- Математическое моделирование динамики молекулярно-генетических систем

Содержание
- 2. Что такое модель? Модель, в широком смысле любой образ, аналог (мысленный или условный: изображение, описание, схема,
- 3. Что такое моделирование? Моделирование - исследование каких-либо явлений, процессов или систем объектов путем построения и изучения
- 4. Для чего нужна модель? Для понимания строения изучаемого объекта: его структуры, основных свойств, законов развития и
- 5. Приемы и способы моделирования Моделирование Материальное (предметное) Способы моделирования, при которых исследование ведется на основе модели,
- 6. При разработке любой модели необходимо определить объект моделирования, цель моделирования и средства моделирования. В соответствии с
- 7. непрерывные - используют аппарат непрерывной математики, дифференциальных, интегро-дифференциальных уравнений стохастические -основаны на применении вероятностных методов логические
- 8. Процесс построения моделей Формируются основные вопросы о поведении системы, ответы на которые планируется получить с помощью
- 9. Фундаментальная проблема наук о жизни Одной из центральных проблем постгеномной молекулярной биологии и генетики является исследование
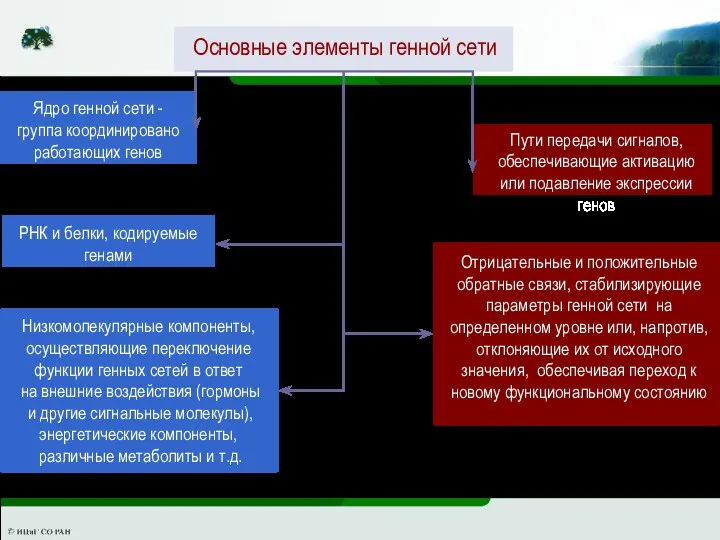
- 10. Генные сети как объект исследования Генная сеть – группа координировано функционирующих генов, контролирующих физиологические, биохимические, молекулярные
- 12. Специфика объекта моделирования Типичные генетические системы состоят из большого количества компонентов различной природы и структурно-функциональной организации,
- 13. Специфика объекта моделирования Биологические системы является самовоспроизводящимися динамическими системами. Особенностью функционирования биологических систем во времени является
- 14. Логические подходы Логические подходы основаны на применении для описания механизмов в терминах логики и дискретной математики.
- 15. Логические подходы X Z Y X(следующее) = NO Z(текущее) Y(следующее) = NO X(текущее) Z(следующее) = NO
- 16. Стохастическое моделирование В стохастических моделях молекулярные события описываются как случайные процессы. В них каждое элементарное событие
- 17. Стохастическое моделирование В стохастическом подходе с непрерывным временем считается, что система из текущего момента времени переходит
- 18. Непрерывные подходы к описанию биологических систем Динамика молекулярно-генетических систем в непрерывных моделях формально описывается системами дифференциальных
- 19. Иерархический метод моделирования ИММ использует в качестве основного подхода принцип иерархической декомпозиции целевой биологической системы до
- 20. Элементы системы (компарт- менты, клетки, органы и т.д.) Блок схема моделирования
- 21. Какие подсистемы считаются элементарными? Элементарной подсистемой моделируемой системы считается подсистема, которая может быть рассмотрена автономно, вне
- 22. Примерный список элементарных подсистем генных сетей. При этом следует помнить, что обычно элементарные подсистемы с биохимической
- 23. В таблице мы привели примерный список элементарных подсистем. Список не претендует на полноту. Не все приведенные
- 24. Общий вид элементарных моделей X - вектор управляемых переменных, Y - вектор управляющих переменных, K –
- 25. Методы реконструкции моделей элементарных подсистем
- 26. Биохимическая природа молекулярно-генетических процессов - основа для вывода функции V Фундаментальной основой для вывода функции V
- 27. Кинетический закон действующих масс выводится на основе теории соударений. Пусть имеется биохимическая реакция A + B
- 28. Если имеется конкретная биохимическая схема реакций вида (1), то мгновенная скорость изменения концентрации любого вещества равняется
- 29. Теорема Корзухина – теоретическая основа для применения химико-кинетического подхода к моделированию молекулярно-генетических систем (не строгая формулировка)
- 30. Значение теоремы Корзухина. При использовании химико-кинетическиого подхода V является полиномом степени не выше двух. Однако на
- 31. Теорема Тихонова Пусть дана система дифференциальных автономных уравнений
- 32. Теорема Тихонова
- 33. Теорема Тихонова позволяет понижать порядок системы дифференциальных уравнений. Для того чтобы применить этот метод к биохимической
- 34. Формула Михаэлиса-Ментен Простейшие преобразования и переход к исходным обозначениям дает знаменитую формулу Пример применения теоремы Тихонова
- 35. Разделение процессов на быстрые и медленные. Как уже говорилось, общего алгоритма разделения переменных на быстрые и
- 36. Квазистационарный метод исключения переменных (метод Кинга-Альтмана)
- 37. Важный частный случай применения принципа квазистационарности для исследования механизмов ферментативных реакций Основу принципа квазистационарности составляет рассмотрение
- 38. Пусть имеется фермент E, который может пребывать в n состояниях: ei, i=1,…,n. Пусть из i-го состояния
- 39. Метод Кинга-Альтмана
- 40. Метод Кинга-Альтмана
- 41. Равновесный подход к моделированию функций промоторов
- 42. Равновесный подход к моделированию функций промоторов g gA gP gR gAR gAP
- 43. Приравняем к нулю правую часть системы и учтем уравнение баланса Равновесный подход к моделированию функций промоторов
- 44. Итоговые формулы элементарной модели функционирования гипотетического промотора Равновесный подход к моделированию функций промоторов
- 45. Аппроксимация механизмов в терминах обобщенных функций Хилла Теперь перейдем к рассмотрению подхода, который используется для моделирования
- 46. Аппроксимация механизмов в терминах обобщенных функций Хилла ОФХ являются естественным расширением класса рациональных полиномов, которые возникают
- 47. Аппроксимация механизмов в терминах обобщенных функций Хилла Регуляция экспрессии гена pyrC в клетке E.coli Экспрессия гена,
- 48. Аппроксимация механизмов в терминах обобщенных функций Хилла Регуляция экспрессии гена pyrC в клетке E.coli Таким образом,
- 49. Аппроксимация механизмов в терминах обобщенных функций Хилла
- 50. Аппроксимация механизмов в терминах обобщенных функций Хилла Модель регуляции экспрессии оперона cyoABCDE Структура промотора оперона cyoABCDE,
- 51. Аппроксимация механизмов в терминах обобщенных функций Хилла Регуляция экспрессии гена pyrC в клетке E.coli
- 52. Аппроксимация механизмов в терминах обобщенных функций Хилла: Модель регуляции экспрессии оперона cyoABCDE Зависимость уровня транскрипции оперона
- 53. Аппроксимация механизмов в терминах обобщенных функций Хилла Сделаем несколько комментариев к рассмотренной модели экспрессии оперона cydAB.
- 54. 1. Рациональный полином 3. Функция, которая может быть получена из ОФХ, путем подстановки в нее вместо
- 55. A ⎯→ B, a = [A], b = [B] k -da/db = db/dt = ka e=[E],
- 56. Закон суммирования локальных скоростей элементарных подсистем
- 57. Закон суммирования локальных скоростей элементарных подсистем
- 59. Скачать презентацию

Слайд 3Что такое моделирование?
Моделирование - исследование каких-либо явлений, процессов или систем объектов путем
Что такое моделирование?
Моделирование - исследование каких-либо явлений, процессов или систем объектов путем

Слайд 4Для чего нужна модель?
Для понимания строения изучаемого объекта: его структуры, основных свойств,
Для чего нужна модель?
Для понимания строения изучаемого объекта: его структуры, основных свойств,

Слайд 5Приемы и способы моделирования
Моделирование
Материальное (предметное)
Способы моделирования, при которых исследование ведется на основе
Приемы и способы моделирования
Моделирование
Материальное (предметное)
Способы моделирования, при которых исследование ведется на основе

Слайд 6При разработке любой модели необходимо определить объект моделирования, цель моделирования и средства
При разработке любой модели необходимо определить объект моделирования, цель моделирования и средства

Слайд 7 непрерывные - используют аппарат непрерывной математики, дифференциальных, интегро-дифференциальных уравнений
стохастические -основаны
непрерывные - используют аппарат непрерывной математики, дифференциальных, интегро-дифференциальных уравнений
стохастические -основаны

Слайд 8Процесс построения моделей
Формируются основные вопросы о поведении системы, ответы на которые планируется
Процесс построения моделей
Формируются основные вопросы о поведении системы, ответы на которые планируется

Слайд 9Фундаментальная проблема наук о жизни
Одной из центральных проблем постгеномной молекулярной биологии и
Фундаментальная проблема наук о жизни
Одной из центральных проблем постгеномной молекулярной биологии и

Слайд 10Генные сети как объект исследования
Генная сеть – группа координировано функционирующих
генов, контролирующих физиологические,
Генные сети как объект исследования
Генная сеть – группа координировано функционирующих
генов, контролирующих физиологические,

Слайд 12Специфика объекта моделирования
Типичные генетические системы состоят из большого количества компонентов различной природы
Специфика объекта моделирования
Типичные генетические системы состоят из большого количества компонентов различной природы

Слайд 13Специфика объекта моделирования
Биологические системы является самовоспроизводящимися динамическими системами.
Особенностью функционирования биологических систем во
Специфика объекта моделирования
Биологические системы является самовоспроизводящимися динамическими системами.
Особенностью функционирования биологических систем во

Слайд 14Логические подходы
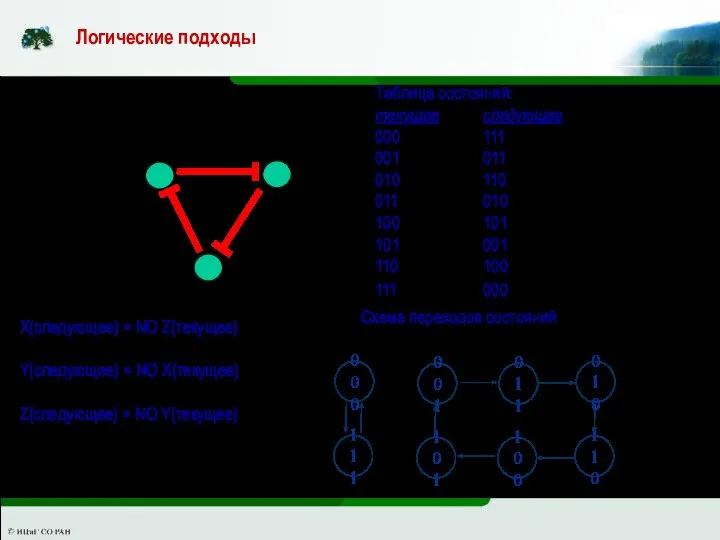
Логические подходы основаны на применении для описания механизмов в терминах логики
Логические подходы
Логические подходы основаны на применении для описания механизмов в терминах логики

Слайд 15Логические подходы
X
Z
Y
X(следующее) = NO Z(текущее)
Y(следующее) = NO X(текущее)
Z(следующее) = NO Y(текущее)
Таблица состояний:
текущее следующее
000 111
001 011
010 110
011 010
100 101
101 001
110 100
111 000
Логические подходы
X
Z
Y
X(следующее) = NO Z(текущее)
Y(следующее) = NO X(текущее)
Z(следующее) = NO Y(текущее)
Таблица состояний:
текущее следующее
000 111
001 011
010 110
011 010
100 101
101 001
110 100
111 000
Слайд 16Стохастическое моделирование
В стохастических моделях молекулярные события описываются как случайные процессы. В них
Стохастическое моделирование
В стохастических моделях молекулярные события описываются как случайные процессы. В них

Слайд 17Стохастическое моделирование
В стохастическом подходе с непрерывным временем считается, что система из текущего
Стохастическое моделирование
В стохастическом подходе с непрерывным временем считается, что система из текущего

Слайд 18Непрерывные подходы к описанию биологических систем
Динамика молекулярно-генетических систем в непрерывных моделях
Непрерывные подходы к описанию биологических систем
Динамика молекулярно-генетических систем в непрерывных моделях

Слайд 19Иерархический метод моделирования
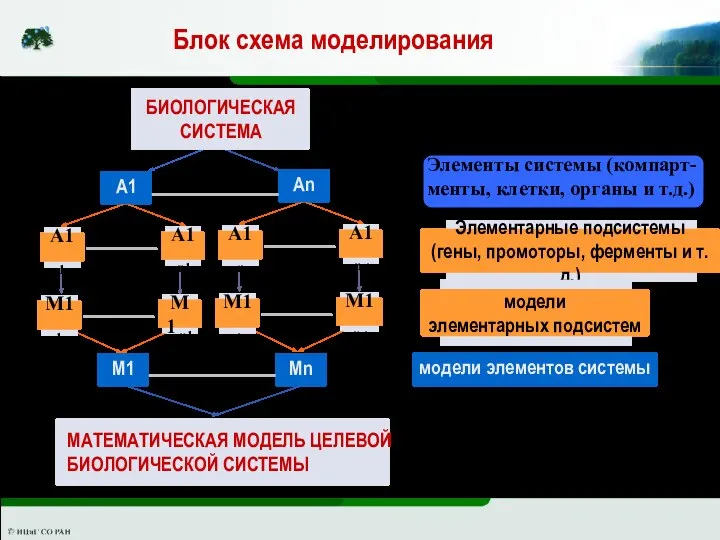
ИММ использует в качестве основного подхода принцип иерархической декомпозиции целевой
Иерархический метод моделирования
ИММ использует в качестве основного подхода принцип иерархической декомпозиции целевой

Слайд 20Элементы системы (компарт- менты, клетки, органы и т.д.)
Блок схема моделирования
Элементы системы (компарт- менты, клетки, органы и т.д.)
Блок схема моделирования
Слайд 21Какие подсистемы считаются элементарными?
Элементарной подсистемой моделируемой системы считается подсистема, которая может быть
Какие подсистемы считаются элементарными?
Элементарной подсистемой моделируемой системы считается подсистема, которая может быть

Слайд 22Примерный список элементарных подсистем генных сетей.
При этом следует помнить, что обычно элементарные
Примерный список элементарных подсистем генных сетей.
При этом следует помнить, что обычно элементарные

Слайд 23В таблице мы привели примерный список элементарных подсистем. Список не претендует на
В таблице мы привели примерный список элементарных подсистем. Список не претендует на

Слайд 24Общий вид элементарных моделей
X - вектор управляемых переменных, Y - вектор управляющих
Общий вид элементарных моделей
X - вектор управляемых переменных, Y - вектор управляющих

Слайд 25Методы реконструкции моделей элементарных подсистем
Методы реконструкции моделей элементарных подсистем

Слайд 26Биохимическая природа молекулярно-генетических процессов - основа для вывода функции V
Фундаментальной основой
Биохимическая природа молекулярно-генетических процессов - основа для вывода функции V
Фундаментальной основой

Слайд 27Кинетический закон действующих масс выводится на основе теории соударений.
Пусть имеется биохимическая
Кинетический закон действующих масс выводится на основе теории соударений.
Пусть имеется биохимическая

Слайд 28Если имеется конкретная биохимическая схема реакций вида (1), то мгновенная скорость изменения
Если имеется конкретная биохимическая схема реакций вида (1), то мгновенная скорость изменения

Слайд 29Теорема Корзухина – теоретическая основа для применения химико-кинетического подхода к моделированию молекулярно-генетических
Теорема Корзухина – теоретическая основа для применения химико-кинетического подхода к моделированию молекулярно-генетических
Слайд 30Значение теоремы Корзухина.
При использовании химико-кинетическиого подхода V является полиномом степени не выше
Значение теоремы Корзухина.
При использовании химико-кинетическиого подхода V является полиномом степени не выше

Слайд 31Теорема Тихонова
Пусть дана система дифференциальных автономных уравнений
Теорема Тихонова
Пусть дана система дифференциальных автономных уравнений

Слайд 32Теорема Тихонова
Теорема Тихонова
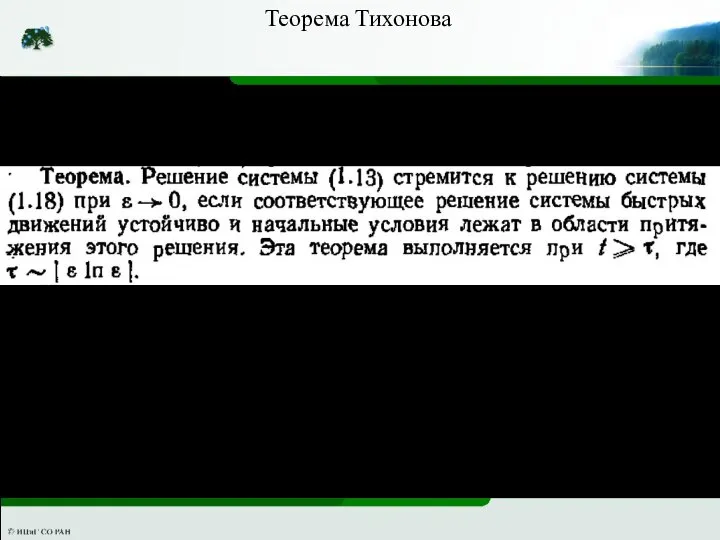
Слайд 33Теорема Тихонова позволяет понижать порядок системы дифференциальных уравнений. Для того чтобы применить
Теорема Тихонова позволяет понижать порядок системы дифференциальных уравнений. Для того чтобы применить

Слайд 34Формула Михаэлиса-Ментен
Простейшие преобразования и переход к исходным обозначениям дает знаменитую формулу
Пример
Формула Михаэлиса-Ментен
Простейшие преобразования и переход к исходным обозначениям дает знаменитую формулу
Пример

Слайд 35Разделение процессов на быстрые и медленные.
Как уже говорилось, общего алгоритма разделения переменных
Разделение процессов на быстрые и медленные.
Как уже говорилось, общего алгоритма разделения переменных

Слайд 36Квазистационарный метод исключения переменных
(метод Кинга-Альтмана)
Квазистационарный метод исключения переменных
(метод Кинга-Альтмана)

Слайд 37Важный частный случай применения принципа квазистационарности для исследования механизмов ферментативных реакций
Основу принципа
Важный частный случай применения принципа квазистационарности для исследования механизмов ферментативных реакций
Основу принципа

Слайд 38Пусть имеется фермент E, который может пребывать в n состояниях: ei, i=1,…,n.
Пусть имеется фермент E, который может пребывать в n состояниях: ei, i=1,…,n.

Слайд 39Метод Кинга-Альтмана
Метод Кинга-Альтмана

Слайд 40Метод Кинга-Альтмана
Метод Кинга-Альтмана

Слайд 41Равновесный подход к моделированию функций промоторов
Равновесный подход к моделированию функций промоторов
Слайд 42Равновесный подход к моделированию функций промоторов
g
gA
gP
gR
gAR
gAP
Равновесный подход к моделированию функций промоторов
g
gA
gP
gR
gAR
gAP
Слайд 43Приравняем к нулю правую часть системы и учтем уравнение баланса
Равновесный подход к
Приравняем к нулю правую часть системы и учтем уравнение баланса
Равновесный подход к

Слайд 44Итоговые формулы элементарной модели функционирования гипотетического промотора
Равновесный подход к моделированию функций промоторов
Итоговые формулы элементарной модели функционирования гипотетического промотора
Равновесный подход к моделированию функций промоторов

Слайд 45Аппроксимация механизмов в терминах обобщенных функций Хилла
Теперь перейдем к рассмотрению подхода, который
Аппроксимация механизмов в терминах обобщенных функций Хилла
Теперь перейдем к рассмотрению подхода, который

Слайд 46Аппроксимация механизмов в терминах обобщенных функций Хилла
ОФХ являются естественным расширением класса рациональных
Аппроксимация механизмов в терминах обобщенных функций Хилла
ОФХ являются естественным расширением класса рациональных

Слайд 47Аппроксимация механизмов в терминах обобщенных функций Хилла
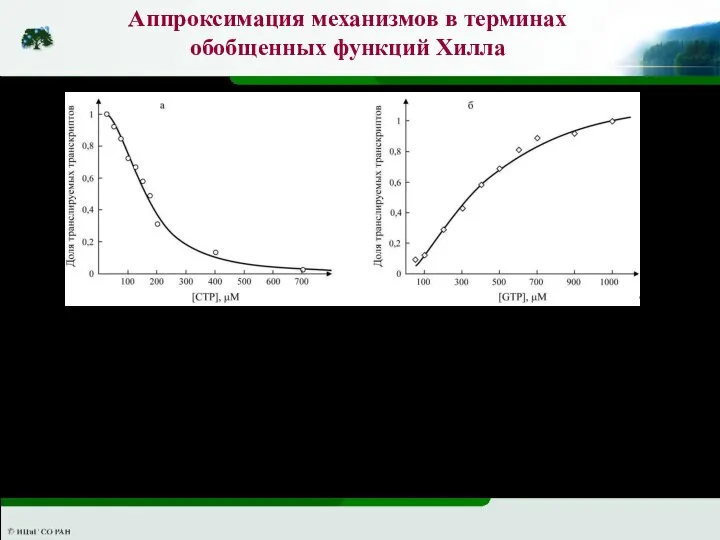
Регуляция экспрессии гена pyrC в клетке
Аппроксимация механизмов в терминах обобщенных функций Хилла
Регуляция экспрессии гена pyrC в клетке

Слайд 48Аппроксимация механизмов в терминах обобщенных функций Хилла
Регуляция экспрессии гена pyrC в клетке
Аппроксимация механизмов в терминах обобщенных функций Хилла
Регуляция экспрессии гена pyrC в клетке

Слайд 49Аппроксимация механизмов в терминах обобщенных функций Хилла
Аппроксимация механизмов в терминах обобщенных функций Хилла
Слайд 50Аппроксимация механизмов в терминах обобщенных функций Хилла
Модель регуляции экспрессии оперона cyoABCDE
Структура промотора
Аппроксимация механизмов в терминах обобщенных функций Хилла
Модель регуляции экспрессии оперона cyoABCDE
Структура промотора

Слайд 51Аппроксимация механизмов в терминах обобщенных функций Хилла
Регуляция экспрессии гена pyrC в клетке
Аппроксимация механизмов в терминах обобщенных функций Хилла
Регуляция экспрессии гена pyrC в клетке

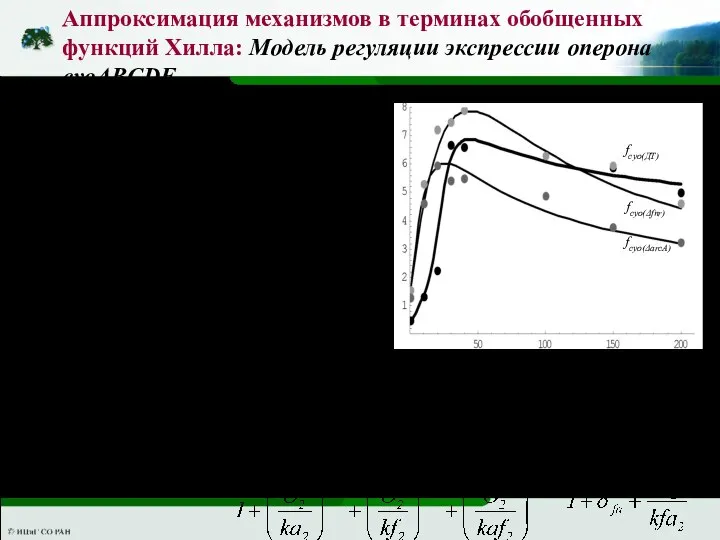
Слайд 52Аппроксимация механизмов в терминах обобщенных функций Хилла: Модель регуляции экспрессии оперона cyoABCDE
Зависимость
Аппроксимация механизмов в терминах обобщенных функций Хилла: Модель регуляции экспрессии оперона cyoABCDE
Зависимость
Слайд 53Аппроксимация механизмов в терминах обобщенных функций Хилла
Сделаем несколько комментариев к рассмотренной модели
Аппроксимация механизмов в терминах обобщенных функций Хилла
Сделаем несколько комментариев к рассмотренной модели

Слайд 541. Рациональный полином
3. Функция, которая может быть получена из ОФХ, путем
1. Рациональный полином
3. Функция, которая может быть получена из ОФХ, путем

Слайд 55A ⎯→ B, a = [A], b = [B]
k
-da/db = db/dt =
A ⎯→ B, a = [A], b = [B]
k
-da/db = db/dt =
![A ⎯→ B, a = [A], b = [B] k -da/db =](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/950867/slide-54.jpg)
Слайд 56Закон суммирования локальных скоростей элементарных подсистем
Закон суммирования локальных скоростей элементарных подсистем

Слайд 57Закон суммирования локальных скоростей элементарных подсистем
Закон суммирования локальных скоростей элементарных подсистем

 Насекомые вредители на станции юных натуралистов г. Тайшета
Насекомые вредители на станции юных натуралистов г. Тайшета Рыбы и насекомые пресных водоемов
Рыбы и насекомые пресных водоемов Грибы, цветы и ягоды леса
Грибы, цветы и ягоды леса Чей хвостик
Чей хвостик Семейство крестоцветных
Семейство крестоцветных Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба
Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба Неклеточные формы жизни
Неклеточные формы жизни Раселение живых организмов
Раселение живых организмов Эпителий
Эпителий Презентация на тему Строение и химический состав клетки
Презентация на тему Строение и химический состав клетки  MOLEKUlyarnaya_biologia_LPZ_1
MOLEKUlyarnaya_biologia_LPZ_1 Фотодыхание
Фотодыхание Вирусология - наука о вирусах
Вирусология - наука о вирусах Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Чабрец (богородская трава, тимьян ползучий)
Чабрец (богородская трава, тимьян ползучий) Составление генетических паспортов и установление родственных связей
Составление генетических паспортов и установление родственных связей От клюва до хвоста
От клюва до хвоста Модификационная изменчивость
Модификационная изменчивость Строение и многообразие грибов
Строение и многообразие грибов Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности
Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности  Что было на Земле, когда она появилась?
Что было на Земле, когда она появилась? Биохимия. Разделы биохимии. Становление биохимии как науки
Биохимия. Разделы биохимии. Становление биохимии как науки История собаководства
История собаководства Морфология бактерий
Морфология бактерий Жизненная Ёмкость Лёгких
Жизненная Ёмкость Лёгких Ткани животных
Ткани животных 2. Урок - .ДНК
2. Урок - .ДНК Лишайники
Лишайники