- Мегагаметофит. Лекция 1. О женском

Содержание
- 2. Что мы будем проходить? Мегаспорангий; пестик и семяпочка (лекция 1) Формирование женского гаметофита o Общие закономерности
- 3. Мегаспорангий Женский гаметофит семенных растений развивается в мегаспорангии (нуцеллусе) семяпочки. Другое название для семяпочки – семязачаток.
- 4. Мегаспороцит Внутри мегаспорангия проходит мейоз мегаспороцита (материнской клетки мегаспор) с образованием четырех мегаспор. У большинства растений
- 5. Семяпочки Семязача́ток, или семяпо́чка (лат. ovulum), — образование у семенных растений, из которого после оплодотворения развивается
- 6. Завязь и семяпочка Завязь – нижняя часть пестика. Внутри завязи развиваются семяпочки. Они закреплены с помощью
- 7. Рыльце и столбик Рыльце – верхняя, рецептивная часть пестика – собирает пыльцу и обеспечивает ее гидратацию
- 8. Разновидности рылец Выделяют два главных типа рылец: сухие и влажные, в зависимости от наличия на их
- 9. Клеточный состав мегагаметофита Зародышевый мешок (женский гаметофит) включает следующие клетки: две синергиды и яйцеклетку в микропилярном
- 10. Как работать с мегагаметофитом? Это объект, неудобный для экспериментальных исследований. Он труднодоступен, скрыт в толще тканей,
- 11. Выделение гаплоидных клеток
- 12. Микроманипуляции Manual Isolation of Living Cells from the Arabidopsis thaliana Female Gametophyte by Micromanipulation Sep 2017
- 13. Лазерная микродиссекция Под микроскопом с помощью лазера вырезают нужную область, затем она отделяется от основного среза
- 14. Развитие мегагаметофита Крупная функциональная мегаспора делится с образованием двухядерного ценоцита. Ядра в нем расходятся к противоположным
- 15. Как это выглядит
- 16. Женский гаметный модуль В результате целлюляризации зародышевого мешка возникает семиклеточная структура, объединяющая две гаметы, две синергиды
- 17. Яйцеклетка Яйцеклетка и синергиды обычно имеют грушевидную форму и полярную структуру. Ядро яйцеклетки часто располагается в
- 18. Cинергиды Полярность синергид обратная по отношению к яйцеклетке: ядро занимает центральную позицию, а вакуоль (вакуоли) смещены
- 19. Центральная клетка Центральная клетка сильно вакуолизирована и занимает большую часть ЗМ. Периферическая цитоплазма соединяется с перинуклеарным
- 20. Антиподы Антиподальный комплекс характеризуется большим видовым разнообразием. У Arabidopsis антиподы не обнаруживают признаков специализации, и принято
- 21. Синхронизация У большинства видов покрытосеменных растений женский гаметофит в основном формируется до опыления. Однако известны растения,
- 22. Спецификация клеток Разная судьба: 2 гаметы (ЯК, ЦК), 2 типа вспомогательных клеток (С, А) Ориентация митозов
- 23. Мутанты и гены
- 24. Мутанты по спецификации Мутант кукурузы с избыточной пролиферацией ядер (ig1, indeterminate gametophyte1) Мутант Arabidopsis с нарушением
- 25. Мутанты по спецификации У мутанта Arabidopsis eostre число ядер в зародышевом мешке нормальное, но на четырехядерной
- 26. Итак, что же в норме? В ходе нормального развития зародышевого мешка ядра, ближайшие к микропиле, окажутся
- 27. Позиционная информация Анализ мутантов показал ключевую роль позиционной информации в определении судьбы клеток женского гаметофита →между
- 28. Позиционная информация и ауксин До митозов ауксин в больших количествах присутствовал в нуцеллусе, но его не
- 29. Ауксиновый градиент Ауксиновый градиент был обнаружен в нормальных зародышевых мешках Гетерогенный синтез ауксина Полярное распределение переносчиков
- 30. Мутанты У мутантов с избыточным биосинтезом ауксина (оверэкспрессия гена YUC1) расположение ядер в зародышевом мешке было
- 31. Мутанты При выключении ауксин-зависимых транскрипционных факторов синергиды в ЗМ не появлялись, концентрация ауксина «дотягивала» только до
- 32. Мутанты У мутантов по гену altered meristem program 1 ( amp1) синергиды превращаются в ЯК. Белок
- 33. Межклеточные взаимодействия в зародышевом мешке Обнаружены три гена, контролирующие судьбу клеток женского гаметофита после целлюляризации: LACHESIS
- 34. Латеральное ингибирование такой тип межклеточных взаимодействий, в ходе которого клетка, судьба которой определилась, не позволяет соседним
- 35. Ronny Völz et al. Development 2012;139:498-502 © 2012. Мутант по латеральному ингибированию LIS::2xGFP_LIS Белок LIS в
- 36. Ronny Völz et al. Development 2012;139:498-502 © 2012. Как это себе представляют? Гены, ответственные за латеральное
- 37. Центральная клетка убивает антиподы Мутация fiona ⁄ syco-1 влияет на целостность митохондрий центральной клетки, а также
- 38. Как приходит сигнал? Растительные клетки могут взаимодействовать, обмениваясь сигналами с участием лиганд-рецепторных систем, или посредством прямых
- 39. Главная – яйцеклетка Это наглядно показали эксперименты, в которых с помощью РНК-интерференции избирательно снижали число транскриптов
- 40. Что за сигнал? Похоже, пептиды Химическая природа сигналов, продуцируемых яйцеклеткой и центральной клеткой Arabidopsis, активно изучается.
- 41. Схематично
- 43. Скачать презентацию
Слайд 2Что мы будем проходить?
Мегаспорангий; пестик и семяпочка (лекция 1)
Формирование женского гаметофита
o Общие
Что мы будем проходить?
Мегаспорангий; пестик и семяпочка (лекция 1)
Формирование женского гаметофита
o Общие

Слайд 3Мегаспорангий
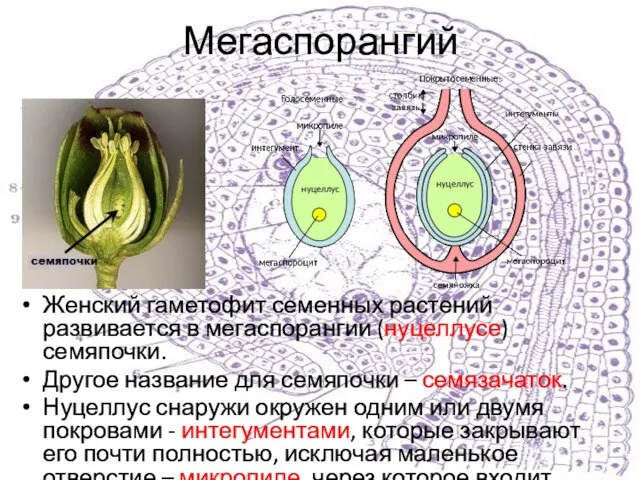
Женский гаметофит семенных растений развивается в мегаспорангии (нуцеллусе) семяпочки.
Другое название для
Мегаспорангий
Женский гаметофит семенных растений развивается в мегаспорангии (нуцеллусе) семяпочки.
Другое название для
Слайд 4Мегаспороцит
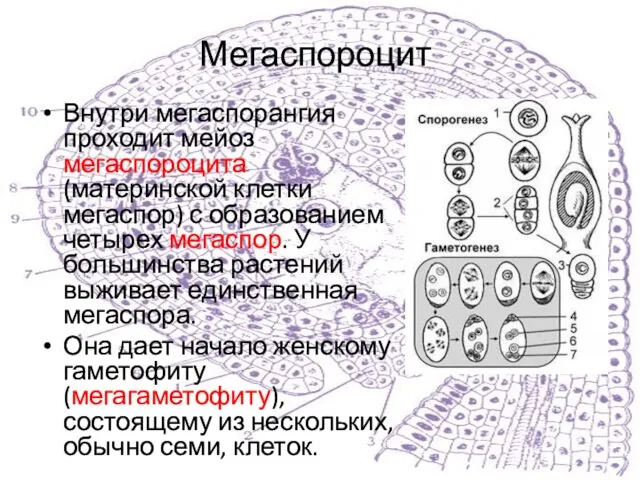
Внутри мегаспорангия проходит мейоз мегаспороцита (материнской клетки мегаспор) с образованием четырех мегаспор.
Мегаспороцит
Внутри мегаспорангия проходит мейоз мегаспороцита (материнской клетки мегаспор) с образованием четырех мегаспор.
Слайд 5Семяпочки
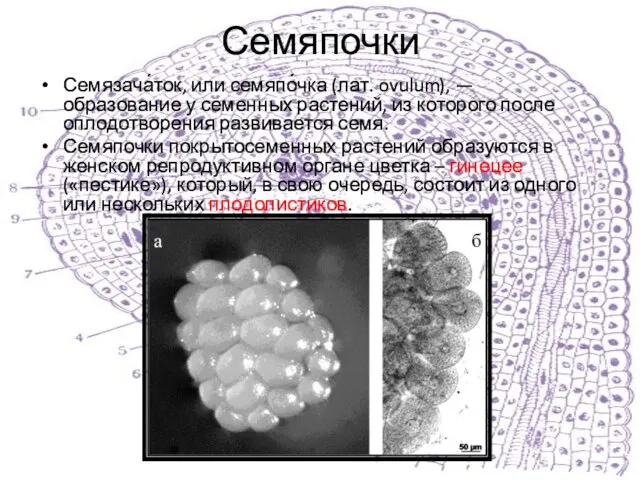
Семязача́ток, или семяпо́чка (лат. ovulum), — образование у семенных растений, из которого после оплодотворения
Семяпочки
Семязача́ток, или семяпо́чка (лат. ovulum), — образование у семенных растений, из которого после оплодотворения
Слайд 6Завязь и семяпочка
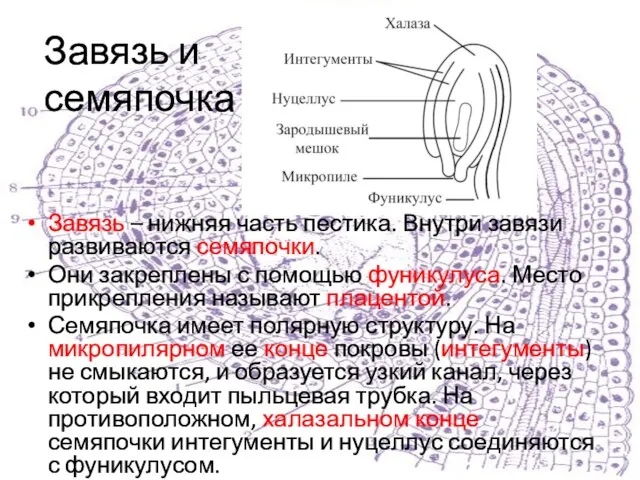
Завязь – нижняя часть пестика. Внутри завязи развиваются семяпочки.
Они
Завязь и семяпочка
Завязь – нижняя часть пестика. Внутри завязи развиваются семяпочки.
Они
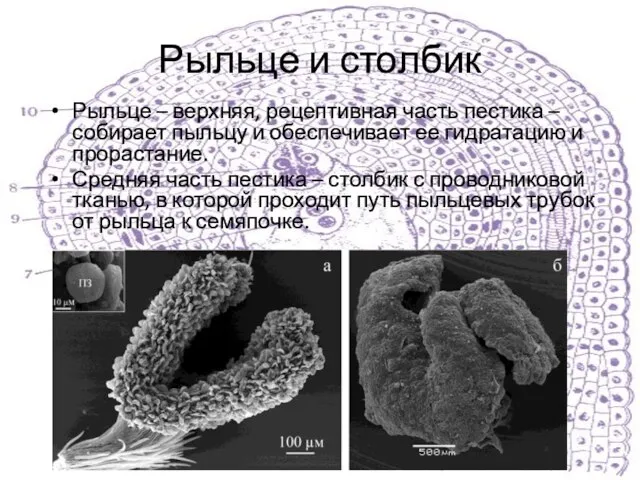
Слайд 7Рыльце и столбик
Рыльце – верхняя, рецептивная часть пестика – собирает пыльцу и
Рыльце и столбик
Рыльце – верхняя, рецептивная часть пестика – собирает пыльцу и
Слайд 8Разновидности рылец
Выделяют два главных типа рылец: сухие и влажные, в зависимости от
Разновидности рылец
Выделяют два главных типа рылец: сухие и влажные, в зависимости от

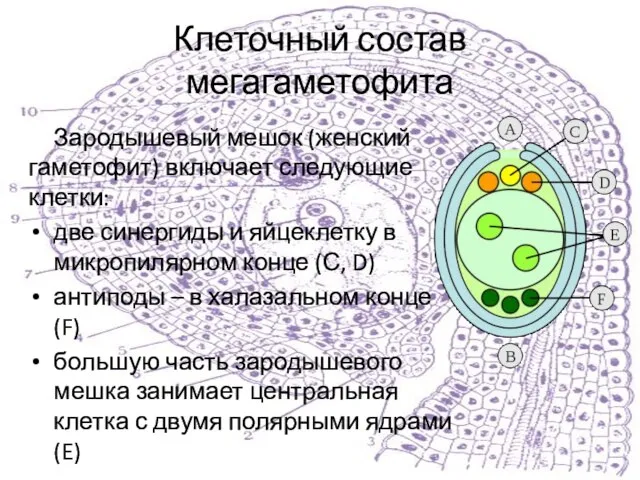
Слайд 9Клеточный состав мегагаметофита
Зародышевый мешок (женский гаметофит) включает следующие клетки:
две синергиды и
Клеточный состав мегагаметофита
Зародышевый мешок (женский гаметофит) включает следующие клетки:
две синергиды и
Слайд 10Как работать с мегагаметофитом?
Это объект, неудобный для экспериментальных исследований. Он труднодоступен, скрыт
Как работать с мегагаметофитом?
Это объект, неудобный для экспериментальных исследований. Он труднодоступен, скрыт
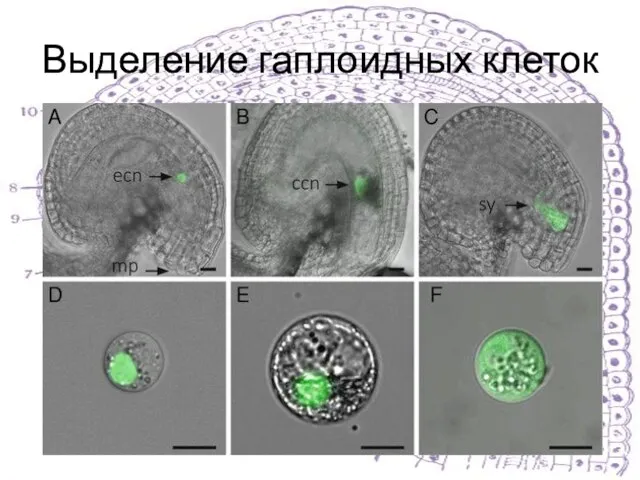
Слайд 11Выделение гаплоидных клеток
Выделение гаплоидных клеток
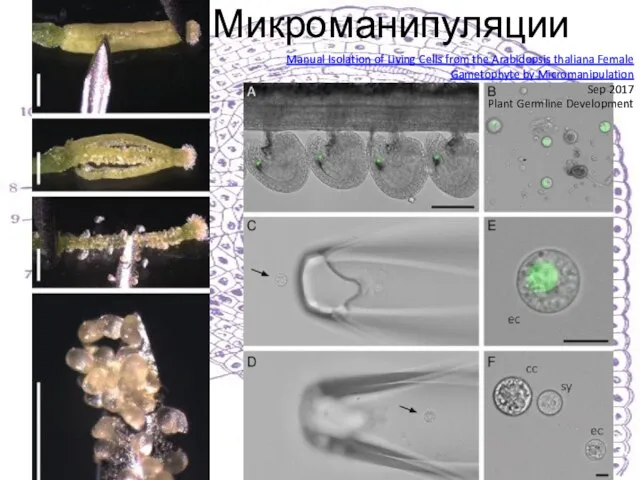
Слайд 12Микроманипуляции
Manual Isolation of Living Cells from the Arabidopsis thaliana Female Gametophyte by
Микроманипуляции
Manual Isolation of Living Cells from the Arabidopsis thaliana Female Gametophyte by
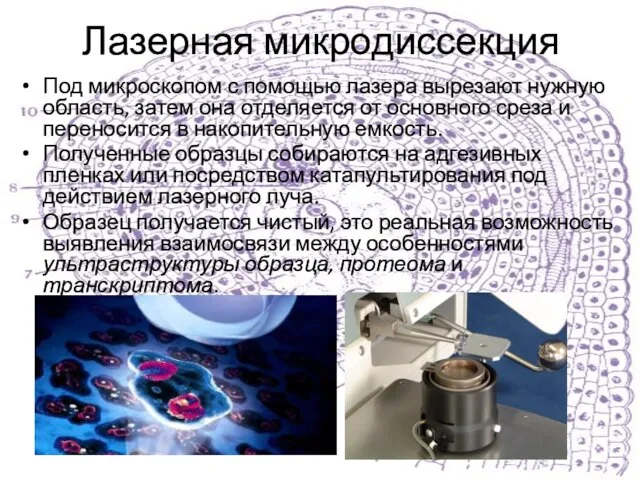
Слайд 13Лазерная микродиссекция
Под микроскопом с помощью лазера вырезают нужную область, затем она отделяется
Лазерная микродиссекция
Под микроскопом с помощью лазера вырезают нужную область, затем она отделяется
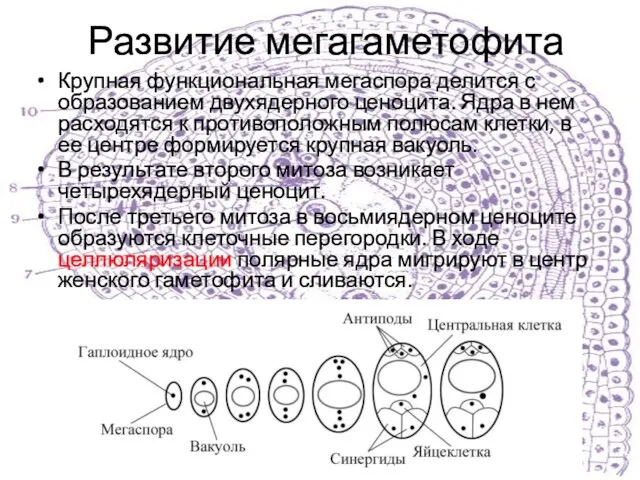
Слайд 14Развитие мегагаметофита
Крупная функциональная мегаспора делится с образованием двухядерного ценоцита. Ядра в нем
Развитие мегагаметофита
Крупная функциональная мегаспора делится с образованием двухядерного ценоцита. Ядра в нем
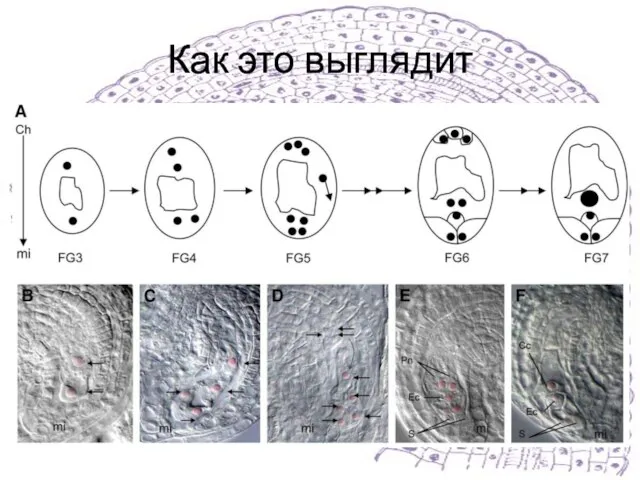
Слайд 15Как это выглядит
Как это выглядит
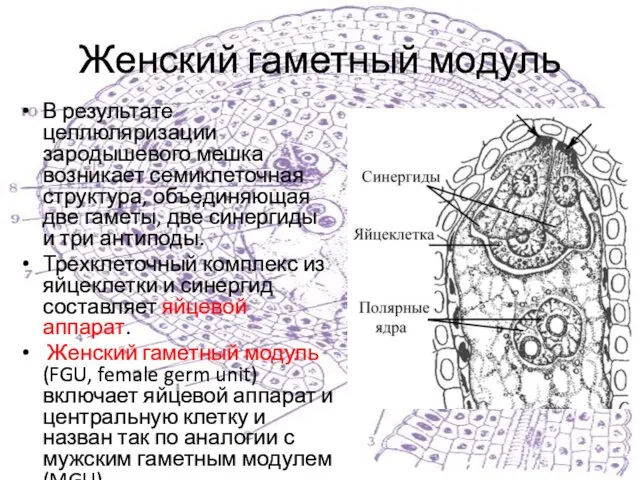
Слайд 16Женский гаметный модуль
В результате целлюляризации зародышевого мешка возникает семиклеточная структура, объединяющая две
Женский гаметный модуль
В результате целлюляризации зародышевого мешка возникает семиклеточная структура, объединяющая две
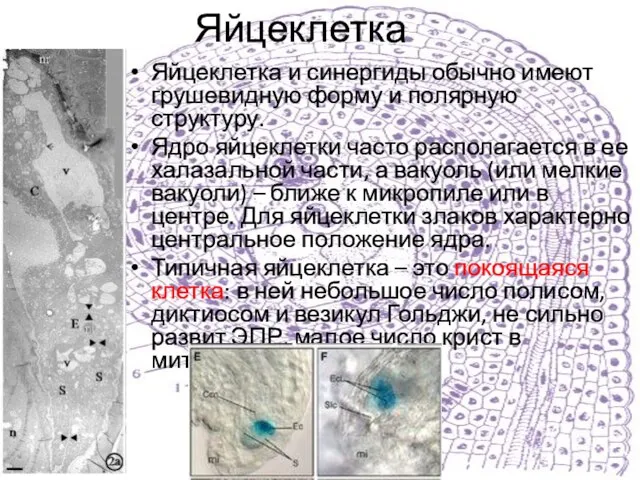
Слайд 17Яйцеклетка
Яйцеклетка и синергиды обычно имеют грушевидную форму и полярную структуру.
Ядро яйцеклетки
Яйцеклетка
Яйцеклетка и синергиды обычно имеют грушевидную форму и полярную структуру.
Ядро яйцеклетки
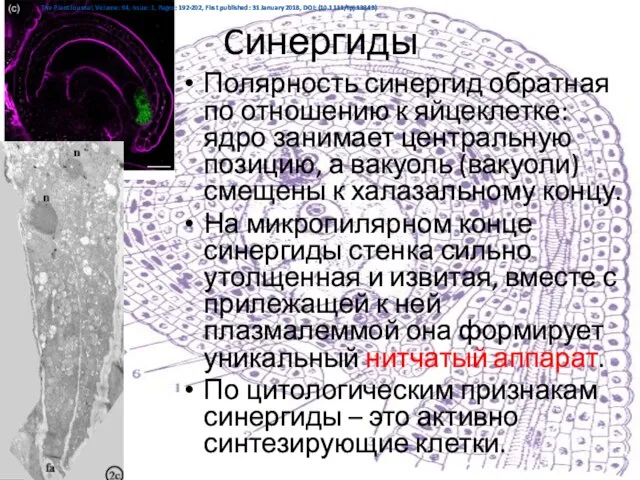
Слайд 18Cинергиды
Полярность синергид обратная по отношению к яйцеклетке: ядро занимает центральную позицию, а
Cинергиды
Полярность синергид обратная по отношению к яйцеклетке: ядро занимает центральную позицию, а
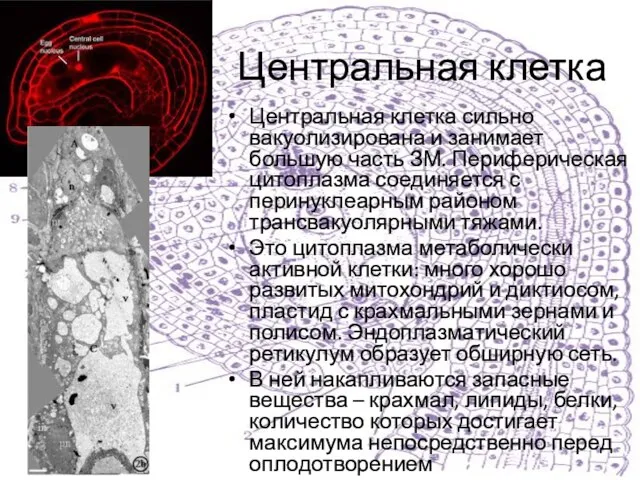
Слайд 19Центральная клетка
Центральная клетка сильно вакуолизирована и занимает большую часть ЗМ. Периферическая цитоплазма
Центральная клетка
Центральная клетка сильно вакуолизирована и занимает большую часть ЗМ. Периферическая цитоплазма
Слайд 20Антиподы
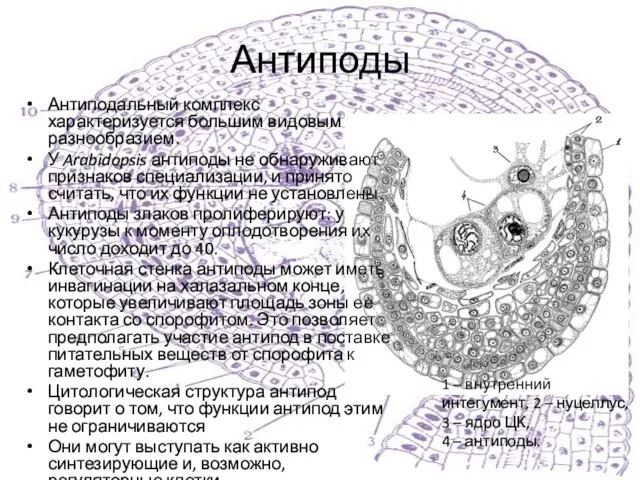
Антиподальный комплекс характеризуется большим видовым разнообразием.
У Arabidopsis антиподы не обнаруживают признаков
Антиподы
Антиподальный комплекс характеризуется большим видовым разнообразием.
У Arabidopsis антиподы не обнаруживают признаков
Слайд 21Синхронизация
У большинства видов покрытосеменных растений женский гаметофит в основном формируется до опыления.
Синхронизация
У большинства видов покрытосеменных растений женский гаметофит в основном формируется до опыления.

Слайд 22Спецификация клеток
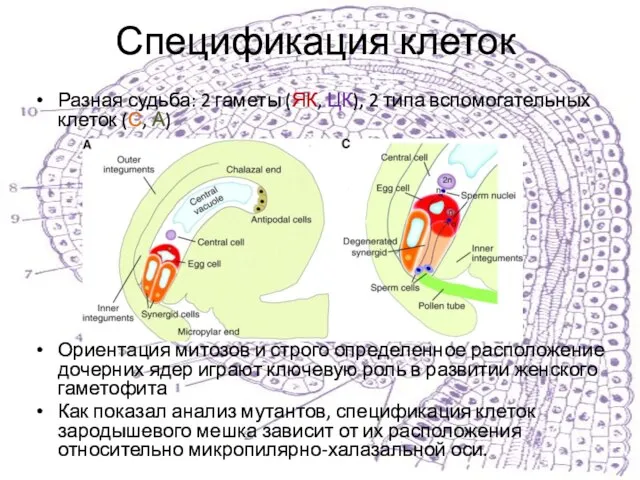
Разная судьба: 2 гаметы (ЯК, ЦК), 2 типа вспомогательных клеток (С,
Спецификация клеток
Разная судьба: 2 гаметы (ЯК, ЦК), 2 типа вспомогательных клеток (С,
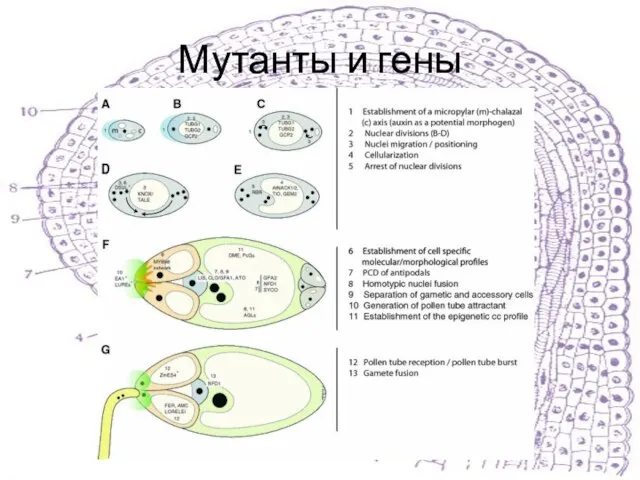
Слайд 23Мутанты и гены
Мутанты и гены
Слайд 24Мутанты по спецификации
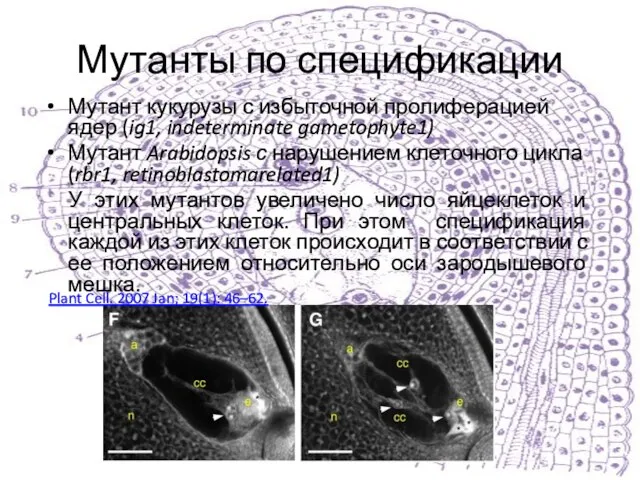
Мутант кукурузы с избыточной пролиферацией ядер (ig1, indeterminate gametophyte1)
Мутант
Мутанты по спецификации
Мутант кукурузы с избыточной пролиферацией ядер (ig1, indeterminate gametophyte1)
Мутант
Слайд 25Мутанты по спецификации
У мутанта Arabidopsis eostre число ядер в зародышевом мешке нормальное,
Мутанты по спецификации
У мутанта Arabidopsis eostre число ядер в зародышевом мешке нормальное,
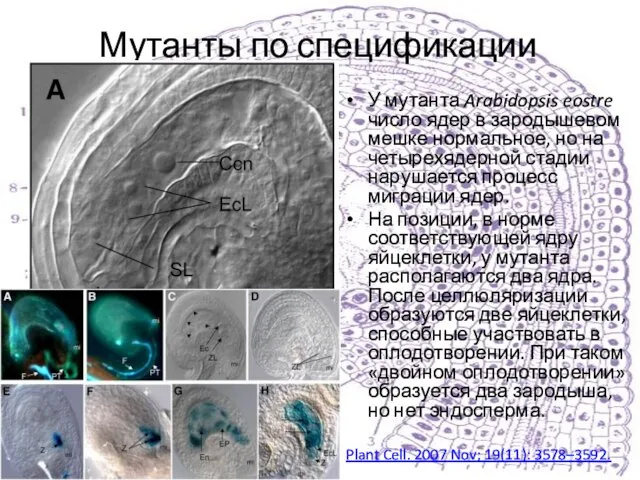
Слайд 26Итак, что же в норме?
В ходе нормального развития зародышевого мешка ядра, ближайшие
Итак, что же в норме?
В ходе нормального развития зародышевого мешка ядра, ближайшие

Слайд 27Позиционная информация
Анализ мутантов показал ключевую роль позиционной информации в определении судьбы клеток
Позиционная информация
Анализ мутантов показал ключевую роль позиционной информации в определении судьбы клеток
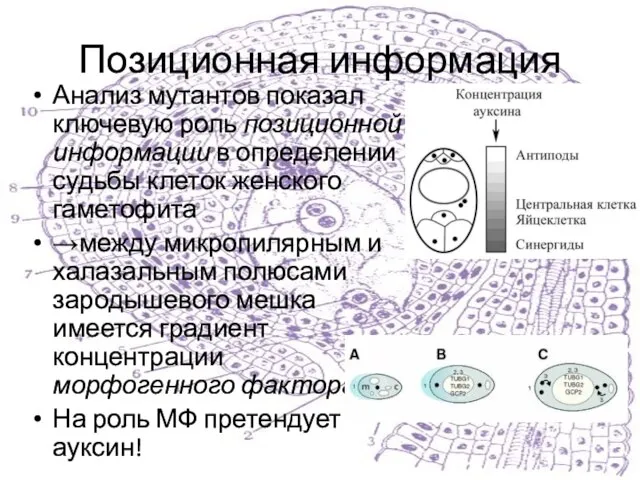
Слайд 28Позиционная информация и ауксин
До митозов ауксин в больших количествах присутствовал в нуцеллусе,
Позиционная информация и ауксин
До митозов ауксин в больших количествах присутствовал в нуцеллусе,
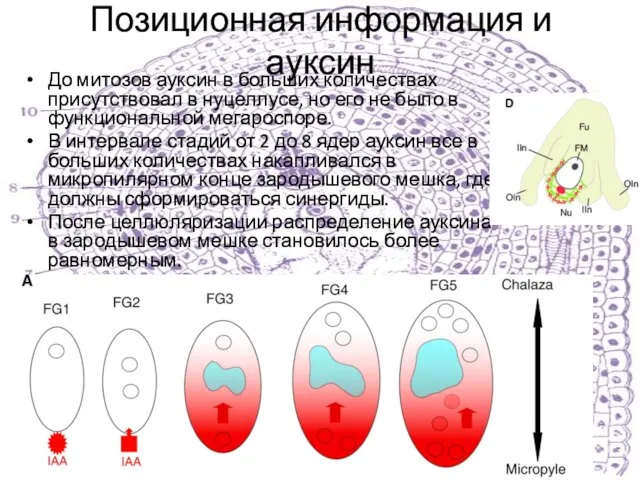
Слайд 29Ауксиновый градиент
Ауксиновый градиент был обнаружен в нормальных зародышевых мешках
Гетерогенный синтез ауксина
Полярное распределение
Ауксиновый градиент
Ауксиновый градиент был обнаружен в нормальных зародышевых мешках
Гетерогенный синтез ауксина
Полярное распределение
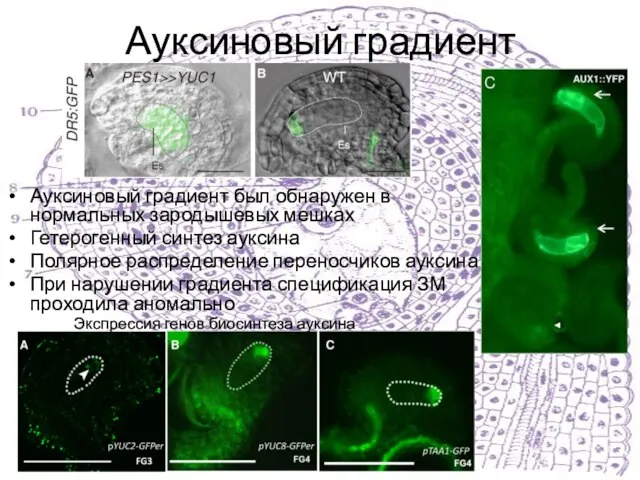
Слайд 30Мутанты
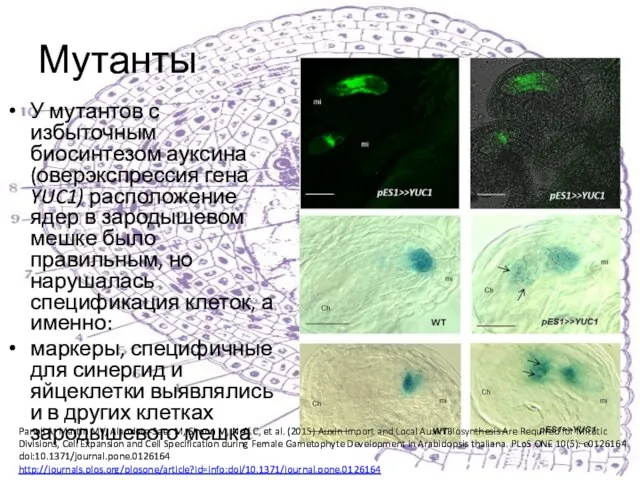
У мутантов с избыточным биосинтезом ауксина (оверэкспрессия гена YUC1) расположение ядер в
Мутанты
У мутантов с избыточным биосинтезом ауксина (оверэкспрессия гена YUC1) расположение ядер в
Слайд 31Мутанты
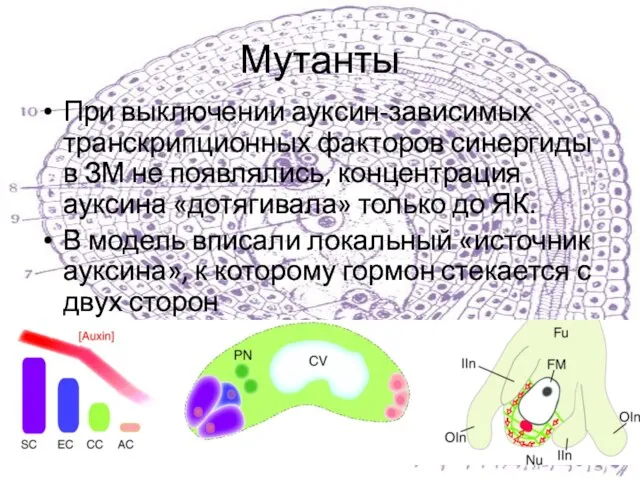
При выключении ауксин-зависимых транскрипционных факторов синергиды в ЗМ не появлялись, концентрация ауксина
Мутанты
При выключении ауксин-зависимых транскрипционных факторов синергиды в ЗМ не появлялись, концентрация ауксина
Слайд 32 Мутанты
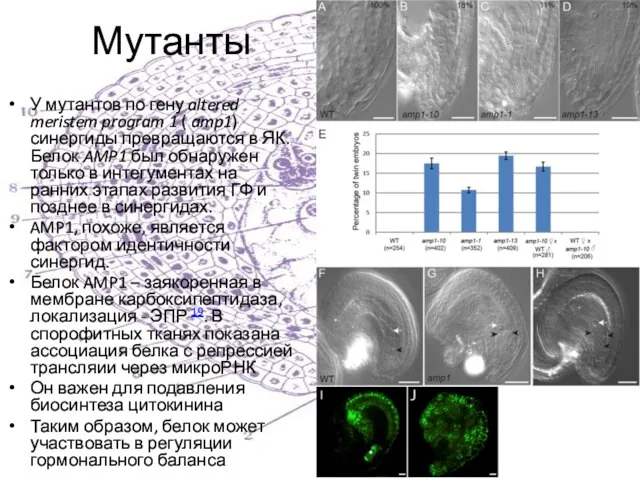
У мутантов по гену altered meristem program 1 ( amp1) синергиды
Мутанты
У мутантов по гену altered meristem program 1 ( amp1) синергиды
Слайд 33Межклеточные взаимодействия в зародышевом мешке
Обнаружены три гена, контролирующие судьбу клеток женского гаметофита
Межклеточные взаимодействия в зародышевом мешке
Обнаружены три гена, контролирующие судьбу клеток женского гаметофита

Слайд 34Латеральное ингибирование
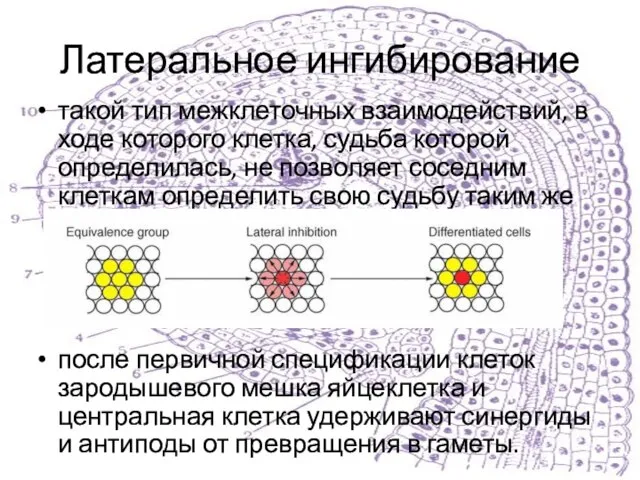
такой тип межклеточных взаимодействий, в ходе которого клетка, судьба которой определилась,
Латеральное ингибирование
такой тип межклеточных взаимодействий, в ходе которого клетка, судьба которой определилась,
Слайд 35Ronny Völz et al. Development 2012;139:498-502
© 2012.
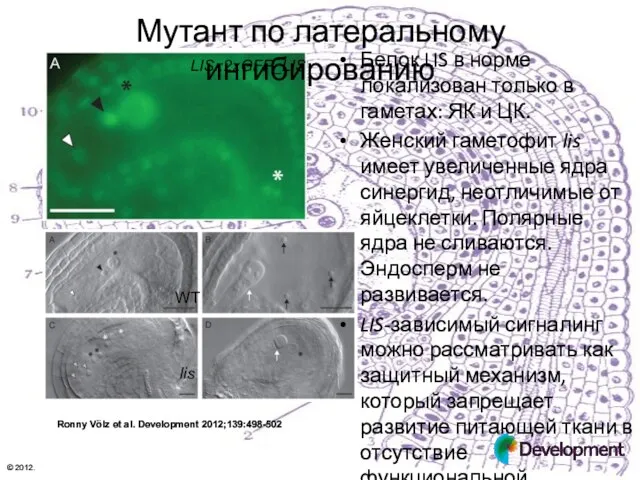
Мутант по латеральному ингибированию
LIS::2xGFP_LIS
Белок LIS в
Ronny Völz et al. Development 2012;139:498-502
© 2012.
Мутант по латеральному ингибированию
LIS::2xGFP_LIS
Белок LIS в
Слайд 36Ronny Völz et al. Development 2012;139:498-502
© 2012.
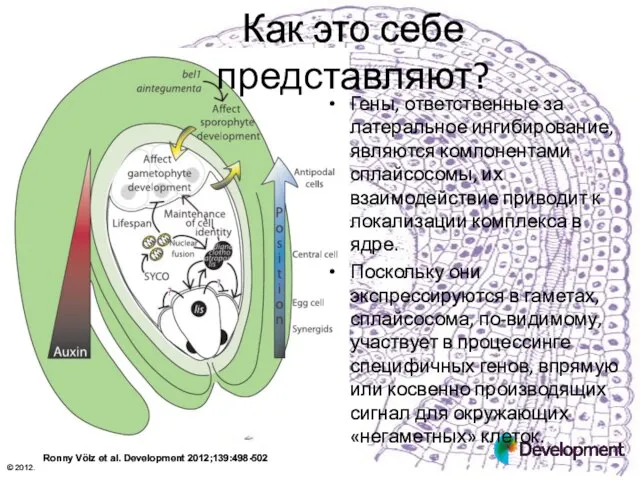
Как это себе представляют?
Гены, ответственные за
Ronny Völz et al. Development 2012;139:498-502
© 2012.
Как это себе представляют?
Гены, ответственные за
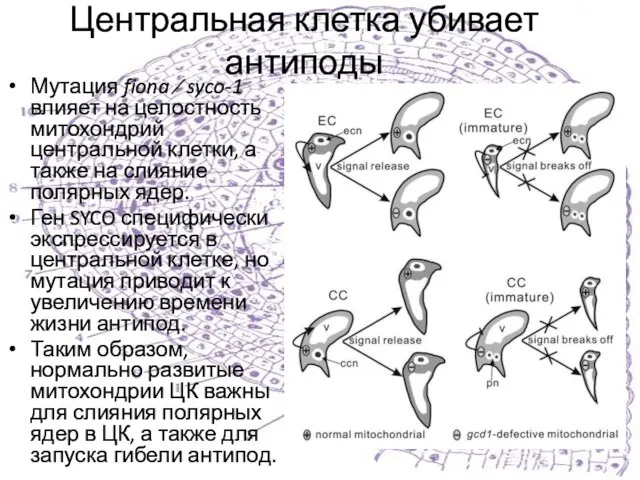
Слайд 37Центральная клетка убивает антиподы
Мутация fiona ⁄ syco-1 влияет на целостность митохондрий центральной
Центральная клетка убивает антиподы
Мутация fiona ⁄ syco-1 влияет на целостность митохондрий центральной
Слайд 38Как приходит сигнал?
Растительные клетки могут взаимодействовать, обмениваясь сигналами с участием лиганд-рецепторных систем,
Как приходит сигнал?
Растительные клетки могут взаимодействовать, обмениваясь сигналами с участием лиганд-рецепторных систем,

Слайд 39Главная – яйцеклетка
Это наглядно показали эксперименты, в которых с помощью РНК-интерференции
Главная – яйцеклетка
Это наглядно показали эксперименты, в которых с помощью РНК-интерференции

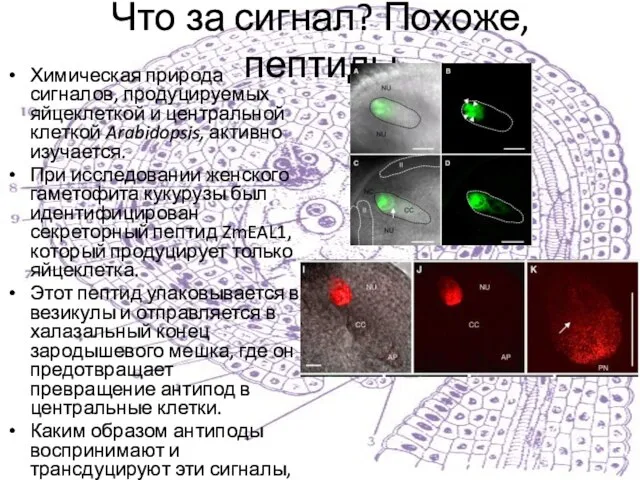
Слайд 40Что за сигнал? Похоже, пептиды
Химическая природа сигналов, продуцируемых яйцеклеткой и центральной клеткой
Что за сигнал? Похоже, пептиды
Химическая природа сигналов, продуцируемых яйцеклеткой и центральной клеткой
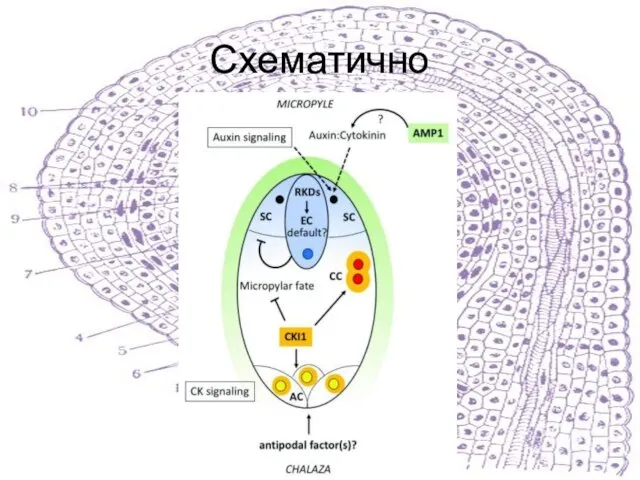
Слайд 41Схематично
Схематично
 Who wants to be a millionaire
Who wants to be a millionaire Побег: развитие разнообразие, строение
Побег: развитие разнообразие, строение 13. Покрытосеменные растения
13. Покрытосеменные растения According to the soil specialization (And some botanical science…)
According to the soil specialization (And some botanical science…) Фотосинтез. Историческая справка
Фотосинтез. Историческая справка Травоядные динозавры
Травоядные динозавры Как ферменты ускоряют химические реакции
Как ферменты ускоряют химические реакции Гормоны
Гормоны Митохондриальная наследственность
Митохондриальная наследственность Современная зоология
Современная зоология Австралія
Австралія Семейства двудольных растений
Семейства двудольных растений Пищеварительная система человека
Пищеварительная система человека Сравнительная характеристика типов червей
Сравнительная характеристика типов червей Биология в средневековье. (Лекция 3)
Биология в средневековье. (Лекция 3) Видоизменения листьев
Видоизменения листьев Тварини. Свійські тварини. Дикі тварини
Тварини. Свійські тварини. Дикі тварини Para qué son Estes partes de cuerpo humano
Para qué son Estes partes de cuerpo humano Мероприятие для 7-9-х классов Северный Ледовитый океан: экология, история, современность
Мероприятие для 7-9-х классов Северный Ледовитый океан: экология, история, современность Наследственность изменчивость. История развития генетики
Наследственность изменчивость. История развития генетики Презентация на тему Пионеры растительности
Презентация на тему Пионеры растительности  Тип Членистоногие
Тип Членистоногие Грудная и брюшная части аорты
Грудная и брюшная части аорты Презентация на тему Класс Земноводные
Презентация на тему Класс Земноводные  Корень
Корень Свойства биосистем
Свойства биосистем Периферический скелет домашних животных
Периферический скелет домашних животных Химический состав клетки
Химический состав клетки