- Мутации. Номенклатура бактериальных генов

Содержание
- 2. Номенклатура фенотипов His+ означает, что данный бактериальный штамм может расти на среде без гистидина. Имеется в
- 3. Ауксотрофные мутанты, или ауксотрофы – мутантные бактерии, не способные расти и вообще жить, если извне в
- 4. Изоляция условно-летальных ауксотрофных мутантов Аналогичным образом можно изолировать и катаболические, и температуро-чувствительные мутанты.
- 5. Мутации устойчивости приводят к тому, что мутанты способны расти в присутствии вещества, которое для нормальной бактерии
- 6. Важнейшее свойство бактериальных мутаций – их клональность. Если в бактериальной клетке случилась мутация, то все ее
- 7. Типы мутаций Замена, делеция или вставка пары оснований (точечная мутация) Делеция Дупликация тандемных повторов Инверсия Вставка
- 8. Замена пары оснований Может случаться из-за ошибок репликации, радиационного повреждения, химических воздействий. Спаривание гуанина с альтернативной
- 9. Дезаминирование цитозина Происходит спонтанно с достаточно высокой частотой, особенно при повышенных температурах, с образованием урацила.
- 10. Удаление урацила из ДНК, где он совершенно не нужен
- 11. Последствия замены пары оснований Такие мутации фенотипически проявляются нечасто, поскольку с высокой вероятностью происходят в некодирующих
- 12. Делеция или вставка пары оснований Основная причина – наличие гомополимерных участков в составе ДНК. ДНК-полимераза плохо
- 13. Последствия делеции или вставки пары оснований Некодирующие участки генома или гены РНК – аналогично заменам пары
- 14. Делеции Спонтанно происходят достаточно редко. Основная причина – эктопическая рекомбинация, то есть рекомбинация, происходящая не там,
- 15. Дупликации тандемных повторов Происходят одновременно с делециями и по тому же механизму. Однако же именно дупликация
- 16. Инверсии Происходят аналогично делециям, но при наличии ИНВЕРТИРОВАННЫХ повторов. Обозначение инверсий: IN(purB-trpA)3 – инверсия №3, затрагивающая
- 17. Вставки Чаще всего происходят вследствие активности мобильных элементов генома. Об этом мы поговорим позже. Обозначение вставок:
- 18. Вследствие высокой скорости мутагенеза у бактерий, эффект мутаций в их геноме часто сводится на нет при
- 19. Классический пример интергенной супрессии: мутация в одном гене галактозного оперона супрессирует мутацию в другом гене этого
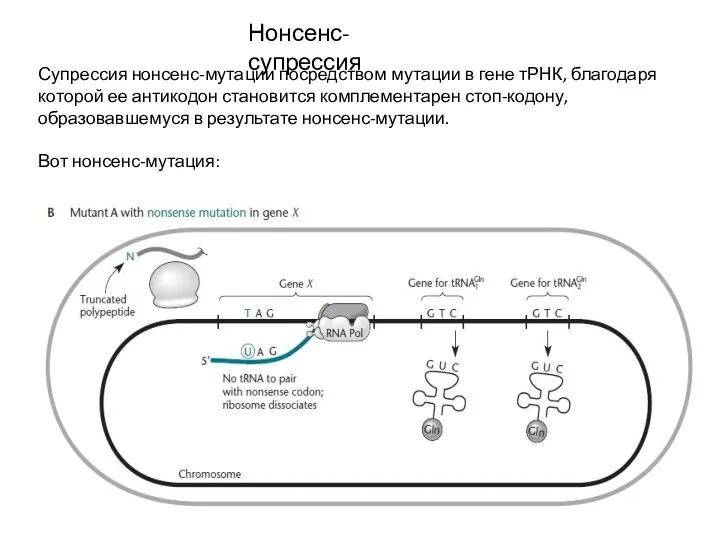
- 20. Нонсенс-супрессия Супрессия нонсенс-мутации посредством мутации в гене тРНК, благодаря которой ее антикодон становится комплементарен стоп-кодону, образовавшемуся
- 21. А вот нонсенс-супрессия: Последовательность белка будет другой, одна аминокислота заменится Супрессорная тРНК будет работать и на
- 22. Анализ бактериальных мутаций Бактерии – удобнейший генетический объект! Поэтому мутационный анализ бактерий до сих пор остается
- 23. Селекция мутантов
- 25. Скачать презентацию

Слайд 3Ауксотрофные мутанты, или ауксотрофы – мутантные бактерии, не способные расти и вообще
Ауксотрофные мутанты, или ауксотрофы – мутантные бактерии, не способные расти и вообще

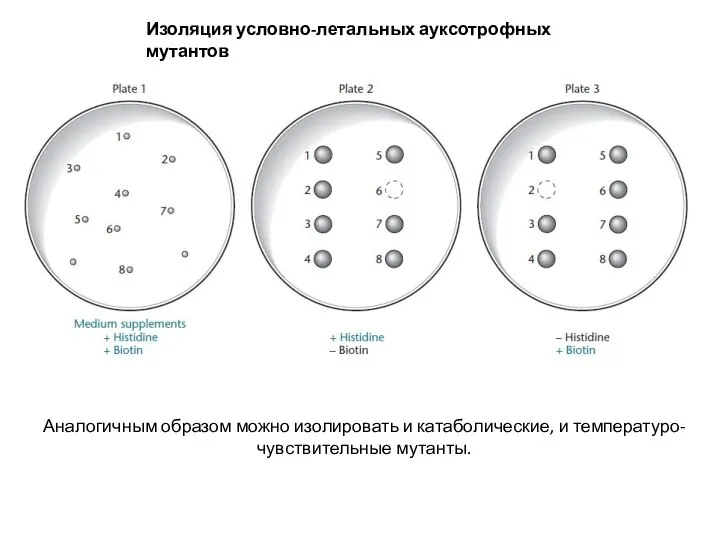
Слайд 4Изоляция условно-летальных ауксотрофных мутантов
Аналогичным образом можно изолировать и катаболические, и температуро-чувствительные мутанты.
Изоляция условно-летальных ауксотрофных мутантов
Аналогичным образом можно изолировать и катаболические, и температуро-чувствительные мутанты.
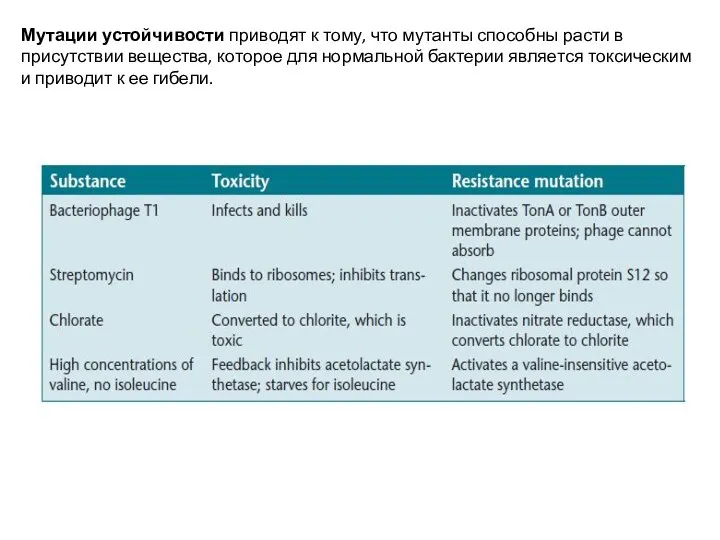
Слайд 5Мутации устойчивости приводят к тому, что мутанты способны расти в присутствии вещества,
Мутации устойчивости приводят к тому, что мутанты способны расти в присутствии вещества,
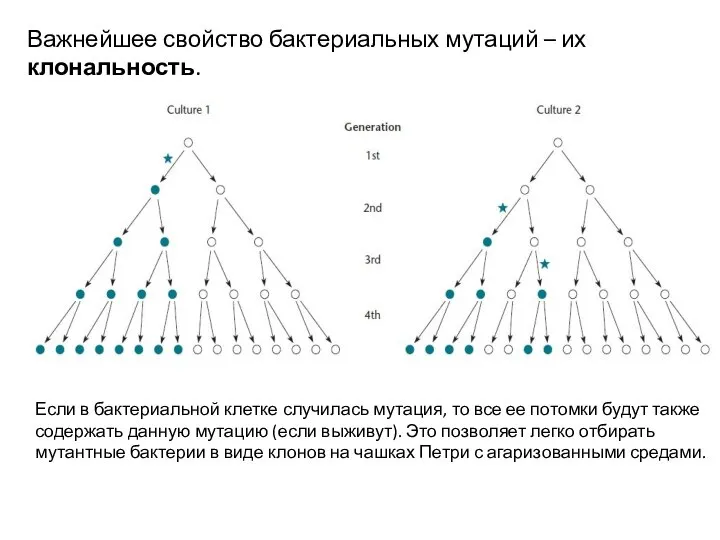
Слайд 6Важнейшее свойство бактериальных мутаций – их клональность.
Если в бактериальной клетке случилась мутация,
Важнейшее свойство бактериальных мутаций – их клональность.
Если в бактериальной клетке случилась мутация,
Слайд 7Типы мутаций
Замена, делеция или вставка пары оснований (точечная мутация)
Делеция
Дупликация тандемных повторов
Инверсия
Вставка
Типы мутаций
Замена, делеция или вставка пары оснований (точечная мутация)
Делеция
Дупликация тандемных повторов
Инверсия
Вставка

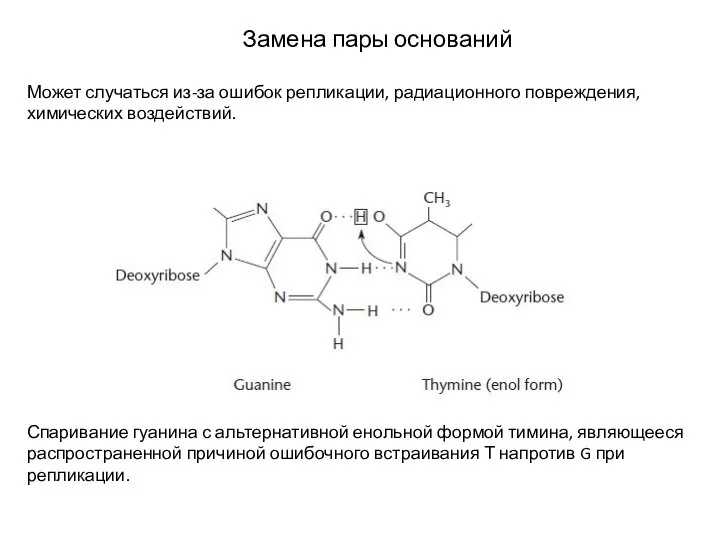
Слайд 8Замена пары оснований
Может случаться из-за ошибок репликации, радиационного повреждения, химических воздействий.
Спаривание
Замена пары оснований
Может случаться из-за ошибок репликации, радиационного повреждения, химических воздействий.
Спаривание
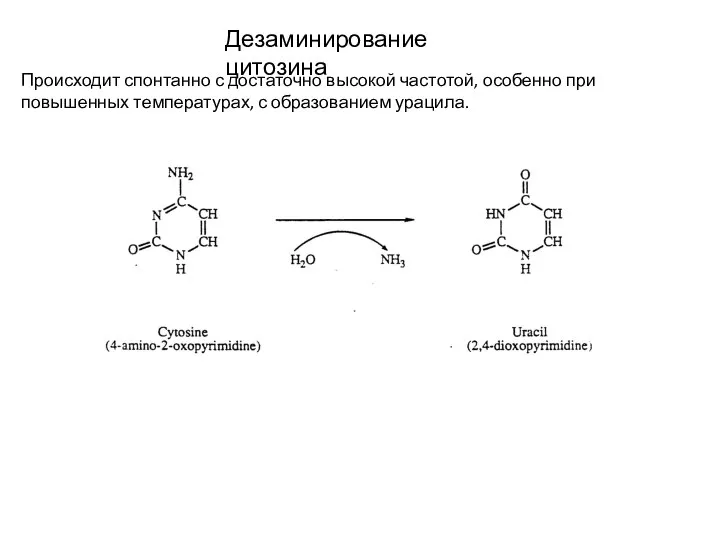
Слайд 9Дезаминирование цитозина
Происходит спонтанно с достаточно высокой частотой, особенно при повышенных температурах, с
Дезаминирование цитозина
Происходит спонтанно с достаточно высокой частотой, особенно при повышенных температурах, с
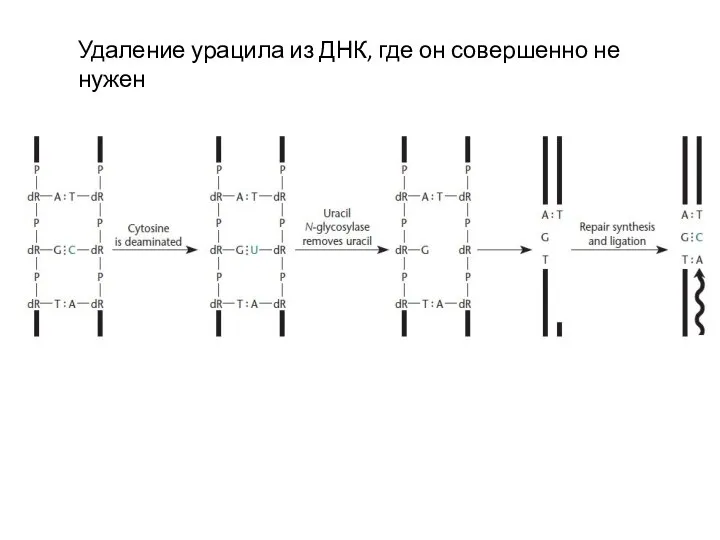
Слайд 10Удаление урацила из ДНК, где он совершенно не нужен
Удаление урацила из ДНК, где он совершенно не нужен
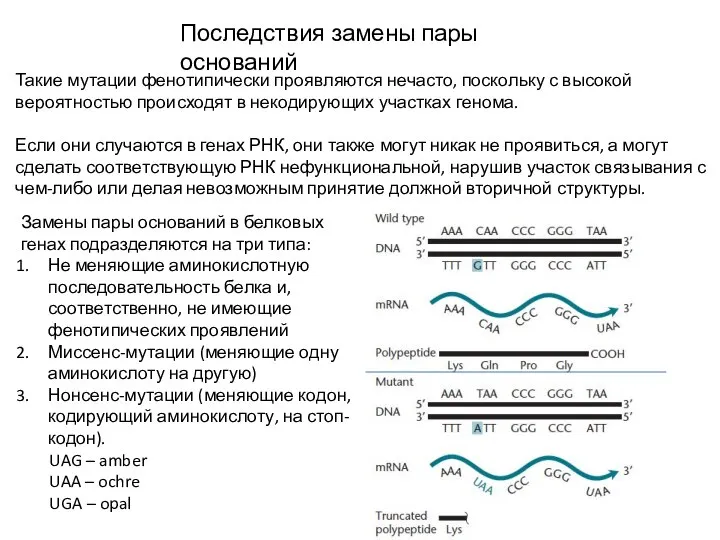
Слайд 11Последствия замены пары оснований
Такие мутации фенотипически проявляются нечасто, поскольку с высокой вероятностью
Последствия замены пары оснований
Такие мутации фенотипически проявляются нечасто, поскольку с высокой вероятностью
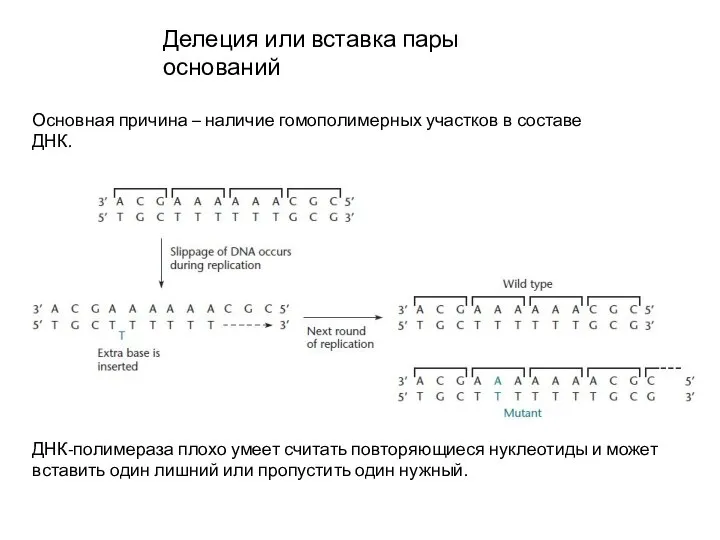
Слайд 12Делеция или вставка пары оснований
Основная причина – наличие гомополимерных участков в составе
Делеция или вставка пары оснований
Основная причина – наличие гомополимерных участков в составе
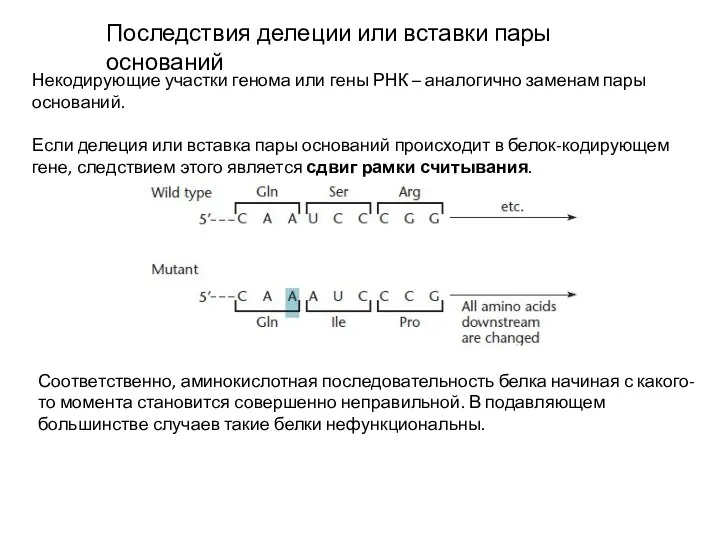
Слайд 13Последствия делеции или вставки пары оснований
Некодирующие участки генома или гены РНК –
Последствия делеции или вставки пары оснований
Некодирующие участки генома или гены РНК –
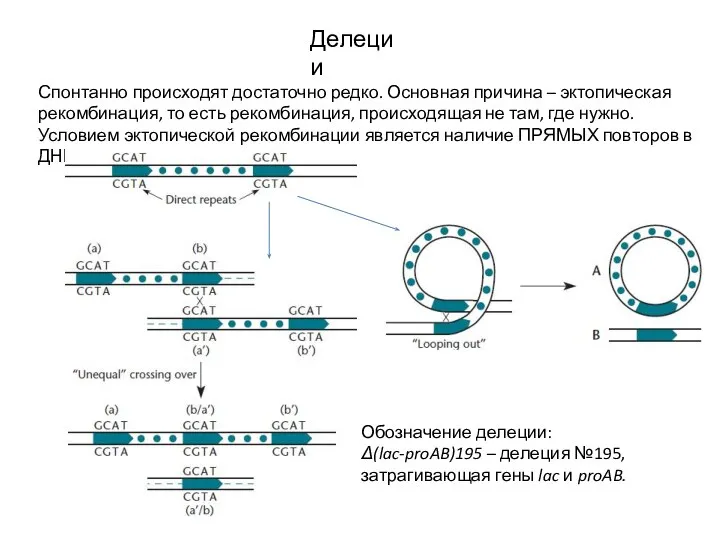
Слайд 14Делеции
Спонтанно происходят достаточно редко. Основная причина – эктопическая рекомбинация, то есть рекомбинация,
Делеции
Спонтанно происходят достаточно редко. Основная причина – эктопическая рекомбинация, то есть рекомбинация,
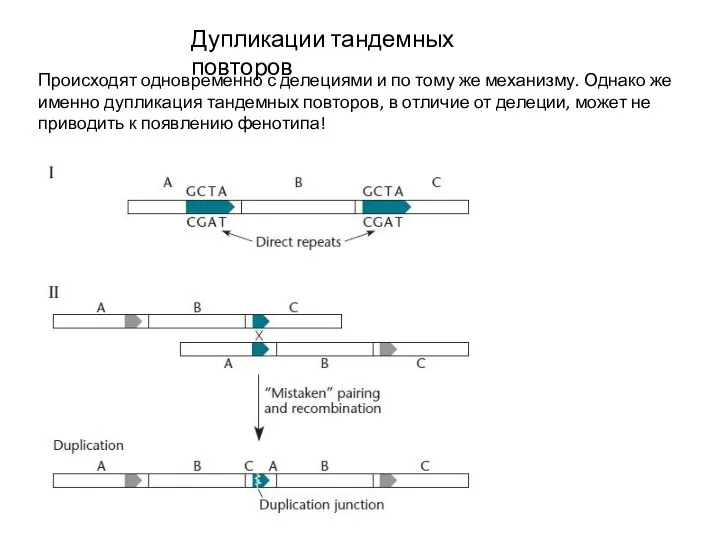
Слайд 15Дупликации тандемных повторов
Происходят одновременно с делециями и по тому же механизму. Однако
Дупликации тандемных повторов
Происходят одновременно с делециями и по тому же механизму. Однако
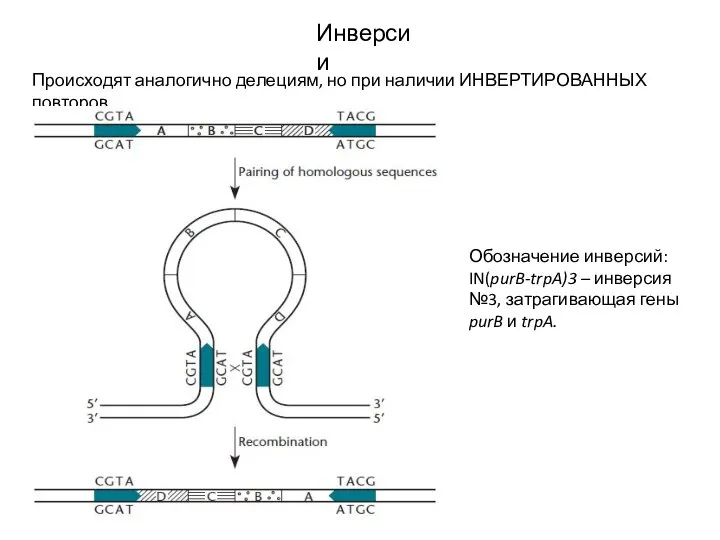
Слайд 16Инверсии
Происходят аналогично делециям, но при наличии ИНВЕРТИРОВАННЫХ повторов.
Обозначение инверсий: IN(purB-trpA)3 – инверсия
Инверсии
Происходят аналогично делециям, но при наличии ИНВЕРТИРОВАННЫХ повторов.
Обозначение инверсий: IN(purB-trpA)3 – инверсия
Слайд 17Вставки
Чаще всего происходят вследствие активности мобильных элементов генома.
Об этом мы поговорим позже.
Обозначение
Вставки
Чаще всего происходят вследствие активности мобильных элементов генома.
Об этом мы поговорим позже.
Обозначение

Слайд 18Вследствие высокой скорости мутагенеза у бактерий, эффект мутаций в их геноме часто
Вследствие высокой скорости мутагенеза у бактерий, эффект мутаций в их геноме часто

Слайд 19Классический пример интергенной супрессии: мутация в одном гене галактозного оперона супрессирует мутацию
Классический пример интергенной супрессии: мутация в одном гене галактозного оперона супрессирует мутацию

Слайд 20Нонсенс-супрессия
Супрессия нонсенс-мутации посредством мутации в гене тРНК, благодаря которой ее антикодон становится
Нонсенс-супрессия
Супрессия нонсенс-мутации посредством мутации в гене тРНК, благодаря которой ее антикодон становится
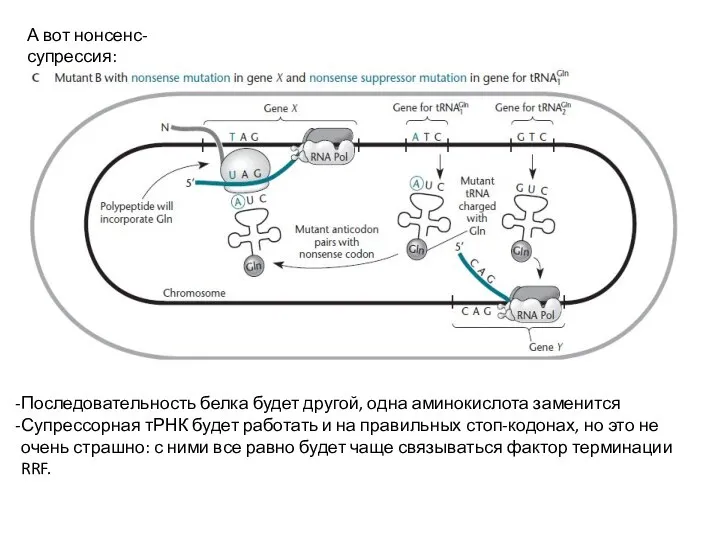
Слайд 21А вот нонсенс-супрессия:
Последовательность белка будет другой, одна аминокислота заменится
Супрессорная тРНК будет работать
А вот нонсенс-супрессия:
Последовательность белка будет другой, одна аминокислота заменится
Супрессорная тРНК будет работать
Слайд 22Анализ бактериальных мутаций
Бактерии – удобнейший генетический объект! Поэтому мутационный анализ бактерий до
Анализ бактериальных мутаций
Бактерии – удобнейший генетический объект! Поэтому мутационный анализ бактерий до

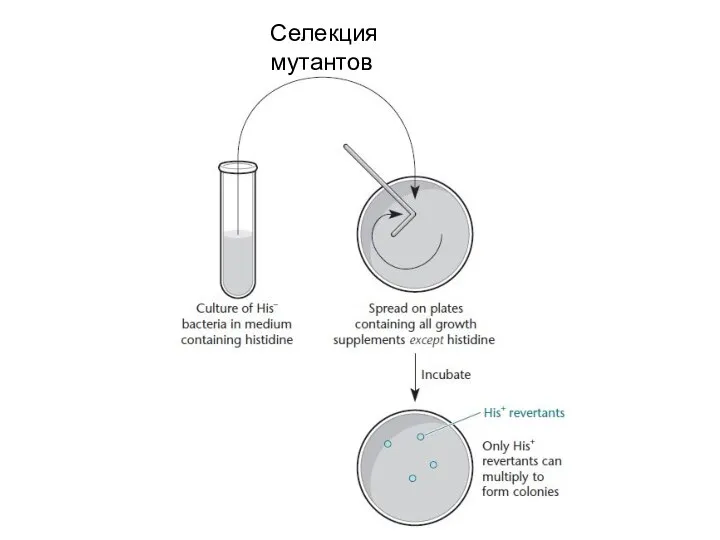
Слайд 23Селекция мутантов
Селекция мутантов
 Презентация на тему Губки. Тупиковая ветвь эволюции
Презентация на тему Губки. Тупиковая ветвь эволюции  Физиология ЦНС
Физиология ЦНС Обмен веществ (метаболизм)
Обмен веществ (метаболизм) Рефлекторная дуга. Коленный рефлекс
Рефлекторная дуга. Коленный рефлекс محاضرة تشريح
محاضرة تشريح Примат горилла
Примат горилла Семейство бобовые (мотыльковые)
Семейство бобовые (мотыльковые) Крестовик. Паукообразные
Крестовик. Паукообразные Значение насекомых в природе
Значение насекомых в природе Земляника нейтрального светового дня. Голландская селекционная компания АBZ Seeds
Земляника нейтрального светового дня. Голландская селекционная компания АBZ Seeds Презентация на тему Культурные растения 6 класс
Презентация на тему Культурные растения 6 класс  Вредители смородины и крыжовника
Вредители смородины и крыжовника Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные
Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные Витаминдер
Витаминдер Микроэволюция. Эволюциялық процес
Микроэволюция. Эволюциялық процес Из какого рода шиншилла?
Из какого рода шиншилла? Спинной мозг
Спинной мозг Выращивание мелиссы лимонной в деревне Хийденсельга
Выращивание мелиссы лимонной в деревне Хийденсельга Презентация на тему Размножение и развитие птиц (7 класс)
Презентация на тему Размножение и развитие птиц (7 класс)  Микология (греч. Микес – гриб) - наука о грибах
Микология (греч. Микес – гриб) - наука о грибах По морям, по волнам. Сюжетная викторина о животных морей и океанов
По морям, по волнам. Сюжетная викторина о животных морей и океанов История создания клеточной теории
История создания клеточной теории Здравствуй, птичья страна!
Здравствуй, птичья страна! Строение и функции кожи
Строение и функции кожи Презентация на тему МИКРООРГАНИЗМЫ-ХУДОЖНИКИ
Презентация на тему МИКРООРГАНИЗМЫ-ХУДОЖНИКИ  Животный мир (фотоальбом)
Животный мир (фотоальбом) Презентация на тему СЕЛЕКЦИЯ РАСТЕНИЙ
Презентация на тему СЕЛЕКЦИЯ РАСТЕНИЙ  Поверхностные явления. Образование мицелл. (Лекция 7)
Поверхностные явления. Образование мицелл. (Лекция 7)