- Молекулярно-генетический уровень жизни. Репликация и репарация

Содержание
- 2. Реализация фундаментальных свойств живых организмов – размножения и роста – на молекулярно-генетическом уровне ! РЕПЛИКАЦИЯ !
- 3. Репликация Репликация – удвоение молекулы ДНК Возможные способы репликации консервативный полуконсервативный М. Мезельсон, Ф. Сталь; 1957
- 4. Репликация. Этапы Подготовительный этап Собственно репликация: Инициация Элонгация Терминация
- 5. Ферменты подготовительного этапа: ДНК-гираза (Топоизомераза I) разрешает топологические проблемы, связанные со спирализацией–деспирализацией ДНК, оборачивается вокруг ДНК
- 6. Репликация
- 7. Начало репликации ВСЕГДА начинается с определенного участка – сайта инициации = ori (от англ. origin –
- 8. Особенности репликации эукариот: меньшая скорость репликации образование сразу нескольких репликационных «вилок»
- 9. ИНИЦИАЦИЯ РЕПЛИКАЦИИ Построение новых цепей на матрице старых осуществляется ДНК-полимеразой. Она неспособна начать синтез новой цепи
- 10. Элонгация репликации Построение новых цепей на матрице старых осуществляется ДНК-полимеразой. Способна осуществлять сборку полинуклеотида в направлении
- 11. Ведущая цепь синтезируется в направлении от 5' к 3' концу (при этом синтез на родительской цепи
- 12. Происходит при достижении репликационной вилкой теломер (концевой участок хоромосом). Теломеры состоят из повторяющихся нуклеотидных участков и
- 13. Терминация репликации Теломераза участвует в репликации теломерных (концевых) участков линейной ДНК. ДНК-полимеразная система, описанная выше, оставляет
- 14. Терминация репликации
- 15. Репарация устранение из ДНК возникающих в ней повреждений, восстановление исходной (нативной) структуры молекулы ДНК-полимеразы
- 16. Репарационные системы: простые (одноэтапные) – фотореактивация, деалкилирование; сложные (многоэтапные, многокомпонентные). Основная цель этих систем – повышение
- 17. Типы повреждений ДНК 1. Спонтанные повреждения: ошибки репликации (появление некомплементарных пар нуклеотидов); апуринизация (отщепление азотистых оснований
- 18. Типы повреждений ДНК 2. Индуцированные повреждения: димеризация (сшивание соседних пиримидиновых оснований с образованием димера); разрывы в
- 19. . Типы повреждений ДНК
- 20. С позиций молекулярного механизма первичные повреждения в молекулах ДНК могут быть устранены тремя путями: прямым возвращением
- 21. По отношению к процессу репликации различают два основные типа репарации ДНК: дорепликативную (фотореактивация и эксцизионная форма);
- 22. Этот тип репарации обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения. Реакциями прямой репарации являются:
- 23. Репарация ДНК (фотореактивация) 1. Нормальная молекула ДНК ↓ Облучение УФ-светом ↓ 2. Мутантная молекула ДНК –
- 24. Репарация ДНК (фотореактивация)
- 25. Репарация ДНК при алкилирующих повреждениях Генетические повреждения, вызываемые присоединением алкильных или метильных групп, могут репарироваться в
- 26. Репарация ДНК – прямая вставка пуринов При некоторых типах повреждений пуриновых оснований ковалентная связь между основанием
- 27. Репарация ДНК при однонитевых разрывах Например, под действием ионизирующего облучения могут возникнуть однонитевые разрывы ДНК. Фермент
- 28. Репарация ДНК (эксцизионная) Этот сложные реакции восстановления, когда поврежденные участки вырезаются из цепи ДНК (excision –
- 29. Репарация ДНК (mismatch repair, MMR) Во время репликации ДНК происходят ошибки спаривания, в результате которых вместо
- 30. Специальные ферменты метилазы присоединяют метильные группы к аденинам в последовательности ГАТЦ на материнскую цепь и она
- 31. Пострепликативная репарация Осуществляется в тех случаях, когда повреждение доживает до фазы репликации или имеет такую природу,
- 32. Рекомбинационная репарация
- 33. Репарация ДНК (SOS-система) Обнаружена в 1974 г. М.Радманом. Включается тогда, когда повреждений в ДНК настолько много,
- 34. Репарация ДНК (SOS-система)
- 35. Репарация ДНК и наследственные болезни человека Нарушение системы репарации у человека является причиной: преждевременного старения; онкозаболеваний
- 36. Прогерия детей – синдром Хатчинсона-Гилфорда маленький рост птичье лицо с клювообразным профилем преобладание мозгового черепа над
- 37. Прогерия взрослых- синдром Вернера Симптомы старения проявляются после полового созревания: Седина, выпадение волос, морщины, гиперпигментация, сухость,
- 39. Скачать презентацию
Слайд 2Реализация фундаментальных свойств живых организмов – размножения и роста – на молекулярно-генетическом
Реализация фундаментальных свойств живых организмов – размножения и роста – на молекулярно-генетическом

Слайд 3Репликация
Репликация – удвоение молекулы ДНК
Возможные способы репликации
консервативный
полуконсервативный
М. Мезельсон,
Ф. Сталь; 1957 г.
дисперсный
Репликация
Репликация – удвоение молекулы ДНК
Возможные способы репликации
консервативный
полуконсервативный
М. Мезельсон,
Ф. Сталь; 1957 г.
дисперсный

Слайд 4Репликация. Этапы
Подготовительный этап
Собственно репликация:
Инициация
Элонгация
Терминация
Репликация. Этапы
Подготовительный этап
Собственно репликация:
Инициация
Элонгация
Терминация

Слайд 5Ферменты подготовительного этапа:
ДНК-гираза (Топоизомераза I) разрешает топологические проблемы, связанные со спирализацией–деспирализацией ДНК,
Ферменты подготовительного этапа:
ДНК-гираза (Топоизомераза I) разрешает топологические проблемы, связанные со спирализацией–деспирализацией ДНК,

Слайд 6Репликация
Репликация
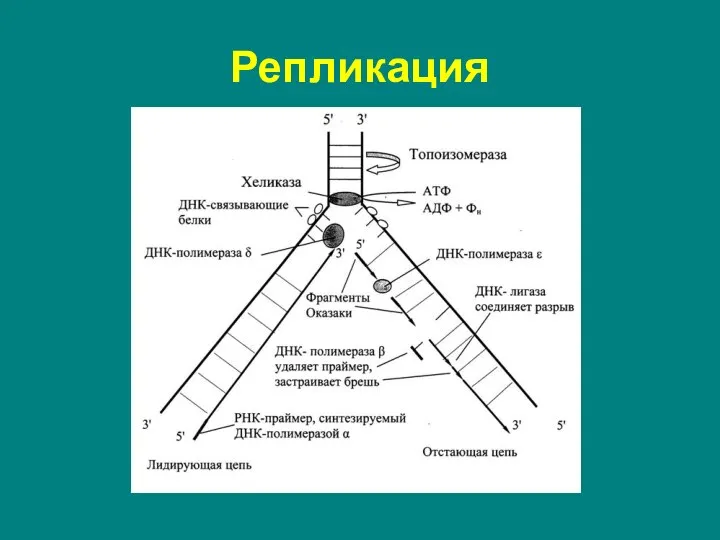
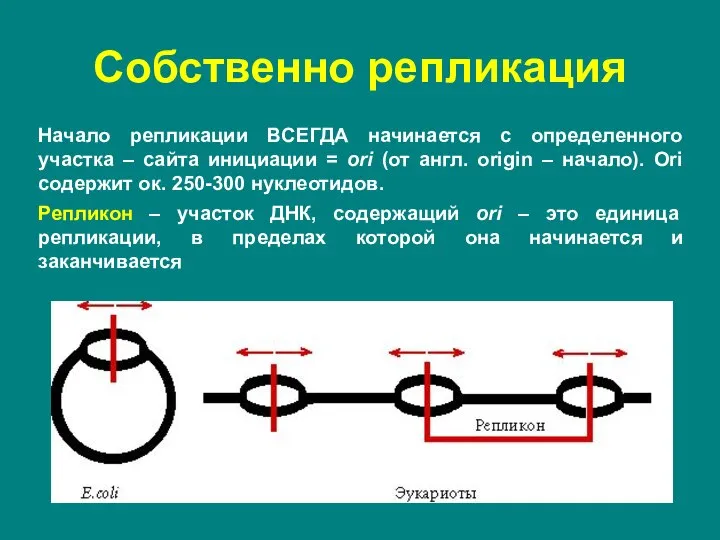
Слайд 7Начало репликации ВСЕГДА начинается с определенного участка – сайта инициации = ori
Начало репликации ВСЕГДА начинается с определенного участка – сайта инициации = ori
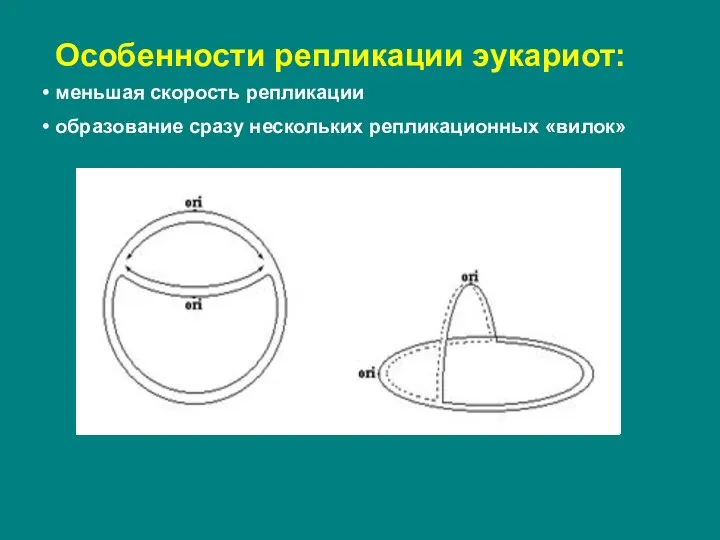
Слайд 8Особенности репликации эукариот:
меньшая скорость репликации
образование сразу нескольких репликационных «вилок»
Особенности репликации эукариот:
меньшая скорость репликации
образование сразу нескольких репликационных «вилок»
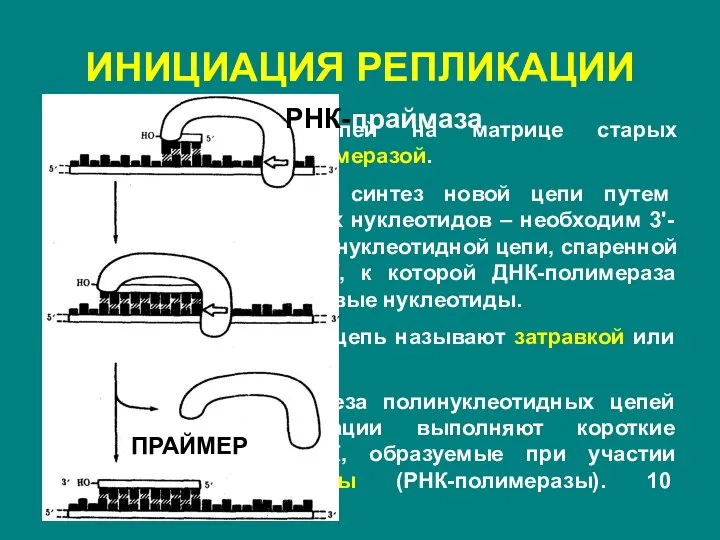
Слайд 9ИНИЦИАЦИЯ РЕПЛИКАЦИИ
Построение новых цепей на матрице старых осуществляется ДНК-полимеразой.
Она неспособна начать
ИНИЦИАЦИЯ РЕПЛИКАЦИИ
Построение новых цепей на матрице старых осуществляется ДНК-полимеразой.
Она неспособна начать
Слайд 10Элонгация репликации
Построение новых цепей на матрице старых осуществляется ДНК-полимеразой.
Способна осуществлять сборку полинуклеотида
Элонгация репликации
Построение новых цепей на матрице старых осуществляется ДНК-полимеразой.
Способна осуществлять сборку полинуклеотида

Слайд 11Ведущая цепь синтезируется в направлении от 5' к 3' концу (при этом
Ведущая цепь синтезируется в направлении от 5' к 3' концу (при этом
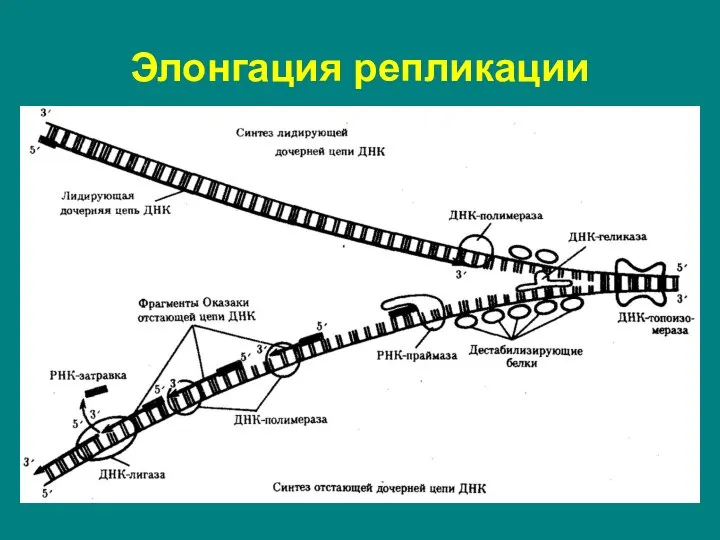
Слайд 12Происходит при достижении репликационной вилкой теломер (концевой участок хоромосом).
Теломеры состоят из повторяющихся
Происходит при достижении репликационной вилкой теломер (концевой участок хоромосом).
Теломеры состоят из повторяющихся

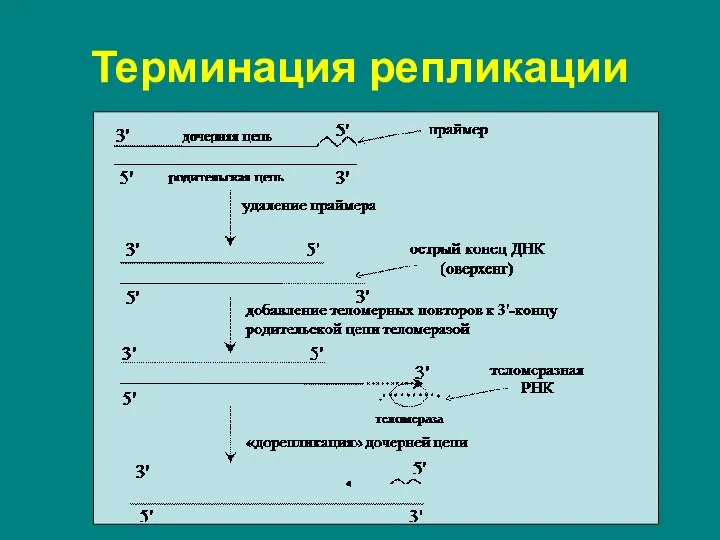
Слайд 13Терминация репликации
Теломераза участвует в репликации теломерных (концевых) участков линейной ДНК.
ДНК-полимеразная система, описанная
Терминация репликации
Теломераза участвует в репликации теломерных (концевых) участков линейной ДНК.
ДНК-полимеразная система, описанная

Слайд 14Терминация репликации
Терминация репликации
Слайд 15Репарация
устранение из ДНК возникающих в ней повреждений, восстановление исходной (нативной) структуры молекулы
Репарация
устранение из ДНК возникающих в ней повреждений, восстановление исходной (нативной) структуры молекулы

Слайд 16Репарационные системы:
простые (одноэтапные) – фотореактивация, деалкилирование;
сложные (многоэтапные, многокомпонентные).
Основная цель этих систем –
Репарационные системы:
простые (одноэтапные) – фотореактивация, деалкилирование;
сложные (многоэтапные, многокомпонентные).
Основная цель этих систем –

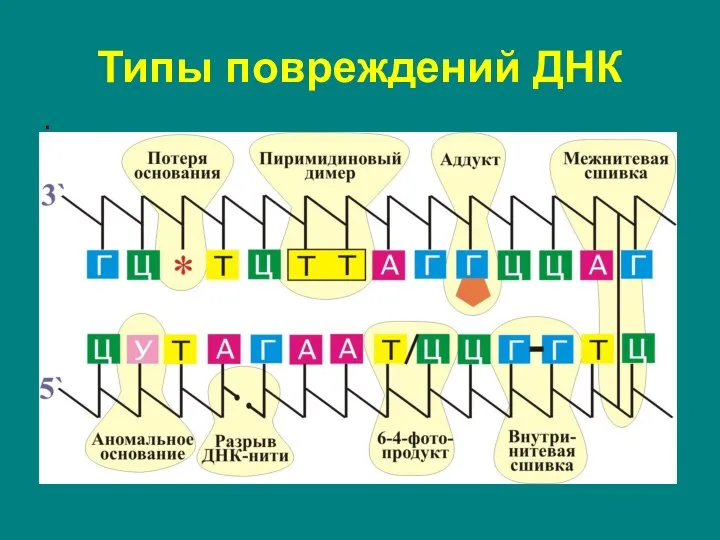
Слайд 17Типы повреждений ДНК
1. Спонтанные повреждения:
ошибки репликации (появление некомплементарных пар нуклеотидов);
апуринизация (отщепление азотистых
Типы повреждений ДНК
1. Спонтанные повреждения:
ошибки репликации (появление некомплементарных пар нуклеотидов);
апуринизация (отщепление азотистых

Слайд 18Типы повреждений ДНК
2. Индуцированные повреждения:
димеризация (сшивание соседних пиримидиновых оснований с образованием димера);
разрывы
Типы повреждений ДНК
2. Индуцированные повреждения:
димеризация (сшивание соседних пиримидиновых оснований с образованием димера);
разрывы

Слайд 19.
Типы повреждений ДНК
.
Типы повреждений ДНК
Слайд 20С позиций молекулярного механизма первичные повреждения в молекулах ДНК могут быть устранены
С позиций молекулярного механизма первичные повреждения в молекулах ДНК могут быть устранены

Слайд 21По отношению к процессу репликации различают два основные типа репарации ДНК:
дорепликативную (фотореактивация
По отношению к процессу репликации различают два основные типа репарации ДНК:
дорепликативную (фотореактивация

Слайд 22Этот тип репарации обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения.
Этот тип репарации обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения.

Слайд 23Репарация ДНК (фотореактивация)
1. Нормальная молекула ДНК
↓
Облучение УФ-светом
↓
2. Мутантная молекула ДНК – образование
Репарация ДНК (фотореактивация)
1. Нормальная молекула ДНК
↓
Облучение УФ-светом
↓
2. Мутантная молекула ДНК – образование

Слайд 24Репарация ДНК (фотореактивация)
Репарация ДНК (фотореактивация)

Слайд 25Репарация ДНК при алкилирующих повреждениях
Генетические повреждения, вызываемые присоединением алкильных или метильных групп,
Репарация ДНК при алкилирующих повреждениях
Генетические повреждения, вызываемые присоединением алкильных или метильных групп,

Слайд 26Репарация ДНК – прямая вставка пуринов
При некоторых типах повреждений пуриновых оснований ковалентная
Репарация ДНК – прямая вставка пуринов
При некоторых типах повреждений пуриновых оснований ковалентная

Слайд 27Репарация ДНК при однонитевых разрывах
Например, под действием ионизирующего облучения могут возникнуть однонитевые
Репарация ДНК при однонитевых разрывах
Например, под действием ионизирующего облучения могут возникнуть однонитевые

Слайд 28Репарация ДНК (эксцизионная)
Этот сложные реакции восстановления, когда поврежденные участки вырезаются из цепи
Репарация ДНК (эксцизионная)
Этот сложные реакции восстановления, когда поврежденные участки вырезаются из цепи

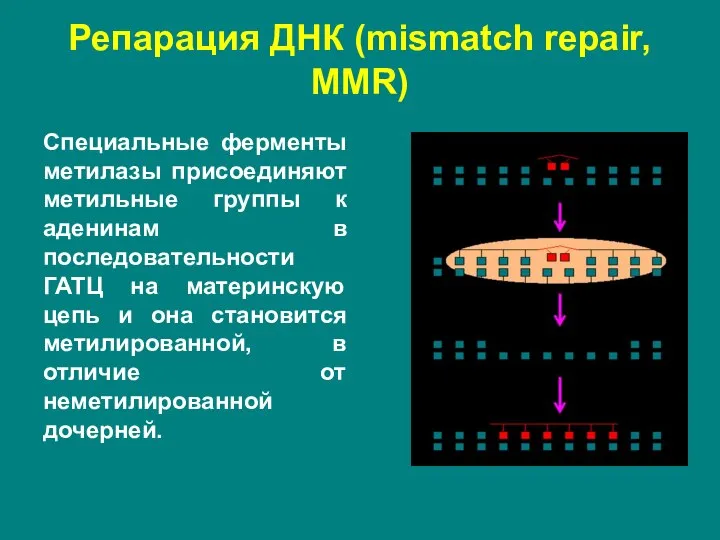
Слайд 29Репарация ДНК (mismatch repair, MMR)
Во время репликации ДНК происходят ошибки спаривания, в
Репарация ДНК (mismatch repair, MMR)
Во время репликации ДНК происходят ошибки спаривания, в

Слайд 30Специальные ферменты метилазы присоединяют метильные группы к аденинам в последовательности ГАТЦ на
Специальные ферменты метилазы присоединяют метильные группы к аденинам в последовательности ГАТЦ на
Слайд 31Пострепликативная репарация
Осуществляется в тех случаях, когда повреждение доживает до фазы репликации или
Пострепликативная репарация
Осуществляется в тех случаях, когда повреждение доживает до фазы репликации или

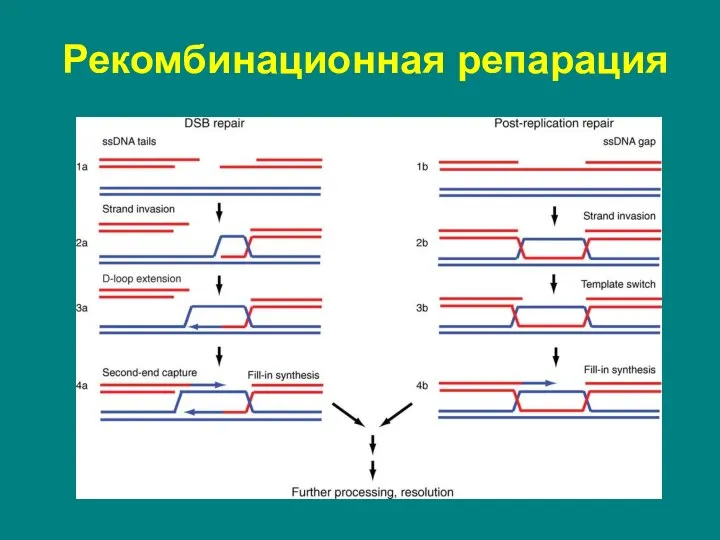
Слайд 32Рекомбинационная репарация
Рекомбинационная репарация
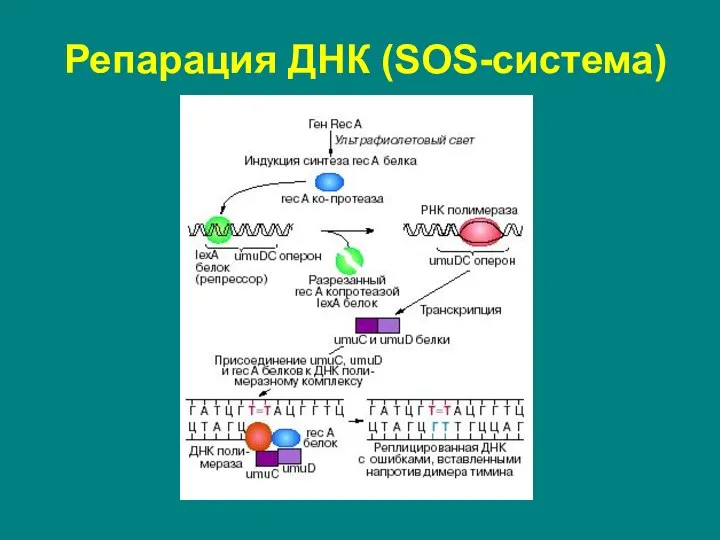
Слайд 33Репарация ДНК (SOS-система)
Обнаружена в 1974 г. М.Радманом.
Включается тогда, когда повреждений в
Репарация ДНК (SOS-система)
Обнаружена в 1974 г. М.Радманом.
Включается тогда, когда повреждений в

Слайд 34Репарация ДНК (SOS-система)
Репарация ДНК (SOS-система)
Слайд 35Репарация ДНК и наследственные болезни человека
Нарушение системы репарации у человека является причиной:
преждевременного
Репарация ДНК и наследственные болезни человека
Нарушение системы репарации у человека является причиной:
преждевременного

Слайд 36Прогерия детей – синдром Хатчинсона-Гилфорда
маленький рост
птичье лицо с клювообразным профилем
преобладание мозгового черепа
Прогерия детей – синдром Хатчинсона-Гилфорда
маленький рост
птичье лицо с клювообразным профилем
преобладание мозгового черепа

Слайд 37Прогерия взрослых- синдром Вернера
Симптомы старения проявляются после полового созревания:
Седина, выпадение волос, морщины,
Прогерия взрослых- синдром Вернера
Симптомы старения проявляются после полового созревания:
Седина, выпадение волос, морщины,

 Сохраним мир птиц
Сохраним мир птиц Альбом по эмбриологии
Альбом по эмбриологии Helicobacter Pylori (НР)
Helicobacter Pylori (НР) Методы регистрации клеточного цикла
Методы регистрации клеточного цикла Внешнее строение птиц
Внешнее строение птиц Me gusta la naturaleza
Me gusta la naturaleza Вирусы. Бактерии. Археи
Вирусы. Бактерии. Археи Внешнее строение птиц. Опорно-двигательная система птиц
Внешнее строение птиц. Опорно-двигательная система птиц Подготовка к ЕГЭ по биологии. Решение заданий из части С
Подготовка к ЕГЭ по биологии. Решение заданий из части С Общебиологическая характеристика организма человека
Общебиологическая характеристика организма человека Основные понятия генетики
Основные понятия генетики Корсак. Внешний вид
Корсак. Внешний вид Папоротники и папоротниковидные растения
Папоротники и папоротниковидные растения Салауатты өмір салтын тәрбиелеу жүйесіндегі дене
Салауатты өмір салтын тәрбиелеу жүйесіндегі дене Презентация на тему Влияние алкаголя на организм человека
Презентация на тему Влияние алкаголя на организм человека  Вегетативное размножение древесных растений
Вегетативное размножение древесных растений Презентация на тему Экологическое многообразие птиц
Презентация на тему Экологическое многообразие птиц  2
2 Во что одеты растения
Во что одеты растения Грудь и брюшко насекомых
Грудь и брюшко насекомых Пифарнин (ribavirin)
Пифарнин (ribavirin) Растения Томской области занесенные в Красную книгу
Растения Томской области занесенные в Красную книгу Презентация на тему Биологические катализаторы
Презентация на тему Биологические катализаторы  Нарушения теплового баланса
Нарушения теплового баланса Общая характеристика клеточного уровня. 9 класс
Общая характеристика клеточного уровня. 9 класс Такие удивительные пауки
Такие удивительные пауки Гровинг (Growing)
Гровинг (Growing) Сафари-парк Тайган
Сафари-парк Тайган