований Кожа и её производные
Кожа и её производные Бром и его биологическое значение
Бром и его биологическое значение Что такое метаболизм?
Что такое метаболизм? Познавательные процессы
Познавательные процессы Презентация на тему Железо внутри нас
Презентация на тему Железо внутри нас