- Механізми утворення різноманітності антитіл i вклітинних рецепторів

Содержание
- 2. В нормі клітина має експресувати лише один тип функціонально перебудованих генів важкого й легкого ланцюгів (явище
- 3. Алельне виключення генів імуноглобулінів можна пояснити механічними перешкодами для одночасної деконденсації хроматину двох гомологічних хромосом, яке
- 4. Молекулярні механізми перебудови генів антитіл та ВкР. Молекулярні механізми з’єднання V- і J-сегментів генів легких ланцюгів
- 5. Розташування спейсерів та RSS-послідовностей (від англ. rесотbіnаtion signal siquences)- фланкувальні послідовності, що оточують V-, D- і
- 6. RAG1 і RAG2 продукти генів rag1 і rag2 (від англ. rесотbіnаtіоn activity genes ), активуються лише
- 7. Антигеннезалежна перебудова генів імуноглобулінів призводить до формування функціональних ІgМ- та ІgD-рецепторів наївних В-клітин. Антигензалежна – до
- 8. Соматичні гіпермутації в генах імуноглобулінів. Створення різноманітності антитіл може бути результатом точкових соматичних мутацій та "помилок"
- 9. Соматичні мутації практично не відбуваються в ділянках, що кодують сталі ділянки імуноглобулінових ланцюгів. Мутації виникають як
- 10. Процеси соматичного гіпермутагенезу залежать від ферменту - індукованої активацією дезамінази цитидину (AICD від англ. activation induced
- 11. "Помилки" під час рекомбінації. місця з'єднання V-D , D-J і V-J-сегментів можуть зміщуватися в частину 5'-
- 12. ОРГАНІЗАЦІЯ I ПЕРЕБУДОВА ГЕНІВ Т-КЛІТИННИХ РЕЦЕПТОРІВ.
- 13. Організація генів Т-клітинного рецептора людини (на малюнку поданоорганізмацію локусів генів ланцюгів α, δ, β, γ, які
- 15. Скачать презентацию

Слайд 3Алельне виключення генів імуноглобулінів можна пояснити механічними перешкодами для одночасної деконденсації хроматину

Слайд 4Молекулярні механізми перебудови генів антитіл та ВкР.
Молекулярні механізми з’єднання V- і J-сегментів
Молекулярні механізми перебудови генів антитіл та ВкР.
Молекулярні механізми з’єднання V- і J-сегментів

Слайд 5
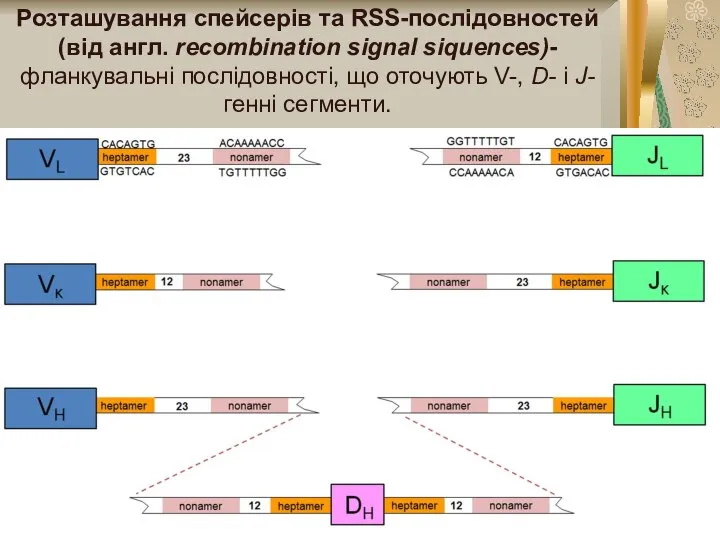
Розташування спейсерів та RSS-послідовностей (від англ. rесотbіnаtion signal siquences)- фланкувальні послідовності, що
Розташування спейсерів та RSS-послідовностей (від англ. rесотbіnаtion signal siquences)- фланкувальні послідовності, що
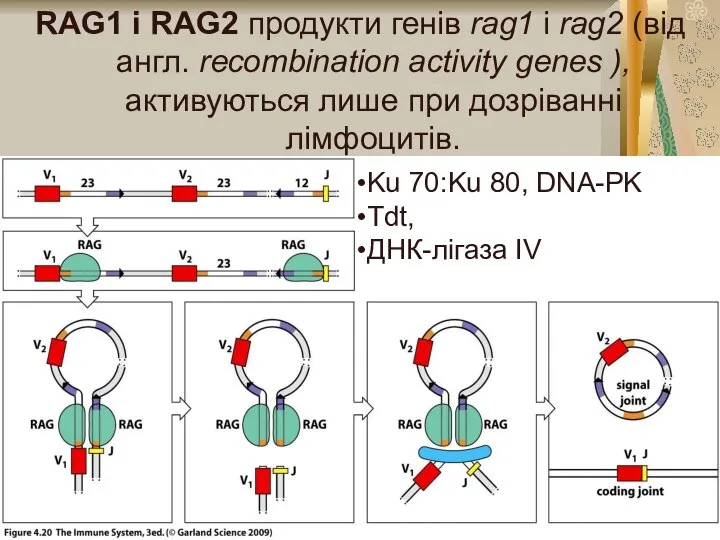
Слайд 6RAG1 і RAG2 продукти генів rag1 і rag2 (від англ. rесотbіnаtіоn activity
RAG1 і RAG2 продукти генів rag1 і rag2 (від англ. rесотbіnаtіоn activity
Слайд 7Антигеннезалежна перебудова генів імуноглобулінів призводить до формування функціональних ІgМ- та ІgD-рецепторів наївних
Антигеннезалежна перебудова генів імуноглобулінів призводить до формування функціональних ІgМ- та ІgD-рецепторів наївних

Слайд 8Соматичні гіпермутації в генах імуноглобулінів.
Створення різноманітності антитіл може бути результатом точкових
Соматичні гіпермутації в генах імуноглобулінів.
Створення різноманітності антитіл може бути результатом точкових

Слайд 9Соматичні мутації практично не відбуваються в ділянках, що кодують сталі ділянки імуноглобулінових

Слайд 10Процеси соматичного гіпермутагенезу залежать від ферменту - індукованої активацією дезамінази цитидину (AICD
Процеси соматичного гіпермутагенезу залежать від ферменту - індукованої активацією дезамінази цитидину (AICD

Слайд 11"Помилки" під час рекомбінації.
місця з'єднання V-D , D-J і V-J-сегментів можуть зміщуватися
"Помилки" під час рекомбінації.
місця з'єднання V-D , D-J і V-J-сегментів можуть зміщуватися

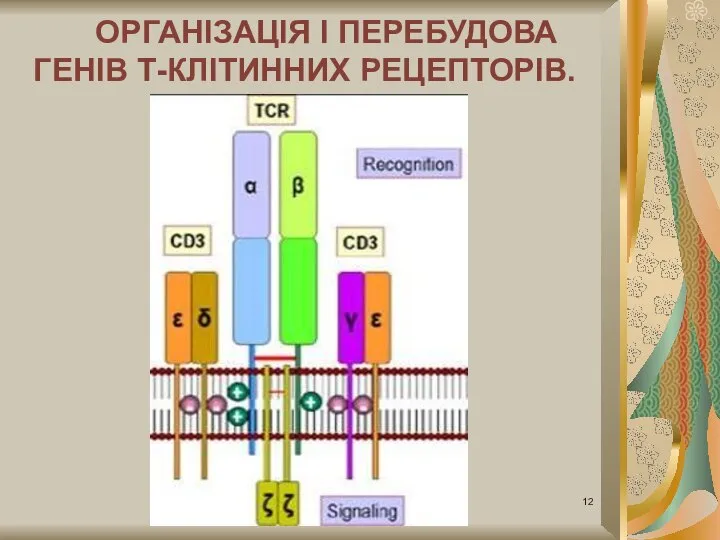
Слайд 12ОРГАНІЗАЦІЯ I ПЕРЕБУДОВА ГЕНІВ Т-КЛІТИННИХ РЕЦЕПТОРІВ.
ОРГАНІЗАЦІЯ I ПЕРЕБУДОВА ГЕНІВ Т-КЛІТИННИХ РЕЦЕПТОРІВ.
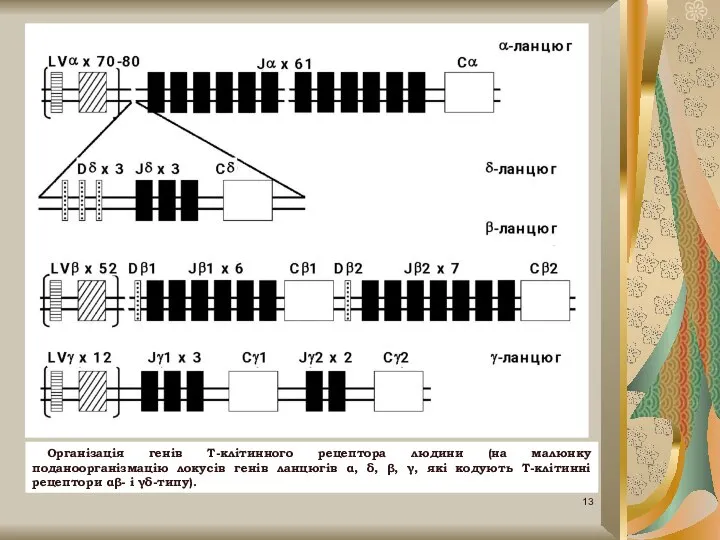
Слайд 13Організація генів Т-клітинного рецептора людини (на малюнку поданоорганізмацію локусів генів ланцюгів α,
Організація генів Т-клітинного рецептора людини (на малюнку поданоорганізмацію локусів генів ланцюгів α,
 Насекомые Крыма
Насекомые Крыма Систематика растений
Систематика растений Изучение механизма наследственности
Изучение механизма наследственности Тест. Коллоквиум №3 (НМА, ЖВС, возбудимые ткани)
Тест. Коллоквиум №3 (НМА, ЖВС, возбудимые ткани) Отдел Покрытосеменные
Отдел Покрытосеменные Многообразие и значение рыб
Многообразие и значение рыб Природные сообщества
Природные сообщества Внутриутробное развитие организма. Развитие после рождения
Внутриутробное развитие организма. Развитие после рождения Презентация на тему Живі організми як особливе середовище існування
Презентация на тему Живі організми як особливе середовище існування  Can dolphin talk
Can dolphin talk О чем рассказал скелет…
О чем рассказал скелет… Зайчишка
Зайчишка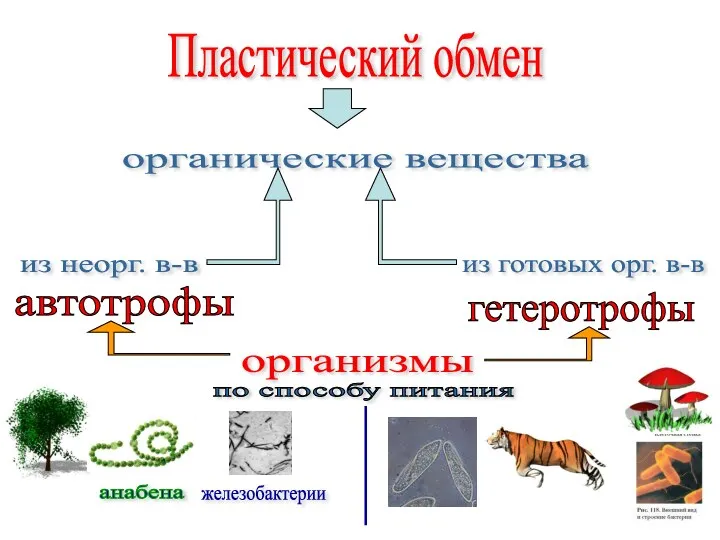 Пластический обмен у автотрофов
Пластический обмен у автотрофов Структура эпидермиса и его функциональное значение
Структура эпидермиса и его функциональное значение Клетка. Возникновение клеточной теории
Клетка. Возникновение клеточной теории Примитивные организмы
Примитивные организмы Опорно-двигательная система. Скелет человека
Опорно-двигательная система. Скелет человека Биоэлементы
Биоэлементы Современные технологии выращивания Окуневых
Современные технологии выращивания Окуневых Энергетический обмен
Энергетический обмен Цікаве про тритонів
Цікаве про тритонів Простейшие организмы. Протисты
Простейшие организмы. Протисты Великий естествоиспытатель Чарльз Дарвин. 5 класс
Великий естествоиспытатель Чарльз Дарвин. 5 класс Высшая Нервная Деятельность
Высшая Нервная Деятельность Презентация на тему Наложение повязок
Презентация на тему Наложение повязок 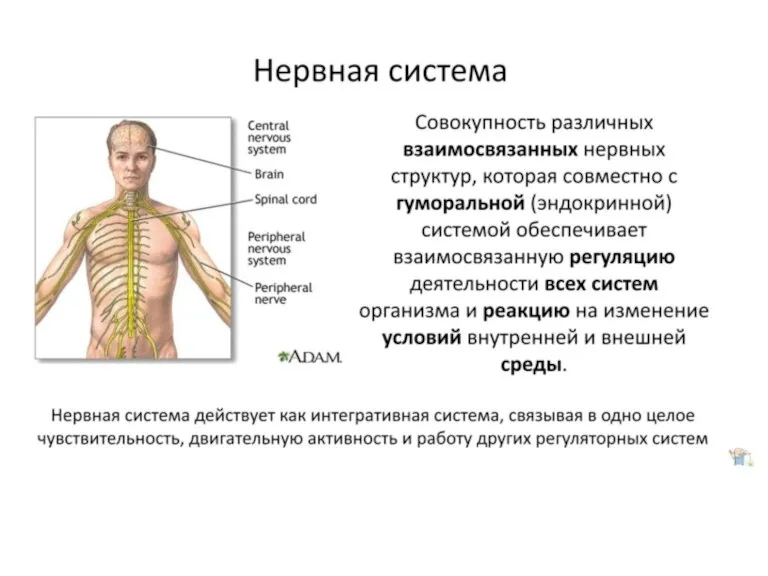 Нервная система человека
Нервная система человека Покрытосеменные растения и их классификация
Покрытосеменные растения и их классификация Сон и здоровье
Сон и здоровье