- Нуклеиновые кислоты. Лекция №3

Содержание
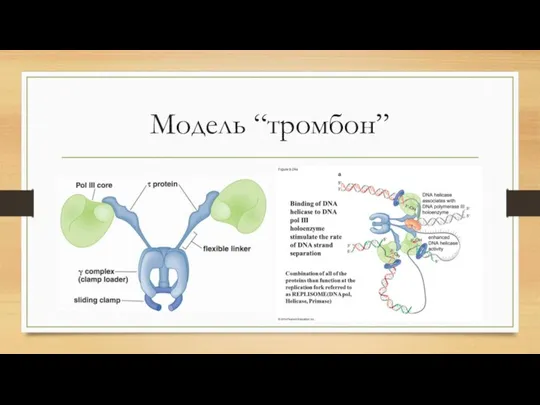
- 2. Модель “тромбон”
- 3. Топология ДНК Топология - раздел математики, имеющий своим назначением выяснение и исследование, в рамках математики, идеи
- 4. Математическое описание сверхспирализации ДНК В 1969 году Уайтом была предложена формула, которая связала порядок зацепления и
- 5. Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в результате которого ось двойной
- 6. Решение проблемы сверхспирализации ДНК Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию ДНК. Топоизомеразы способны релаксировать
- 7. Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы без затрат АТФ. Механизм
- 8. Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в виде димеров. Они осуществляют
- 9. Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины Топоизомеразы играют важную роль в процессах роста
- 10. Транскрипция РНК Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех
- 11. Свойства генетического кода Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность
- 12. Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует
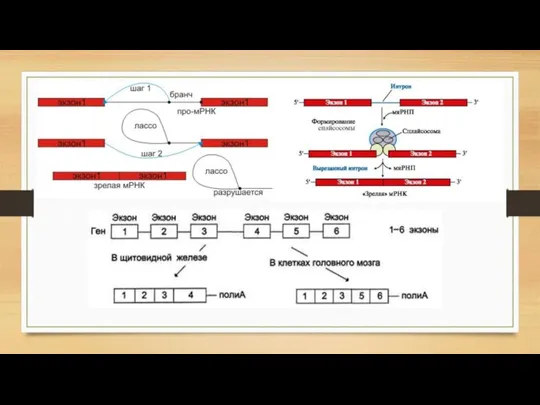
- 13. Процессинг РНК Кэпирование Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в
- 14. 2) Полиаденилирование Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК (пре-мРНК). Иными словами,
- 16. 4) Редактирование Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется путём химической модификации
- 17. Общий вывод о созревании мРНК
- 18. Список использованной литературы 1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и др. 2) «Клетки»,
- 20. Скачать презентацию
Слайд 3Топология ДНК
Топология - раздел математики, имеющий своим назначением выяснение и исследование, в
Топология ДНК
Топология - раздел математики, имеющий своим назначением выяснение и исследование, в

Слайд 4Математическое описание сверхспирализации ДНК
В 1969 году Уайтом была предложена формула, которая связала
Математическое описание сверхспирализации ДНК
В 1969 году Уайтом была предложена формула, которая связала

Слайд 5Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в
Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в

Слайд 6Решение проблемы сверхспирализации ДНК
Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию
Решение проблемы сверхспирализации ДНК
Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию

Слайд 7Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы
Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы

Слайд 8Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в
Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в

Слайд 9Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины
Топоизомеразы играют важную роль
Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины
Топоизомеразы играют важную роль

Слайд 10Транскрипция РНК
Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы,
Транскрипция РНК
Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы,

Слайд 11Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет,
Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет,

Слайд 12Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA

Слайд 13Процессинг РНК
Кэпирование
Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные
Процессинг РНК
Кэпирование
Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные

Слайд 142) Полиаденилирование
Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК
2) Полиаденилирование
Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК

Слайд 164) Редактирование
Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется
4) Редактирование
Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется

Слайд 17Общий вывод о созревании мРНК
Общий вывод о созревании мРНК

Слайд 18Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и
Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и

 Анализаторы. Строение и общие закономерности развития сенсорных систем
Анализаторы. Строение и общие закономерности развития сенсорных систем Брюхоногие моллюски или гастроподы
Брюхоногие моллюски или гастроподы Овощные культуры
Овощные культуры Подводный мир (слйды)
Подводный мир (слйды) Витамины A,B,C,D
Витамины A,B,C,D Морфология микроорганизмов. Методы окраски
Морфология микроорганизмов. Методы окраски Основы биомеханического контроля
Основы биомеханического контроля Зоотенический учёт. Методы разведения сельскохозяйственных животных
Зоотенический учёт. Методы разведения сельскохозяйственных животных Ротовые аппараты насекомых
Ротовые аппараты насекомых Класс земноводные
Класс земноводные Опорно - двигательная система. Урок № 2.3. Скелет конечностей
Опорно - двигательная система. Урок № 2.3. Скелет конечностей Основы анатомии, физиологии и биофизики
Основы анатомии, физиологии и биофизики Палеозойская эра
Палеозойская эра Презентация на тему Состав почвы
Презентация на тему Состав почвы  Питание и здоровье
Питание и здоровье Когда были открыты витамины
Когда были открыты витамины Апельсин китайский
Апельсин китайский Повторение и обобщение знаний по теме: Класс Насекомые. 7 класс
Повторение и обобщение знаний по теме: Класс Насекомые. 7 класс Выращивание сеянца дуба из желудя разными способами в комнатных условиях
Выращивание сеянца дуба из желудя разными способами в комнатных условиях Гигиена питания
Гигиена питания Выращивание цветочной рассады
Выращивание цветочной рассады Витамины. Тест
Витамины. Тест Пищеварительная система
Пищеварительная система Декоративное растениеводство
Декоративное растениеводство Видоизменения корней
Видоизменения корней Zheludochno-kishechny_trakt_Prezentatsia_3
Zheludochno-kishechny_trakt_Prezentatsia_3 Пищеварение в желудке. Цель: исследовать действие желудочного сока на белок в зависимости от разных условий
Пищеварение в желудке. Цель: исследовать действие желудочного сока на белок в зависимости от разных условий Презентация на тему Дыхание и здоровье человека
Презентация на тему Дыхание и здоровье человека