- МБ ЛК3, Тема3(2), Транскрипция

Содержание
- 2. Тема 3(2). ФУНКЦИИ ДНК ТРАНСКРИПЦИЯ
- 3. Т Р А Н С К Р И П Ц И Я (прокариоты)
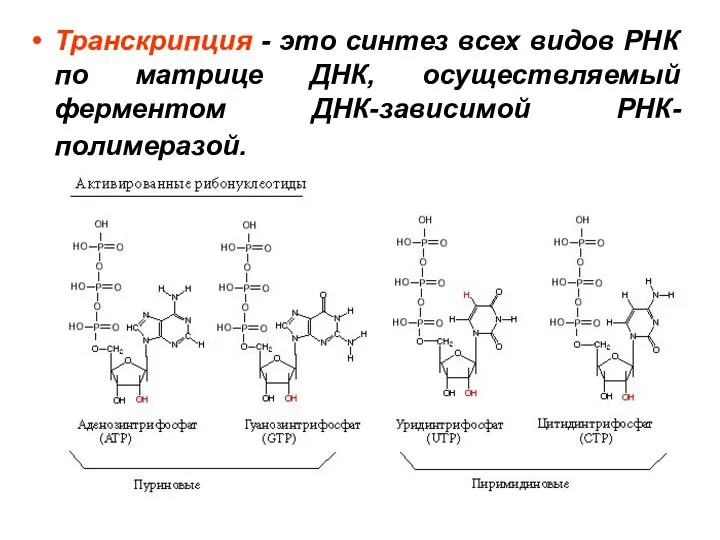
- 4. Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой РНК-полимеразой.
- 5. Принципы транскрипции: 1. Комплементарность. 2. Антипараллельность. 3. Униполярность. 4. Беззатравочность. 5. Асимметричность. РНК синтезируется комплементарно и
- 6. Понятие об опероне Оперон - единица транскрипции у прокариот
- 7. Промотор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как посадочная площадка и старт синтеза РНК. Только
- 8. Оператор - особая последовательность нуклеотидов ДНК, узнаваемая белком-репрессором.
- 9. Асимметричность Транскрибируются обе цепи ДНК, но в каждом отдельном опероне только одна из них. Какая именно,
- 10. Особенности структуры промотора
- 11. Узнавание и прочное связывание происходит на разных участках ДНК. Эти участки отличаются и по первичной, и
- 12. Структура ДНК-зависимой РНК-полимеразы E.coli 6 субъединиц: 2α β β’ ω δ - холофермент 2α β β’
- 13. α2: две α-субъединицы связывают остальные элементы фермента и распознают регулирующие факторы. Каждая субъединица состоит из двух
- 21. Этапы транскрипции 1. Узнавание и прочное связывание
- 22. Примерно 5% промоторов у прокариот имеют только участок "-10", однако, тем не менее, хорошо узнаются РНК-полимеразой.
- 23. 2. Инициация заключается в образовании первой фосфодиэфирной связи между пурин-трифосфатом (АТФ или ГТФ) и следующим нуклеотидом.
- 24. СХЕМА ЭТАПОВ ТРАНСКРИПЦИИ
- 26. - независимая - зависимая терминация терминация
- 27. - независимая терминация
- 28. Регуляция транскрипции у прокариот Схема негативной индукции Жакоба и Моно Lac-оперон E. coli содержит 3 гена,
- 29. Эта схема называется так потому, что контролирующим транскрипцию фактором является негативный фактор, "выключатель" - белок -
- 30. Схема позитивной индукции Аra-оперон E. сoli Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент -
- 31. Схема позитивной репрессии Оперон синтеза рибофлавина у Вacilus subtilis. Позитивная репрессия, поскольку в регуляции участвует белок
- 32. Схема негативной репрессии Оперон синтеза триптофана у E. сoli. Схема регуляции - негативная репрессия, потому что
- 33. Т Р А Н С К Р И П Ц И Я (эукариоты) ЛЕКЦИЯ 8
- 34. У эукариотов процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция
- 35. Особенности транскрипции эукариот Единицей транскрипции у эукариот является отдельный ген, а не оперон, как у прокариот.
- 36. Для любого гена, кодирующего белок, есть энхансеры (усилители). Энхансеры - последовательности ДНК, усиливающие транскрипцию при взаимодействии
- 37. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют
- 38. Этапы транскрипции Инициация – выбор фиксированного места начала транскрипции (специфическое связывание РНК-пол с матричной нитью), промотор
- 39. Три этапа транскрипции Инициация Формирование транскрипционного пузырька 12-14 п.н. Синтез РНК в направлении от 5’-конца к
- 40. Три этапа транскрипции Элонгация Нематричная нить – кодирующая (кодогенная) Матричная нить –транскрибируемая РНК-пол движется по нити
- 41. РНК-полимераза II для осуществления своей работы требует много различных белковых факторов (транскрипционные факторы – ТФ). РНК-полимераза
- 42. Комплекс ДНК и РНК-полимеразы II
- 43. Белки, необходимые для инициации транскрипции на эукариотических промоторах РНК-полимеразой II
- 45. ИНГИБИТОРЫ РНК-ПОЛИМЕРАЗ Актиномицин D и акридин – подавляют работу на стадии элонгации α- аманитин (токсин бледной
- 46. Процессинг мРНК Процессинг мРНК состоит из нескольких этапов. 1. Кепирование 100% мРНК 2. Полиаденилирование ~95% мРНК
- 47. Процессинг мРНК
- 48. Кепирование Кепирование - надевание "шапочки". "Сар" представляет собой метилированный GTP, присоединенный в необычной позиции 5'-5' и
- 49. Назначение “Кэп" 1. Защита 5'-конца мРНК от действия экзонуклеаз. 2. За счет узнавания “Кэп" связывающими белками
- 50. Полиаденилирование Когда синтез пре-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3' -
- 52. мРНК ряда генов не полиаденилируется (например гистоновых генов). Полиаденилированные пре-мРНК подвергаются сплайсингу.
- 53. Сплайсинг Экзоны - кодирующие участки генов. Интроны - некодирующие участки генов. Сплайсинг - вырезание копий интронов
- 54. Для мРНК высших организмов существуют обязательные правила сплайсинга: Правило 1. 5' и 3' концы интрона очень
- 55. Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене, но могут быть выброшены
- 56. Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в которых помимо ферментов, вырезающих и сшивающих участки пре-мРНК, имеются
- 57. Редактирование Редактирование - изменение генетической информации на уровне мРНК. Пример- редактирование мРНК цитохромоксидазы у трипаносомы: когда
- 58. Процессинг РНК в клетках прокариот и эукариот ЛЕКЦИЯ 9
- 59. Этапы реализации генетической информации Транскрипция – синтез молекул РНК, образование первичного транскрипта (пре-РНК) Процессинг –модификация первичного
- 60. Процессинг РНК в клетках прокариот Ген – это участок молекулы ДНК, кодирующий синтез функциональной молекулы РНК
- 61. Процессинг рРНК и тРНК Зрелые молекулы рРНК и тРНК образуются у прокариот и эукариот в результате
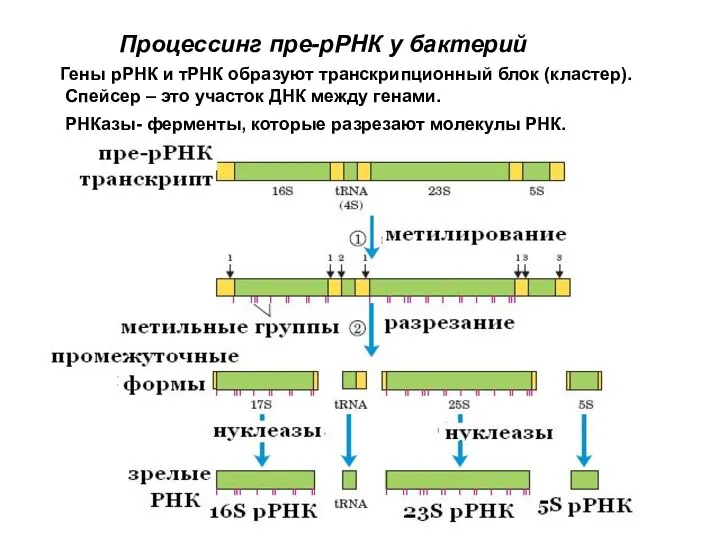
- 62. Гены рРНК и тРНК образуют транскрипционный блок (кластер). Спейсер – это участок ДНК между генами. РНКазы-
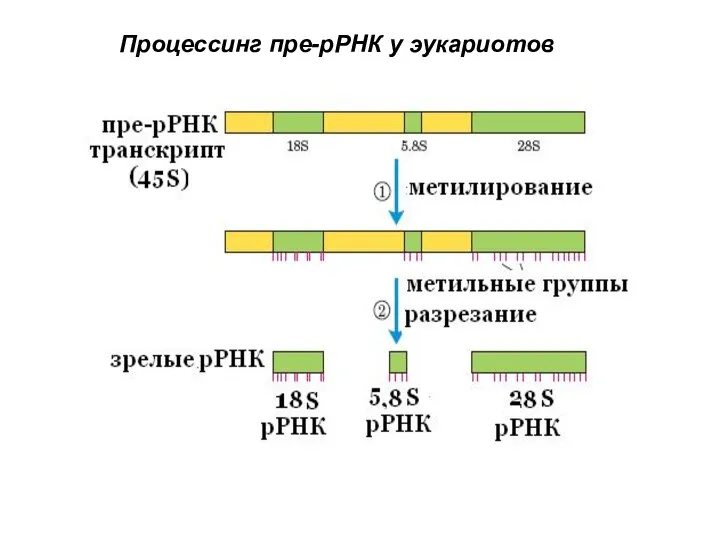
- 63. Процессинг рРНК эукариот Транскрипционный блок содержит гены 18S ; 5,8S; 28S - рРНК, разделенные спейсерами три
- 64. Процессинг пре-рРНК у эукариотов
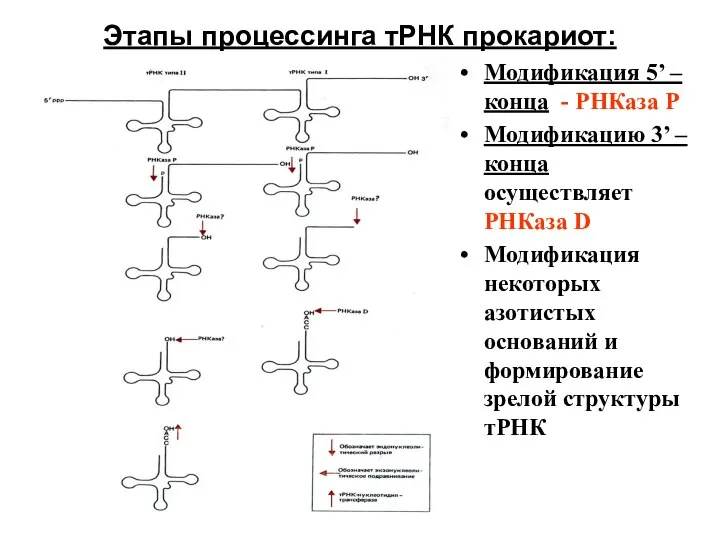
- 65. Этапы процессинга тРНК прокариот: Модификация 5’ – конца - РНКаза Р Модификацию 3’ – конца осуществляет
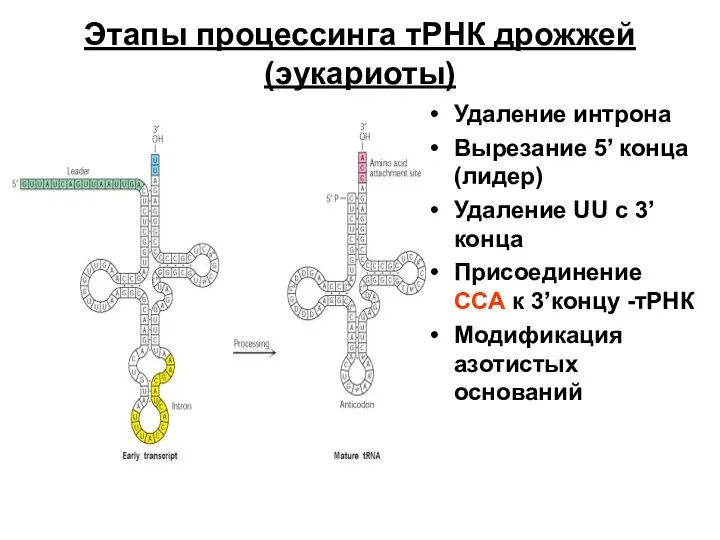
- 66. Этапы процессинга тРНК дрожжей (эукариоты) Удаление интрона Вырезание 5’ конца (лидер) Удаление UU с 3’конца Присоединение
- 67. Экзон-интронная структура генов эукариот Структура α- и β-глобиновых генов Экзоны (тёмно-красный цвет ), разделены интронами (голубой
- 68. Этапы процессинга пре - мРНК эукариот Кэпирование - модификация 5’-конца Полиаденилирование - модификация 3’-конца Сплайсинг -
- 70. Модификация 5’-конца – кэпирование Кэп – это 7-метил-гуанозин соединенный в 5’-5’-ориентации с первым нуклеотидом мРНК Кэп
- 71. Модификация 3’-конца пре-мРНК - полиаденилирование
- 72. Механизмы сплайсинга интронов: Тип I - интроны подвергаются аутосплайсингу в присутствии только ионов Mg +2 и
- 73. Сплайсинг интронов типа I
- 74. Сплайсинг ядерной мРНК происходит в сплайсосоме Сплайсосома - специальная ядерная структура, в которой происходит сплайсинг В
- 75. Взаимодействие компонентов сплайсосомы с экзонами и интронами РНК
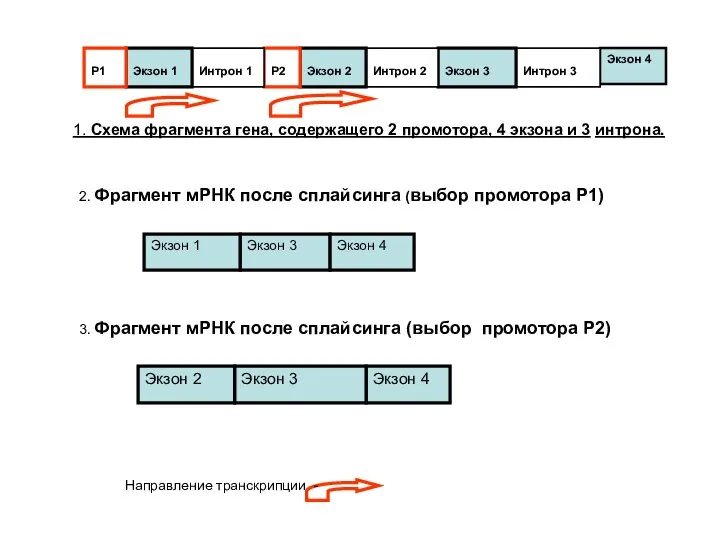
- 76. Механизмы альтернативного сплайсинга: Альтернативный выбор промотора Альтернативный выбор сигнала полиаденилирования Альтернативный выбор разных наборов экзонов Транс-сплайсинг
- 77. Экзон 4 Интрон 3 Экзон 3 Интрон 2 Экзон 2 Р2 Интрон 1 Экзон 1 Р1
- 78. Альтернативный сплайсинг мРНК кальцитонинового гена у млекопитающих (крыса)
- 79. Структура мРНК прокариот Лидер - это 5’ не транслируемый участок - 5’ UTR (UnTranslated Region) Трейлер
- 81. Скачать презентацию

Слайд 3
Т Р А Н С К Р И П Ц И Я
(прокариоты)
Т Р А Н С К Р И П Ц И Я
(прокариоты)

Слайд 4Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом
Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом
Слайд 5
Принципы транскрипции:
1. Комплементарность.
2. Антипараллельность.
3. Униполярность.
4. Беззатравочность.
Принципы транскрипции:
1. Комплементарность.
2. Антипараллельность.
3. Униполярность.
4. Беззатравочность.

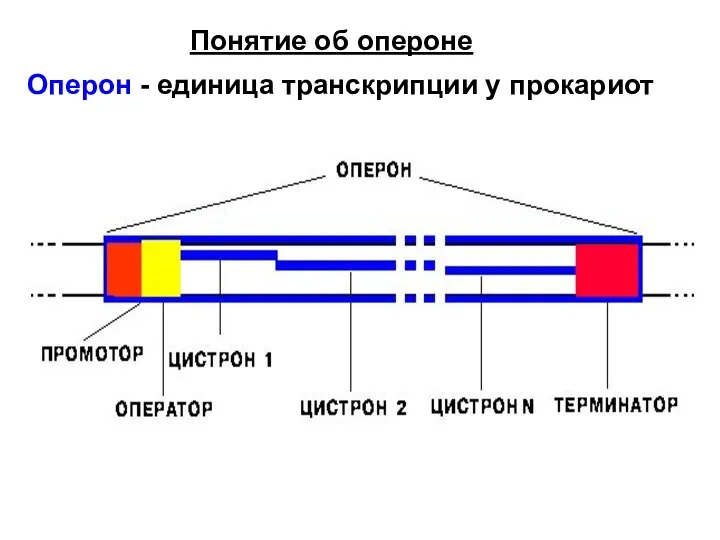
Слайд 6 Понятие об опероне
Оперон - единица транскрипции у прокариот
Понятие об опероне
Оперон - единица транскрипции у прокариот
Слайд 7
Промотор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как посадочная площадка
Промотор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как посадочная площадка

Слайд 8
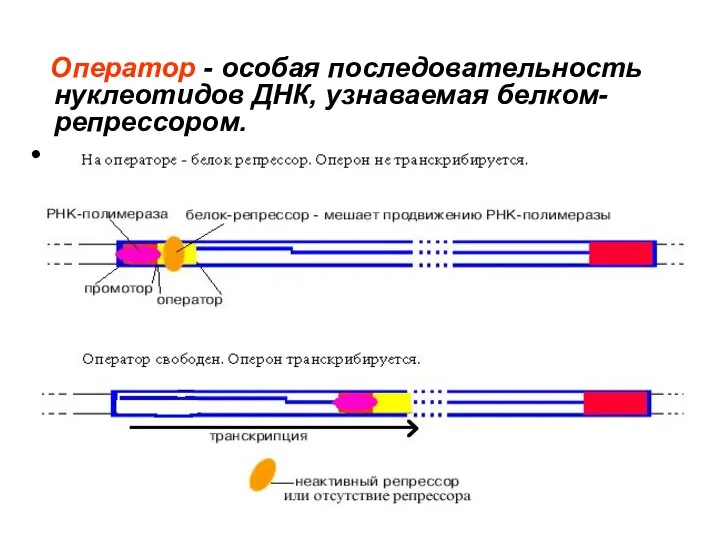
Оператор - особая последовательность нуклеотидов ДНК, узнаваемая белком-репрессором.
Оператор - особая последовательность нуклеотидов ДНК, узнаваемая белком-репрессором.
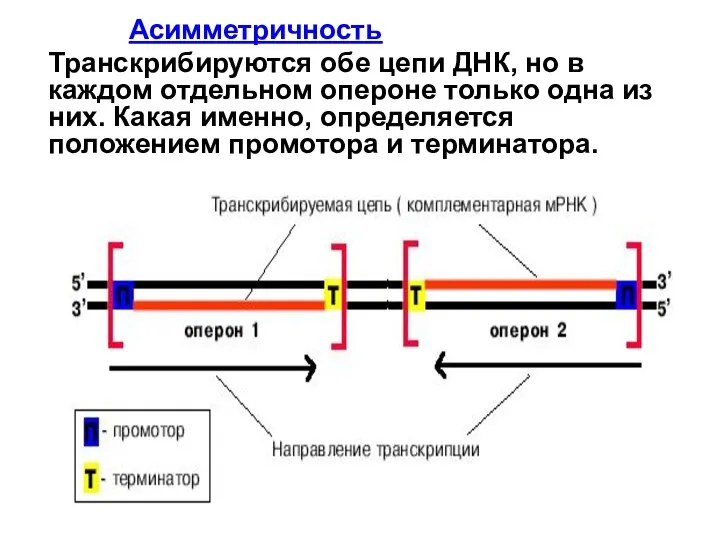
Слайд 9 Асимметричность
Транскрибируются обе цепи ДНК, но в каждом отдельном опероне только одна
Асимметричность
Транскрибируются обе цепи ДНК, но в каждом отдельном опероне только одна
Слайд 10
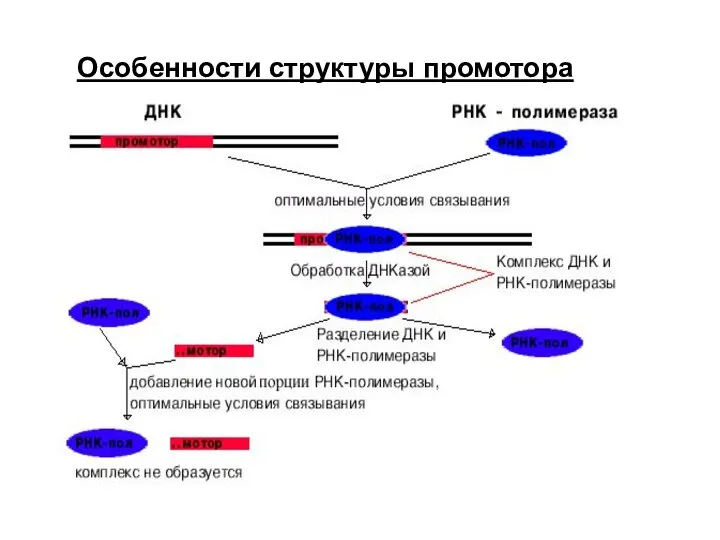
Особенности структуры промотора
Особенности структуры промотора
Слайд 11
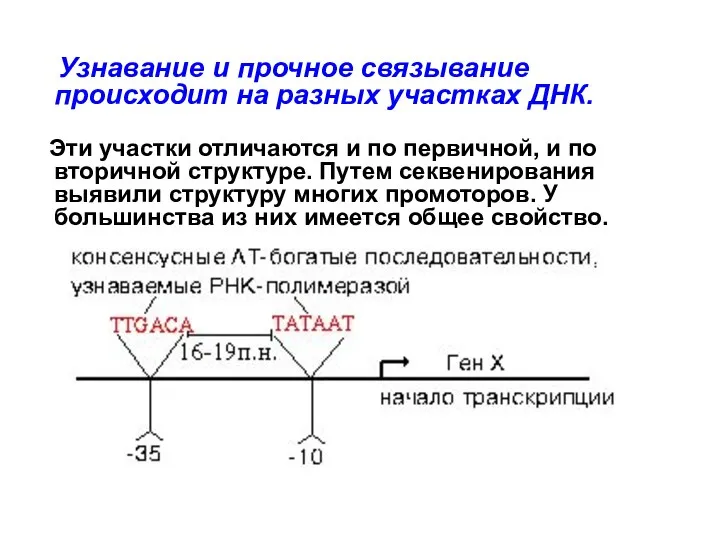
Узнавание и прочное связывание происходит на разных участках ДНК.
Эти
Узнавание и прочное связывание происходит на разных участках ДНК.
Эти
Слайд 12
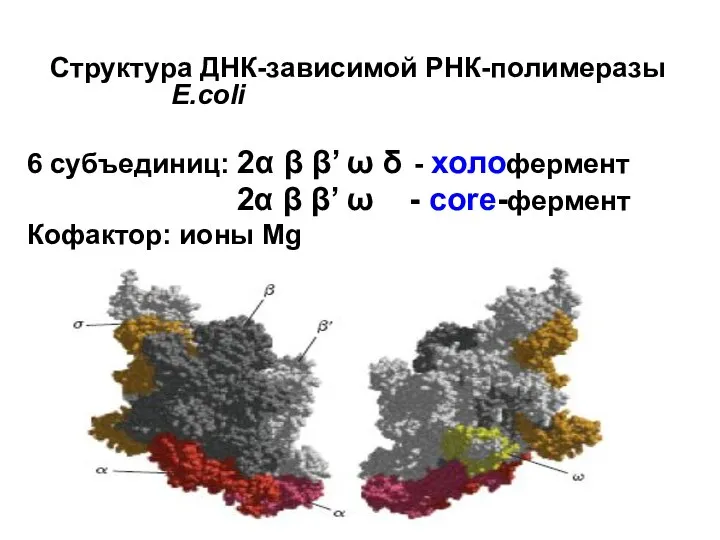
Структура ДНК-зависимой РНК-полимеразы E.coli
6 субъединиц: 2α β β’ ω
Структура ДНК-зависимой РНК-полимеразы E.coli
6 субъединиц: 2α β β’ ω
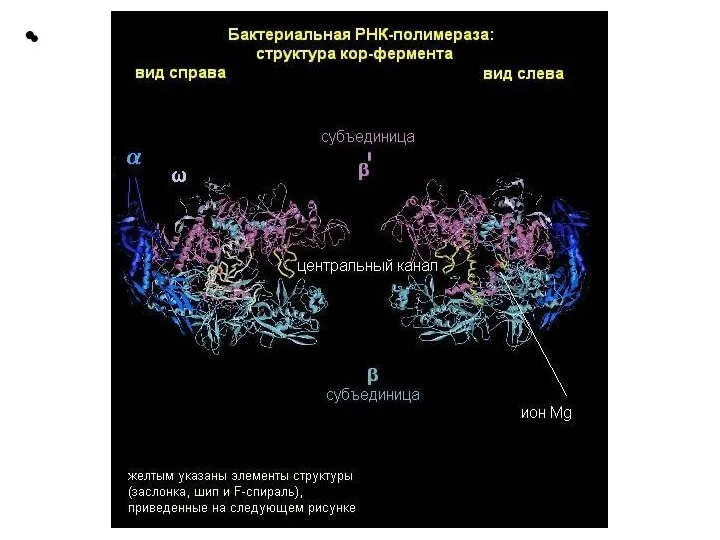
Слайд 13α2: две α-субъединицы связывают остальные элементы фермента и распознают регулирующие факторы. Каждая
α2: две α-субъединицы связывают остальные элементы фермента и распознают регулирующие факторы. Каждая

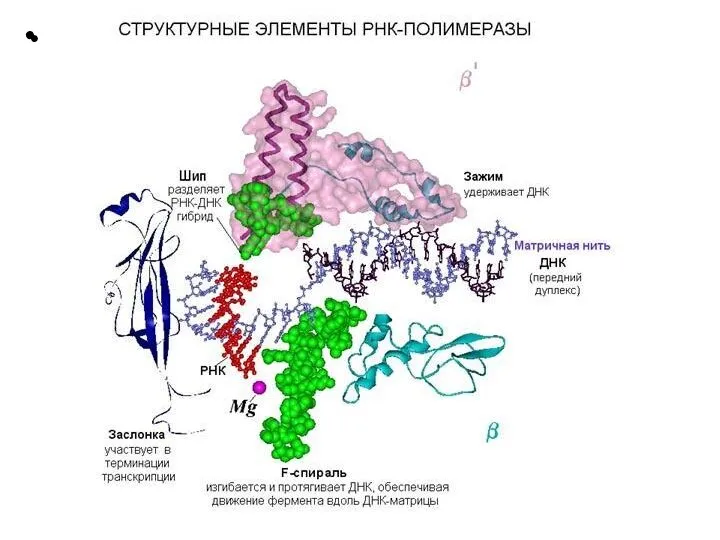
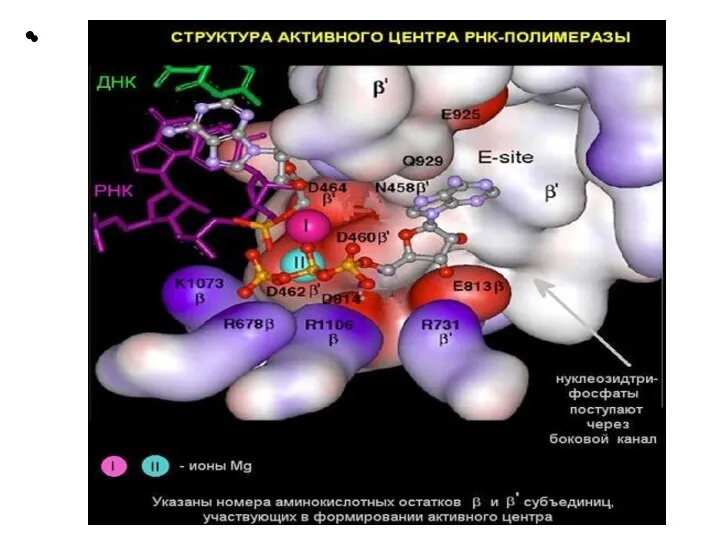
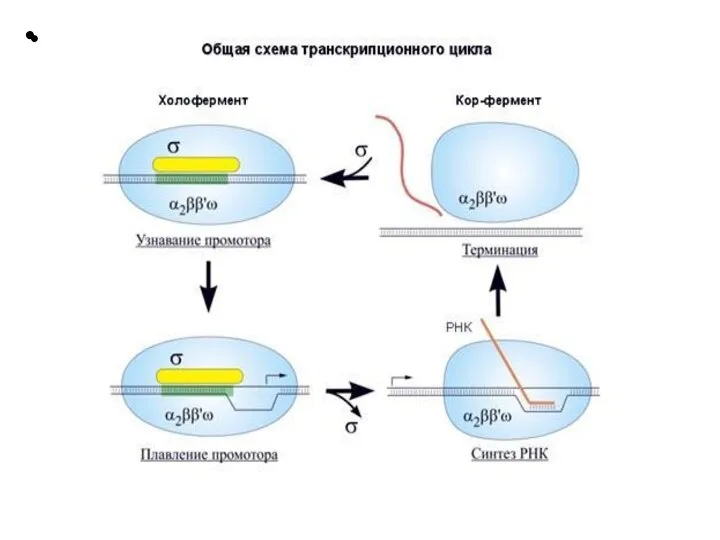
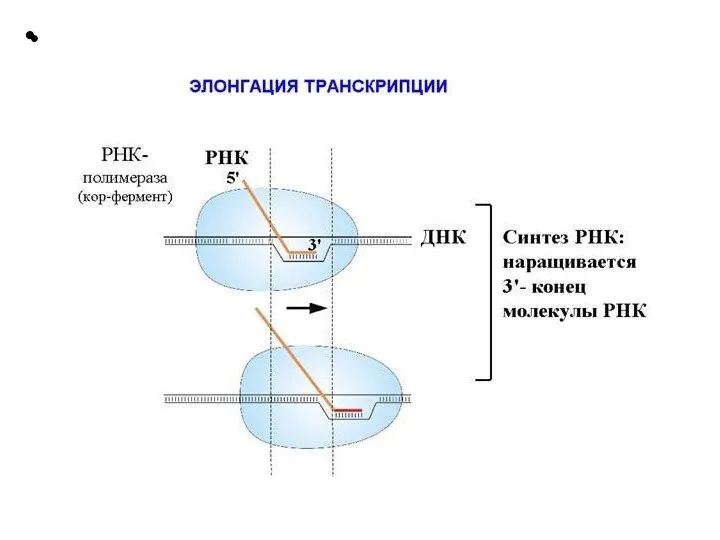
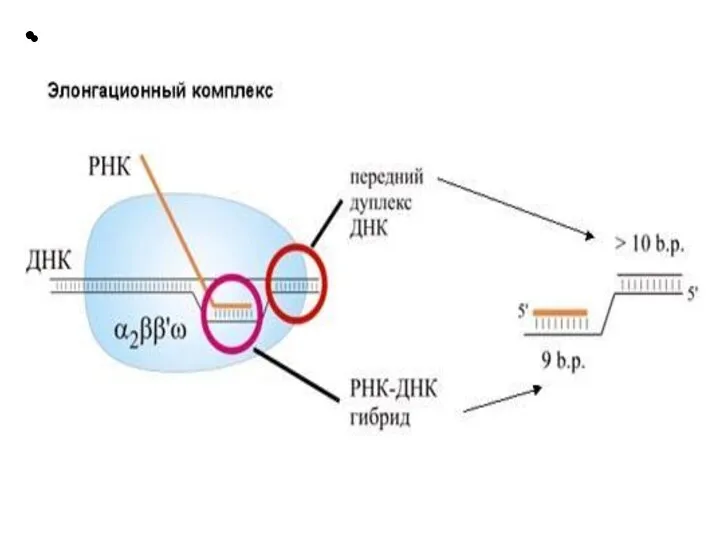
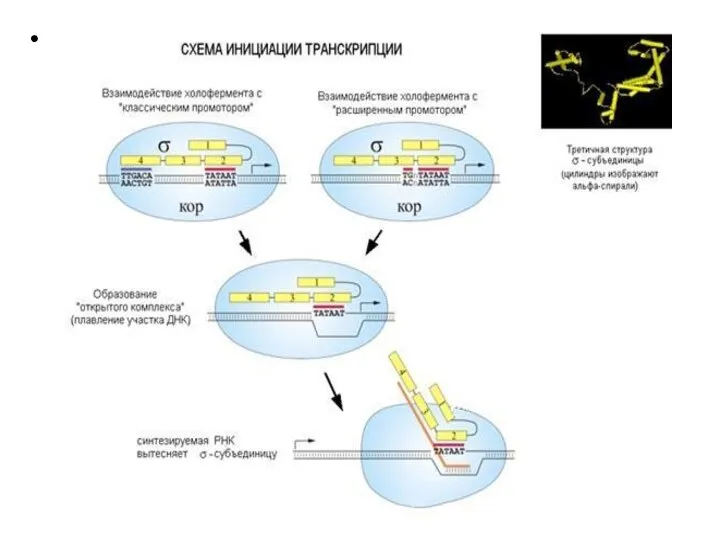
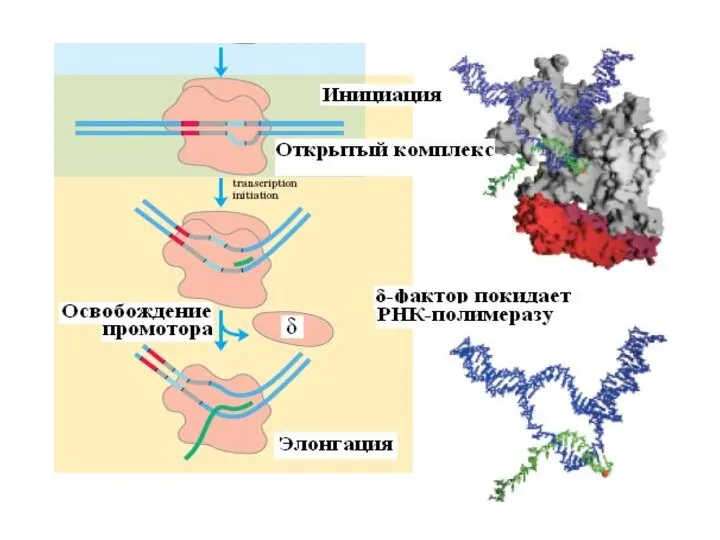
Слайд 21 Этапы транскрипции
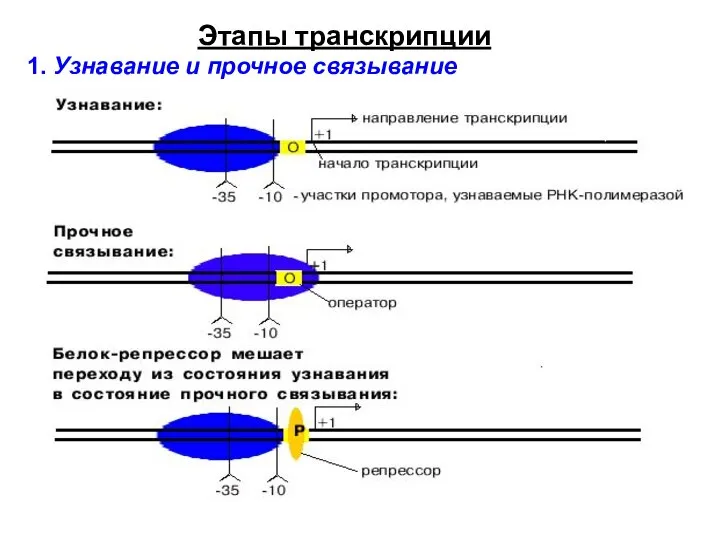
1. Узнавание и прочное связывание
Этапы транскрипции
1. Узнавание и прочное связывание
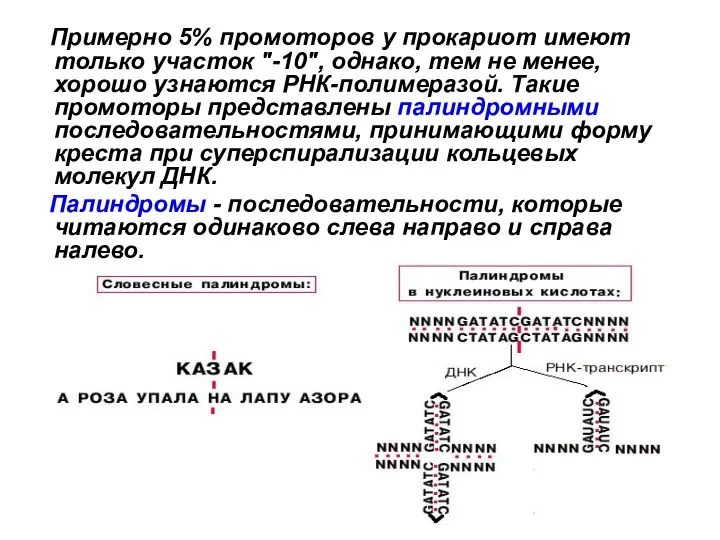
Слайд 22 Примерно 5% промоторов у прокариот имеют только участок "-10", однако, тем
Примерно 5% промоторов у прокариот имеют только участок "-10", однако, тем
Слайд 23 2. Инициация заключается в образовании первой фосфодиэфирной связи между пурин-трифосфатом (АТФ
2. Инициация заключается в образовании первой фосфодиэфирной связи между пурин-трифосфатом (АТФ

Слайд 24
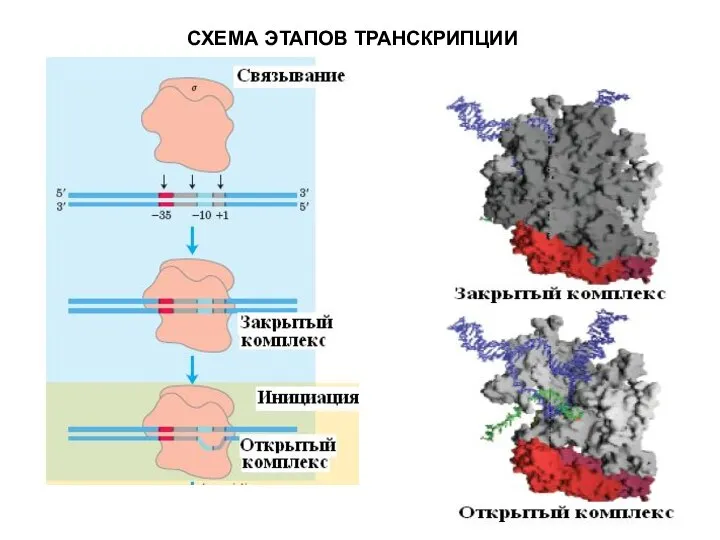
СХЕМА ЭТАПОВ ТРАНСКРИПЦИИ
СХЕМА ЭТАПОВ ТРАНСКРИПЦИИ
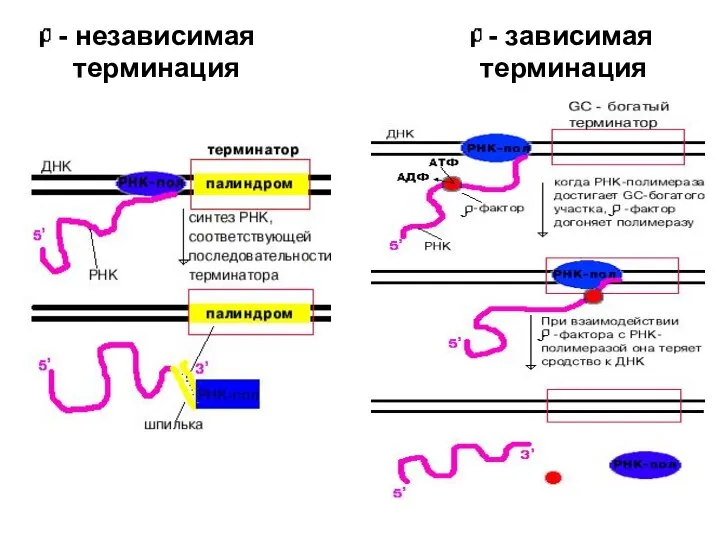
Слайд 26 - независимая - зависимая
терминация терминация
- независимая - зависимая
терминация терминация
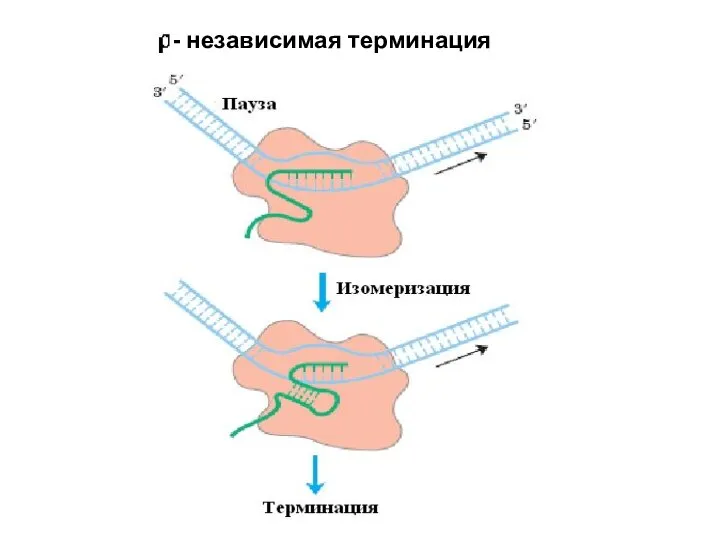
Слайд 27
- независимая терминация
- независимая терминация
Слайд 28
Регуляция транскрипции у прокариот
Схема негативной индукции Жакоба и Моно
Lac-оперон E.
Регуляция транскрипции у прокариот
Схема негативной индукции Жакоба и Моно
Lac-оперон E.

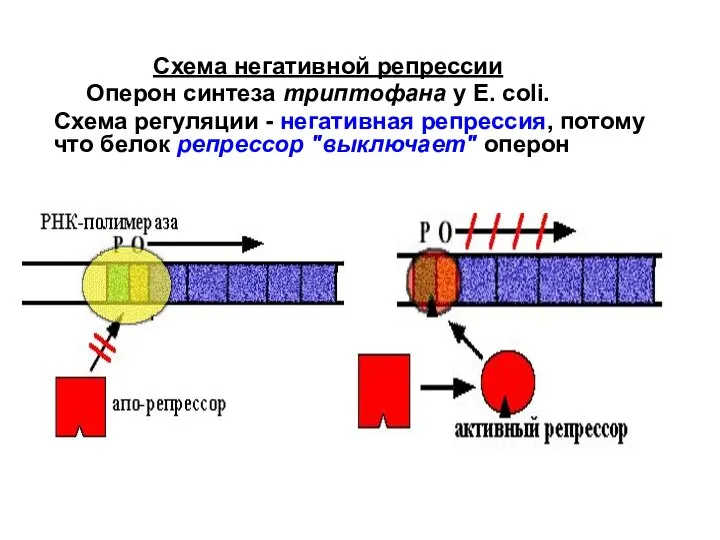
Слайд 29 Эта схема называется так потому, что контролирующим транскрипцию фактором является негативный
Эта схема называется так потому, что контролирующим транскрипцию фактором является негативный
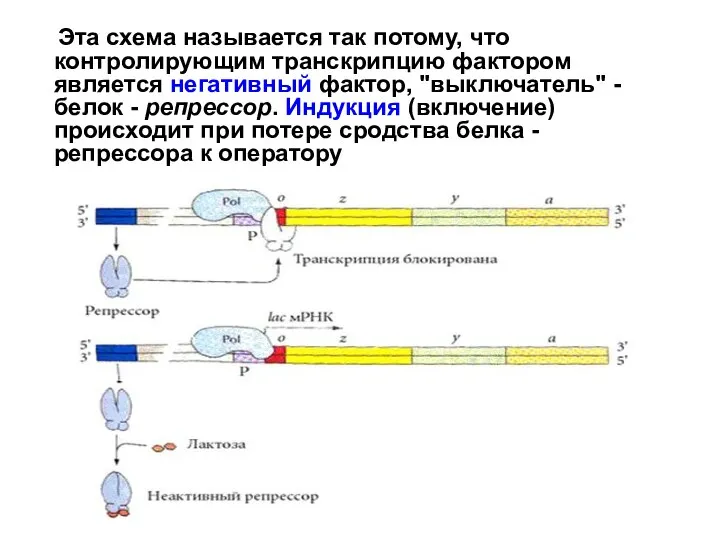
Слайд 30 Схема позитивной индукции Аra-оперон E. сoli
Эта схема регуляции называется позитивной
Схема позитивной индукции Аra-оперон E. сoli
Эта схема регуляции называется позитивной
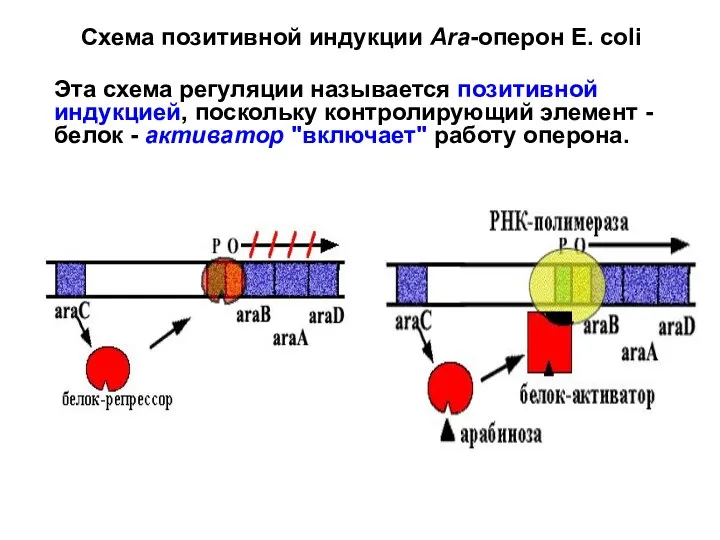
Слайд 31
Схема позитивной репрессии
Оперон синтеза рибофлавина у Вacilus subtilis.
Позитивная
Схема позитивной репрессии
Оперон синтеза рибофлавина у Вacilus subtilis.
Позитивная
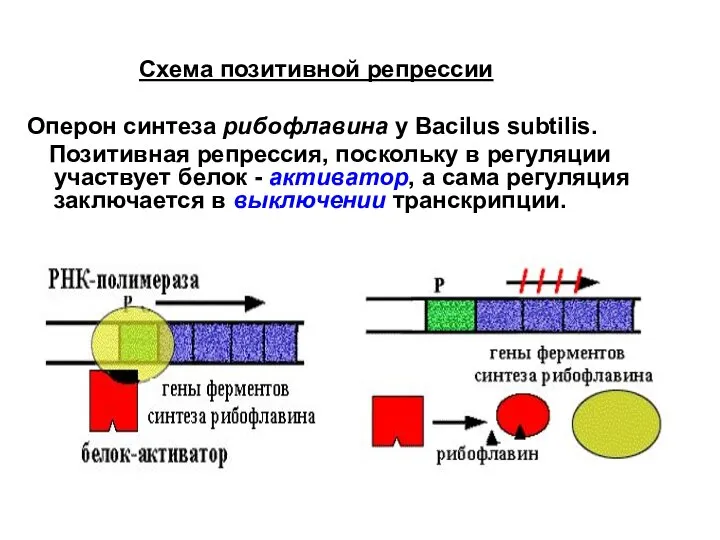
Слайд 32
Схема негативной репрессии
Оперон синтеза триптофана у E. сoli.
Схема
Схема негативной репрессии
Оперон синтеза триптофана у E. сoli.
Схема
Слайд 33
Т Р А Н С К Р И П Ц И Я
(эукариоты)
ЛЕКЦИЯ
Т Р А Н С К Р И П Ц И Я
(эукариоты)
ЛЕКЦИЯ

Слайд 34У эукариотов процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция
У эукариотов процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция

Слайд 35Особенности транскрипции эукариот
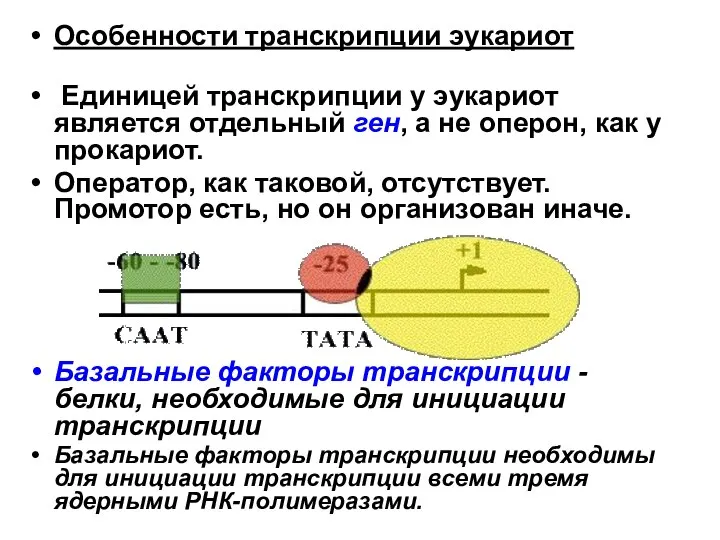
Единицей транскрипции у эукариот является отдельный ген, а не оперон,
Особенности транскрипции эукариот
Единицей транскрипции у эукариот является отдельный ген, а не оперон,
Слайд 36
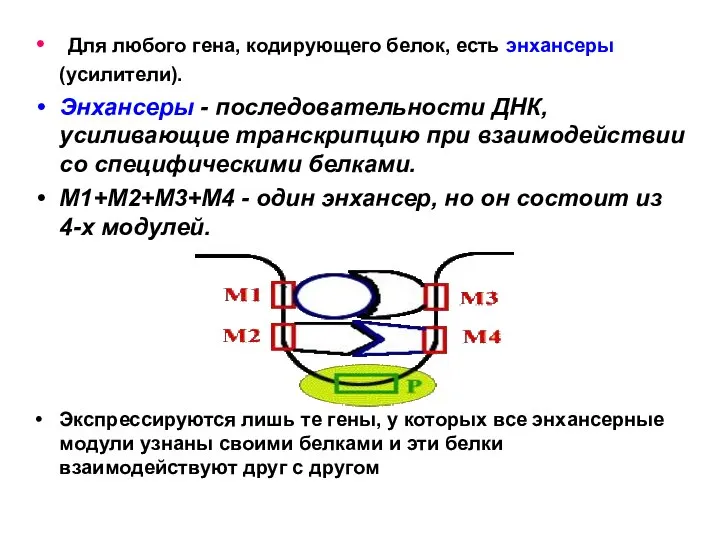
Для любого гена, кодирующего белок, есть энхансеры (усилители).
Энхансеры - последовательности ДНК,
Для любого гена, кодирующего белок, есть энхансеры (усилители).
Энхансеры - последовательности ДНК,
Слайд 37Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками
Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками

Слайд 38Этапы транскрипции
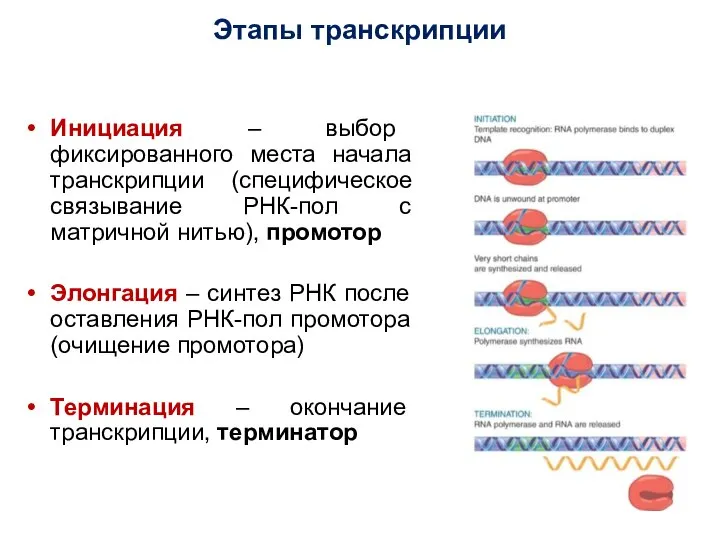
Инициация – выбор фиксированного места начала транскрипции (специфическое связывание РНК-пол с
Этапы транскрипции
Инициация – выбор фиксированного места начала транскрипции (специфическое связывание РНК-пол с
Слайд 39Три этапа транскрипции
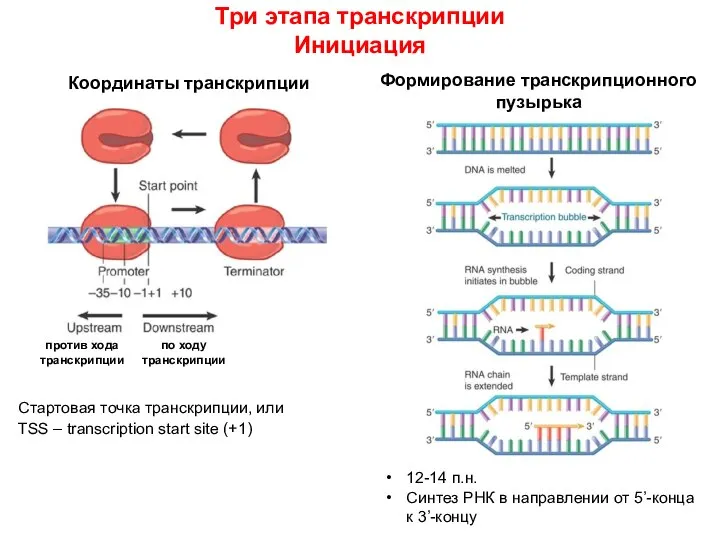
Инициация
Формирование транскрипционного пузырька
12-14 п.н.
Синтез РНК в направлении от 5’-конца к
Три этапа транскрипции
Инициация
Формирование транскрипционного пузырька
12-14 п.н.
Синтез РНК в направлении от 5’-конца к
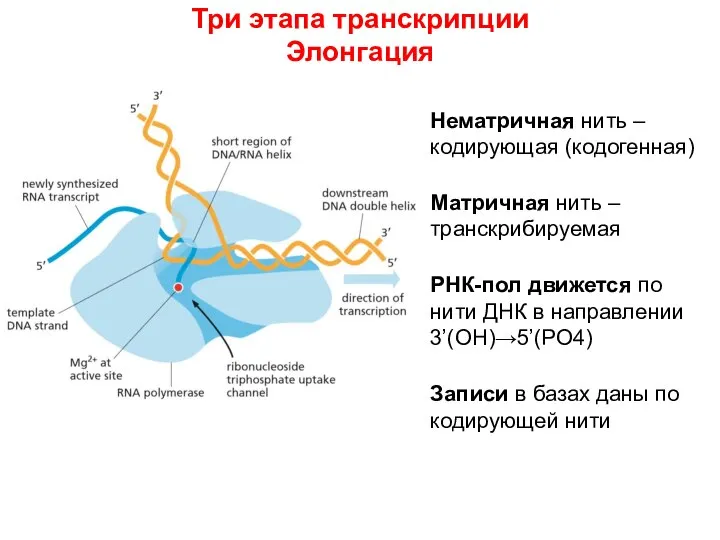
Слайд 40Три этапа транскрипции
Элонгация
Нематричная нить – кодирующая (кодогенная)
Матричная нить –транскрибируемая
РНК-пол движется по нити
Три этапа транскрипции
Элонгация
Нематричная нить – кодирующая (кодогенная)
Матричная нить –транскрибируемая
РНК-пол движется по нити
Слайд 41РНК-полимераза II для осуществления своей работы требует много различных белковых факторов (транскрипционные
РНК-полимераза II для осуществления своей работы требует много различных белковых факторов (транскрипционные

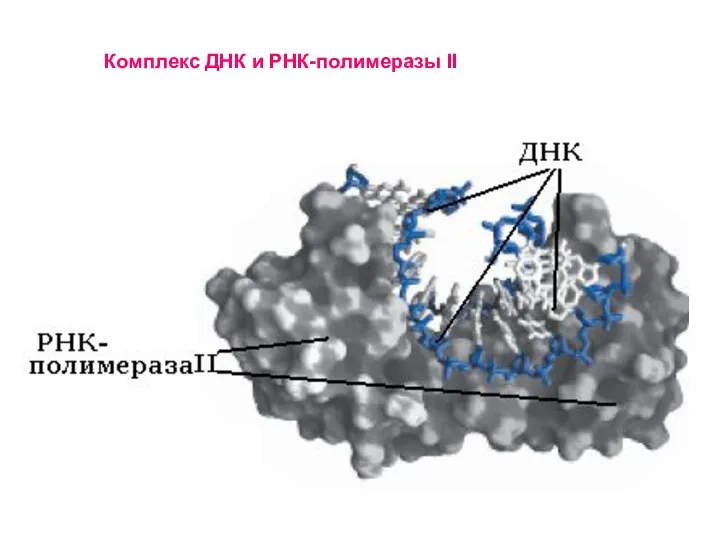
Слайд 42
Комплекс ДНК и РНК-полимеразы II
Комплекс ДНК и РНК-полимеразы II
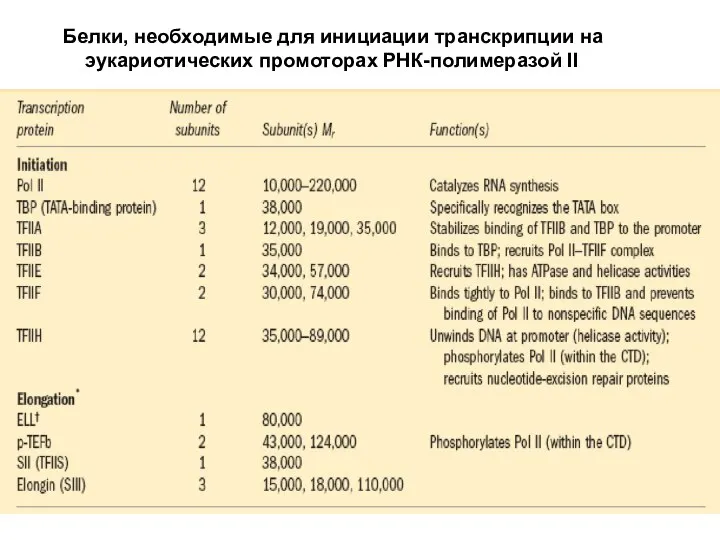
Слайд 43Белки, необходимые для инициации транскрипции на эукариотических промоторах РНК-полимеразой II
Белки, необходимые для инициации транскрипции на эукариотических промоторах РНК-полимеразой II
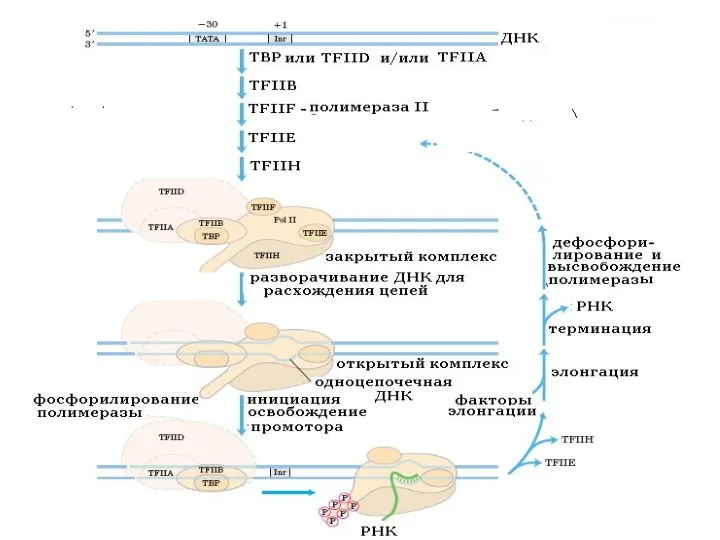
Слайд 45
ИНГИБИТОРЫ РНК-ПОЛИМЕРАЗ
Актиномицин D и акридин – подавляют работу на стадии элонгации
α-
ИНГИБИТОРЫ РНК-ПОЛИМЕРАЗ
Актиномицин D и акридин – подавляют работу на стадии элонгации
α-

Слайд 46Процессинг мРНК
Процессинг мРНК состоит из нескольких этапов.
1. Кепирование 100% мРНК
2. Полиаденилирование ~95%
Процессинг мРНК
Процессинг мРНК состоит из нескольких этапов.
1. Кепирование 100% мРНК
2. Полиаденилирование ~95%

Слайд 47 Процессинг мРНК
Процессинг мРНК
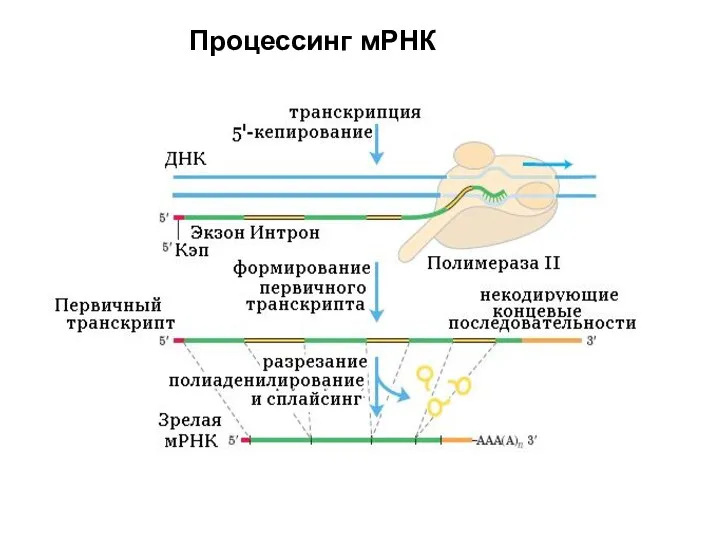
Слайд 48
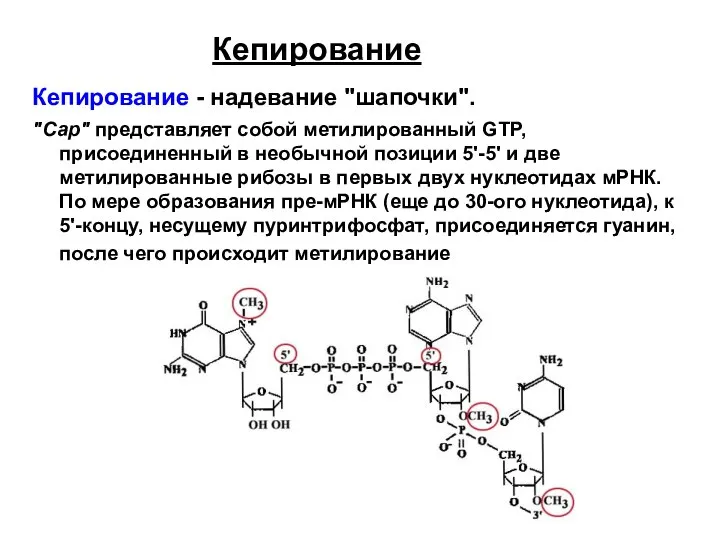
Кепирование
Кепирование - надевание "шапочки".
"Сар" представляет собой метилированный GTP, присоединенный в
Кепирование
Кепирование - надевание "шапочки".
"Сар" представляет собой метилированный GTP, присоединенный в
Слайд 49
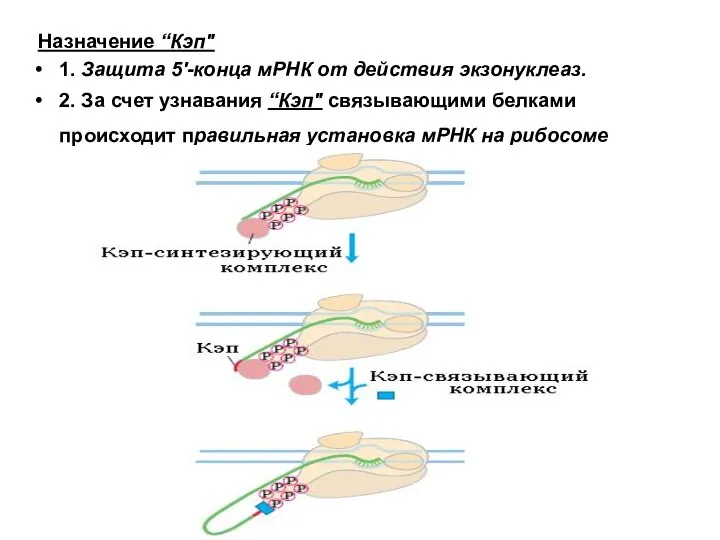
Назначение “Кэп"
1. Защита 5'-конца мРНК от действия экзонуклеаз.
2. За счет
Назначение “Кэп"
1. Защита 5'-конца мРНК от действия экзонуклеаз.
2. За счет
Слайд 50
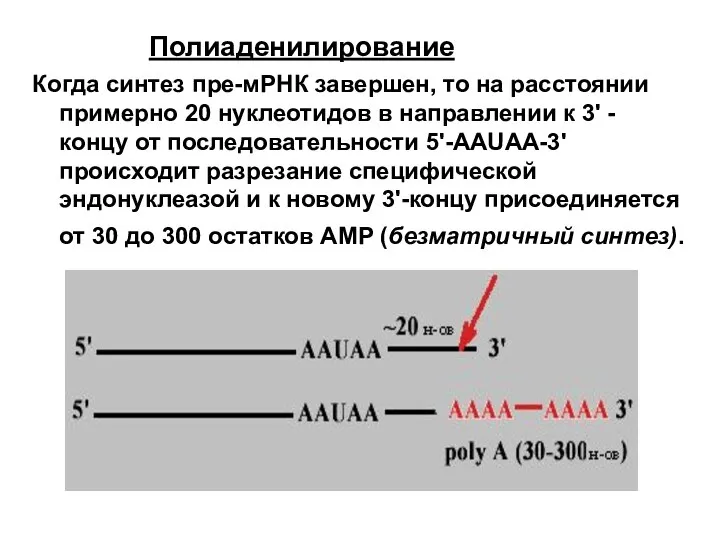
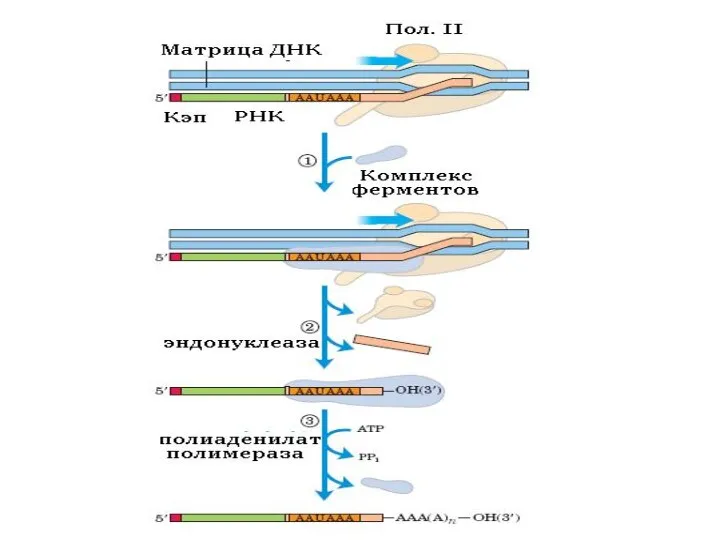
Полиаденилирование
Когда синтез пре-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в
Полиаденилирование
Когда синтез пре-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в
Слайд 52
мРНК ряда генов не полиаденилируется (например гистоновых генов).
Полиаденилированные пре-мРНК подвергаются сплайсингу.
мРНК ряда генов не полиаденилируется (например гистоновых генов).
Полиаденилированные пре-мРНК подвергаются сплайсингу.

Слайд 53
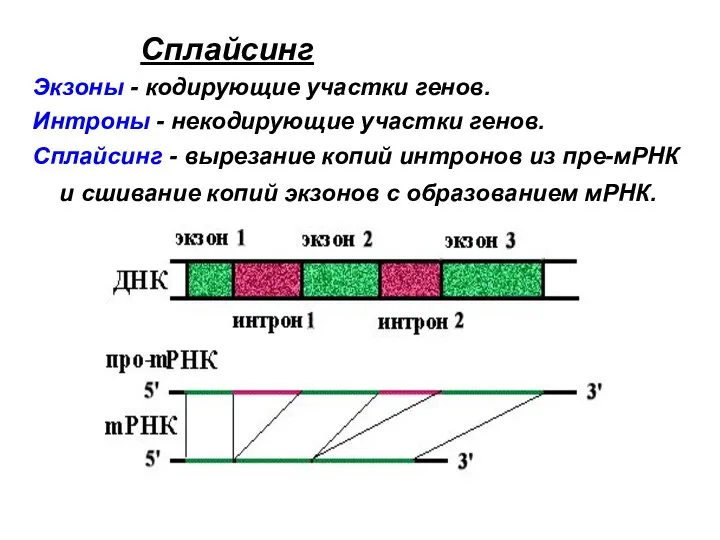
Сплайсинг
Экзоны - кодирующие участки генов.
Интроны - некодирующие участки генов.
Сплайсинг -
Сплайсинг
Экзоны - кодирующие участки генов.
Интроны - некодирующие участки генов.
Сплайсинг -
Слайд 54
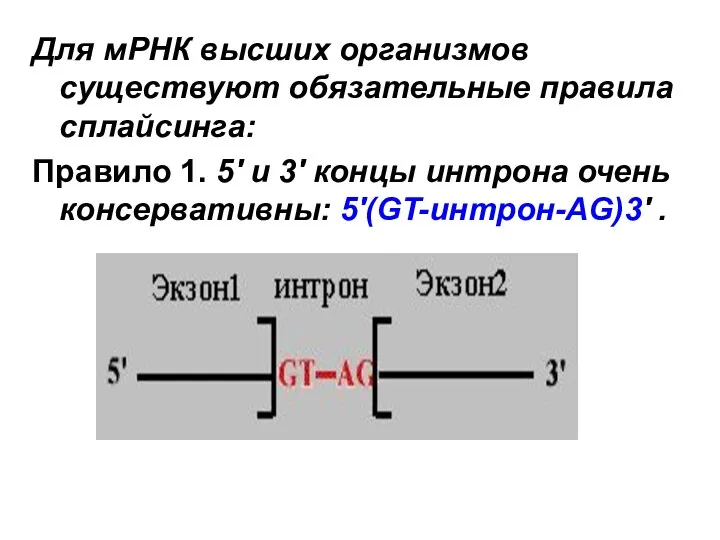
Для мРНК высших организмов существуют обязательные правила сплайсинга:
Правило 1. 5' и
Для мРНК высших организмов существуют обязательные правила сплайсинга:
Правило 1. 5' и
Слайд 55
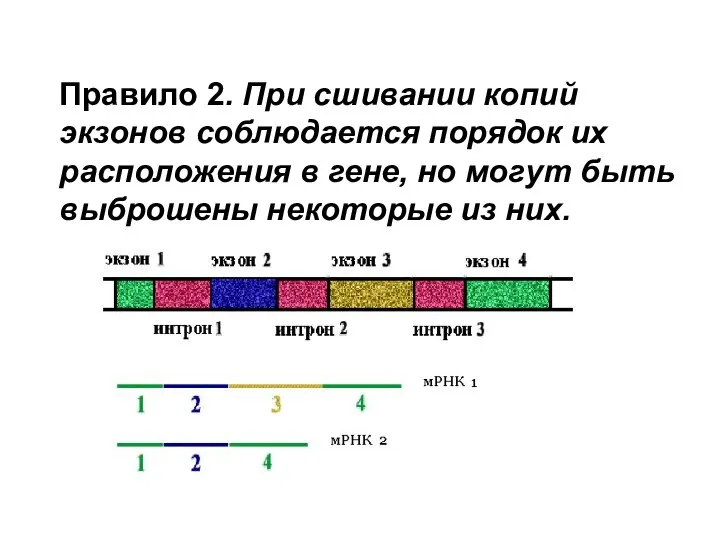
Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене,
Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене,
Слайд 56
Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в которых помимо ферментов, вырезающих и
Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в которых помимо ферментов, вырезающих и

Слайд 57
Редактирование
Редактирование - изменение генетической информации на уровне мРНК.
Пример- редактирование мРНК
Редактирование
Редактирование - изменение генетической информации на уровне мРНК.
Пример- редактирование мРНК

Слайд 58Процессинг РНК в клетках прокариот и эукариот
ЛЕКЦИЯ 9
Процессинг РНК в клетках прокариот и эукариот
ЛЕКЦИЯ 9

Слайд 59Этапы реализации генетической информации
Транскрипция – синтез молекул РНК, образование первичного транскрипта (пре-РНК)
Этапы реализации генетической информации
Транскрипция – синтез молекул РНК, образование первичного транскрипта (пре-РНК)

Слайд 60Процессинг РНК в клетках прокариот
Ген – это участок молекулы ДНК, кодирующий синтез
Процессинг РНК в клетках прокариот
Ген – это участок молекулы ДНК, кодирующий синтез

Слайд 61Процессинг рРНК и тРНК
Зрелые молекулы рРНК и тРНК образуются у прокариот и
Процессинг рРНК и тРНК
Зрелые молекулы рРНК и тРНК образуются у прокариот и

Слайд 62
Гены рРНК и тРНК образуют транскрипционный блок (кластер).
Спейсер – это участок
Гены рРНК и тРНК образуют транскрипционный блок (кластер).
Спейсер – это участок
Слайд 63Процессинг рРНК эукариот
Транскрипционный блок содержит гены 18S ; 5,8S; 28S - рРНК,
Процессинг рРНК эукариот
Транскрипционный блок содержит гены 18S ; 5,8S; 28S - рРНК,

Слайд 64
Процессинг пре-рРНК у эукариотов
Процессинг пре-рРНК у эукариотов
Слайд 65Этапы процессинга тРНК прокариот:
Модификация 5’ – конца - РНКаза Р
Модификацию 3’ –
Этапы процессинга тРНК прокариот:
Модификация 5’ – конца - РНКаза Р
Модификацию 3’ –
Слайд 66Этапы процессинга тРНК дрожжей (эукариоты)
Удаление интрона
Вырезание 5’ конца (лидер)
Удаление UU с 3’конца
Присоединение
Этапы процессинга тРНК дрожжей (эукариоты)
Удаление интрона
Вырезание 5’ конца (лидер)
Удаление UU с 3’конца
Присоединение
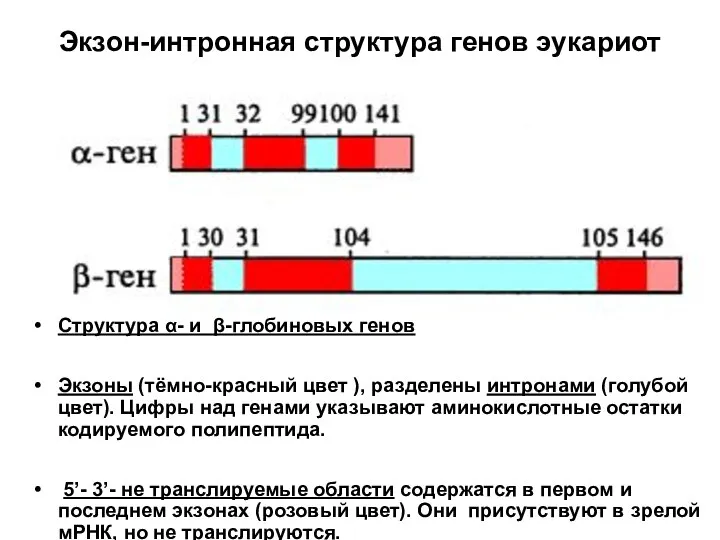
Слайд 67Экзон-интронная структура генов эукариот
Структура α- и β-глобиновых генов
Экзоны (тёмно-красный цвет ), разделены
Экзон-интронная структура генов эукариот
Структура α- и β-глобиновых генов
Экзоны (тёмно-красный цвет ), разделены
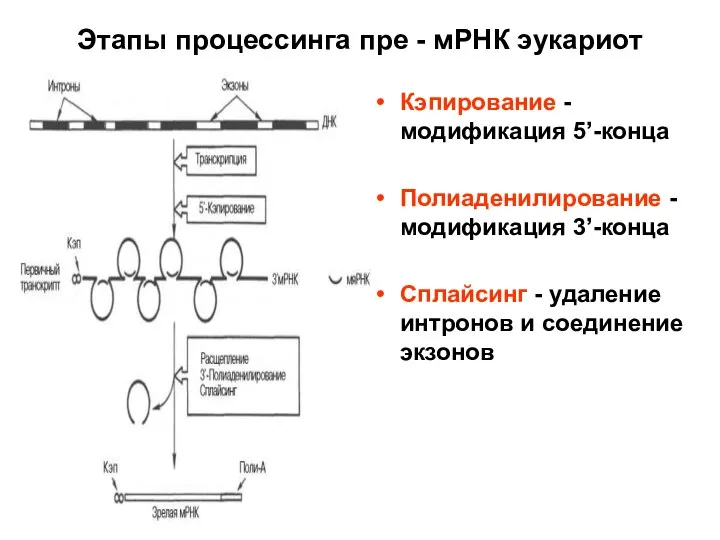
Слайд 68Этапы процессинга пре - мРНК эукариот
Кэпирование - модификация 5’-конца
Полиаденилирование - модификация 3’-конца
Сплайсинг
Этапы процессинга пре - мРНК эукариот
Кэпирование - модификация 5’-конца
Полиаденилирование - модификация 3’-конца
Сплайсинг
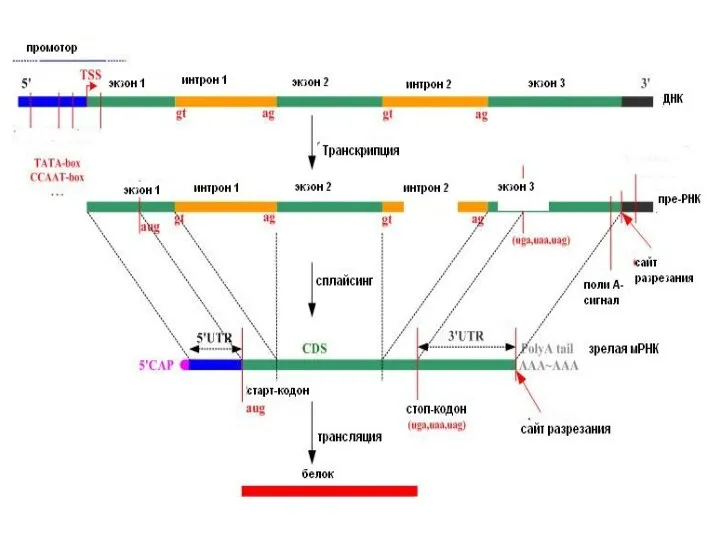
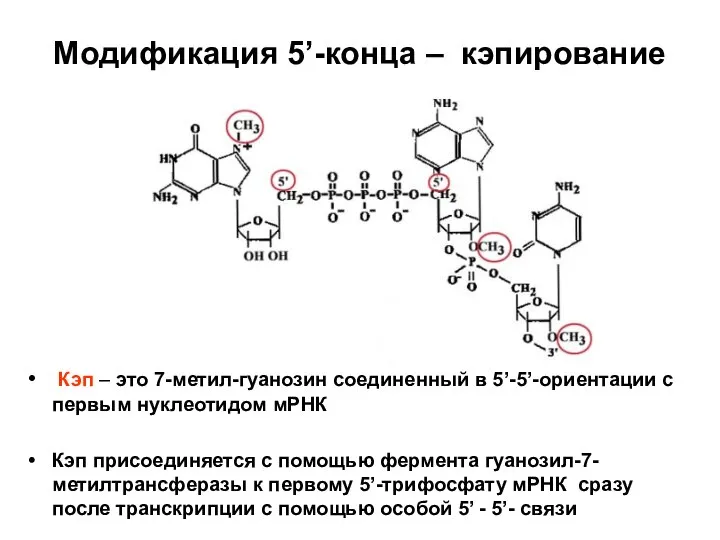
Слайд 70Модификация 5’-конца – кэпирование
Кэп – это 7-метил-гуанозин соединенный в 5’-5’-ориентации с
Модификация 5’-конца – кэпирование
Кэп – это 7-метил-гуанозин соединенный в 5’-5’-ориентации с
Слайд 71Модификация 3’-конца пре-мРНК - полиаденилирование
Модификация 3’-конца пре-мРНК - полиаденилирование
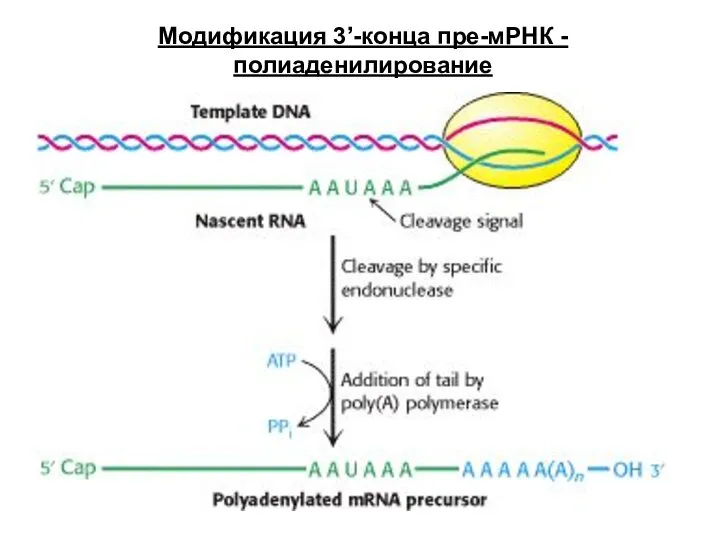
Слайд 72Механизмы сплайсинга интронов:
Тип I - интроны подвергаются аутосплайсингу в присутствии только ионов
Механизмы сплайсинга интронов:
Тип I - интроны подвергаются аутосплайсингу в присутствии только ионов

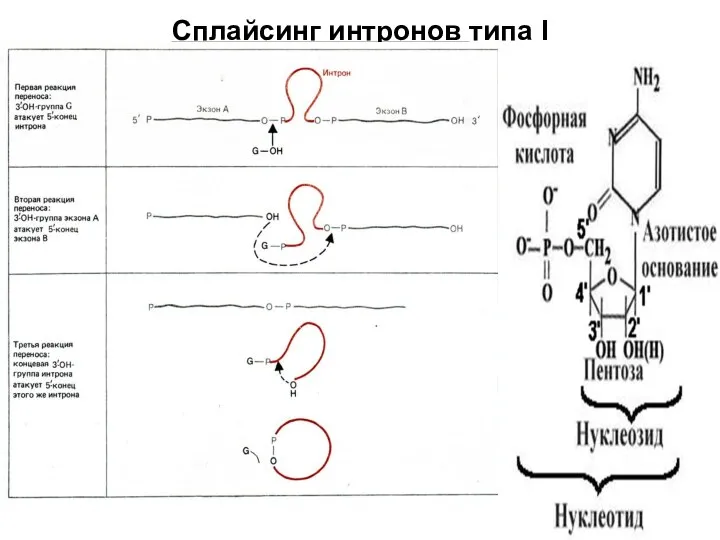
Слайд 73Сплайсинг интронов типа I
Сплайсинг интронов типа I
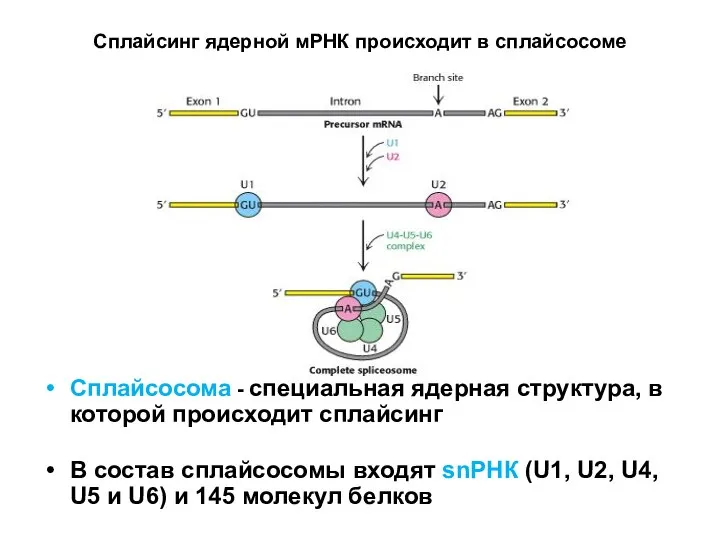
Слайд 74Сплайсинг ядерной мРНК происходит в сплайсосоме
Сплайсосома - специальная ядерная структура, в которой
Сплайсинг ядерной мРНК происходит в сплайсосоме
Сплайсосома - специальная ядерная структура, в которой
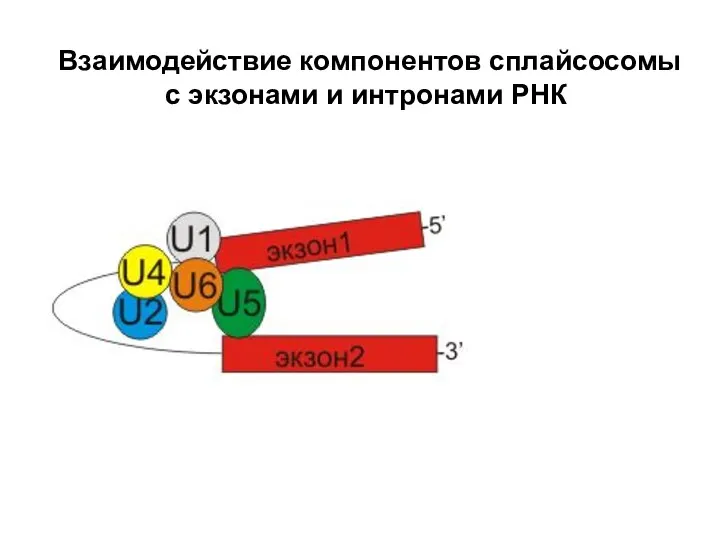
Слайд 75 Взаимодействие компонентов сплайсосомы с экзонами и интронами РНК
Взаимодействие компонентов сплайсосомы с экзонами и интронами РНК
Слайд 76Механизмы альтернативного сплайсинга:
Альтернативный выбор промотора
Альтернативный выбор сигнала полиаденилирования
Альтернативный выбор разных наборов экзонов
Транс-сплайсинг
Механизмы альтернативного сплайсинга:
Альтернативный выбор промотора
Альтернативный выбор сигнала полиаденилирования
Альтернативный выбор разных наборов экзонов
Транс-сплайсинг

Слайд 77Экзон 4
Интрон 3
Экзон 3
Интрон 2
Экзон 2
Р2
Интрон 1
Экзон 1
Р1
Экзон 3
Экзон 1
Экзон 4
Экзон 4
Экзон
Экзон 4
Интрон 3
Экзон 3
Интрон 2
Экзон 2
Р2
Интрон 1
Экзон 1
Р1
Экзон 3
Экзон 1
Экзон 4
Экзон 4
Экзон
Слайд 78
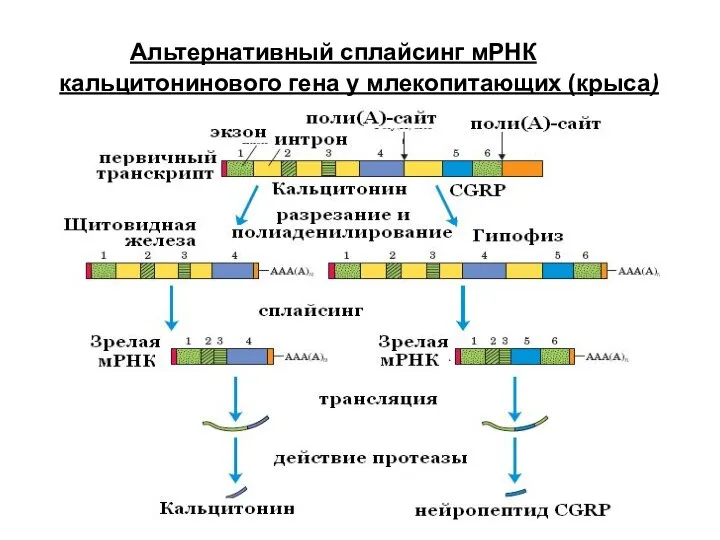
Альтернативный сплайсинг мРНК кальцитонинового гена у млекопитающих (крыса)
Альтернативный сплайсинг мРНК кальцитонинового гена у млекопитающих (крыса)
Слайд 79Структура мРНК прокариот
Лидер - это 5’ не транслируемый участок - 5’ UTR
Структура мРНК прокариот
Лидер - это 5’ не транслируемый участок - 5’ UTR
 Виды растений
Виды растений «ПОЛЯРНЫЕ ЖИВОТНЫЕ»
«ПОЛЯРНЫЕ ЖИВОТНЫЕ» Загадки о грибах. 5 класс
Загадки о грибах. 5 класс Отдел Папоротниковидные (тема 29)
Отдел Папоротниковидные (тема 29) Клеточный цикл. Митоз. Мейоз
Клеточный цикл. Митоз. Мейоз Ветреница Дубравная Anemone Nemorosa
Ветреница Дубравная Anemone Nemorosa Опыление растений
Опыление растений Тема урока «Комнатные растения в интерьере» 7 класс Тема урока «Комнатные растения
Тема урока «Комнатные растения в интерьере» 7 класс Тема урока «Комнатные растения  Профилактика отравлений грибами
Профилактика отравлений грибами Внешнее и внутреннее строение стебля растения
Внешнее и внутреннее строение стебля растения Строение зрительного анализатора
Строение зрительного анализатора Функции корня
Функции корня Биология - наука о живой природе
Биология - наука о живой природе Близнецовый метод изучения генетики
Близнецовый метод изучения генетики Биогеоценоз бөлік
Биогеоценоз бөлік Борьба с абиотическими факторами
Борьба с абиотическими факторами Селекция (часть 2)
Селекция (часть 2) Обобщающий урок по теме Анализаторы. Взаимодействие и взаимозаменяемость анализаторов
Обобщающий урок по теме Анализаторы. Взаимодействие и взаимозаменяемость анализаторов Общая микробиология. Лекция 1
Общая микробиология. Лекция 1 Lektsia_2
Lektsia_2 Адаптация к повышенному и пониженному барометрическому давлению
Адаптация к повышенному и пониженному барометрическому давлению Рыбы, гады и насекомые
Рыбы, гады и насекомые Цветоведение. Тест
Цветоведение. Тест Procesy energetyczne
Procesy energetyczne Нейруляция
Нейруляция Презентация на тему Поле и его обитатели
Презентация на тему Поле и его обитатели  Систематика и характеристика покрытосеменных. Подкласс Магнолииды (Magnoliidae)
Систематика и характеристика покрытосеменных. Подкласс Магнолииды (Magnoliidae) Карнотавр - род хищных динозавров
Карнотавр - род хищных динозавров