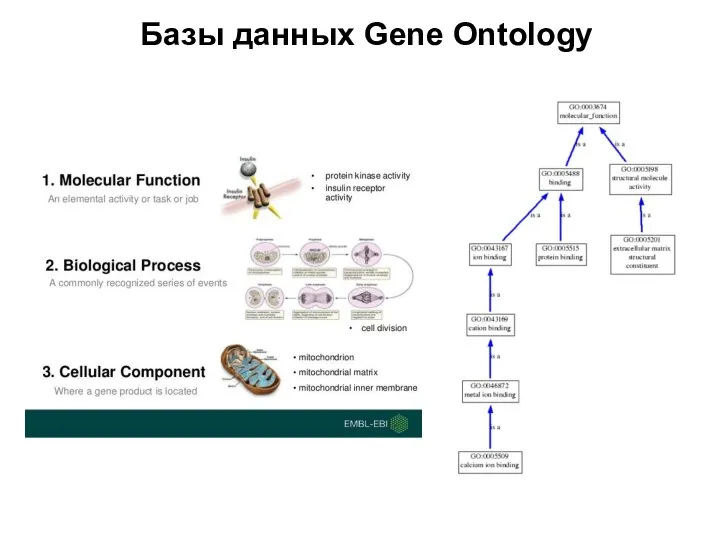
- Базы данных Gene Ontology

Содержание
- 2. Базы данных Gene Ontology
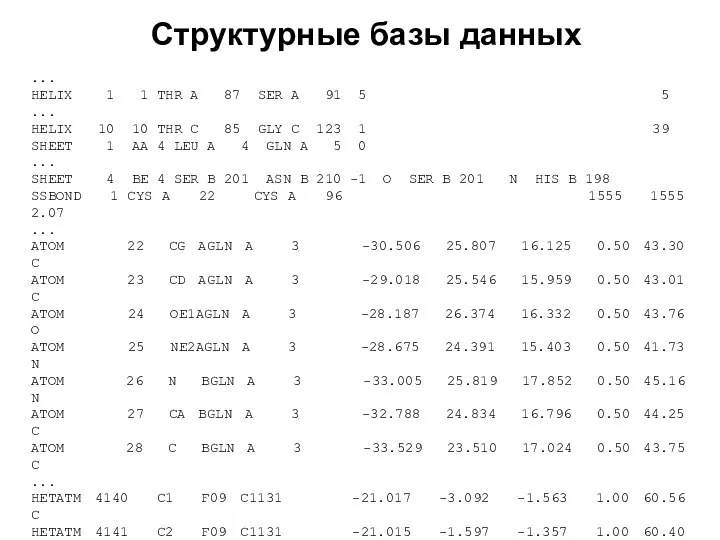
- 3. Структурные базы данных PDB (Brookhaven Protein DataBank) – коллекция экспериментально определенных 3D-структур биологических макромолекул. Раньше содержала
- 4. Структурные базы данных
- 5. Структурные базы данных
- 6. Структурные базы данных ... REMARK 465 REMARK 465 MISSING RESIDUES REMARK 465 THE FOLLOWING RESIDUES WERE
- 7. Структурные базы данных ... HELIX 1 1 THR A 87 SER A 91 5 5 ...
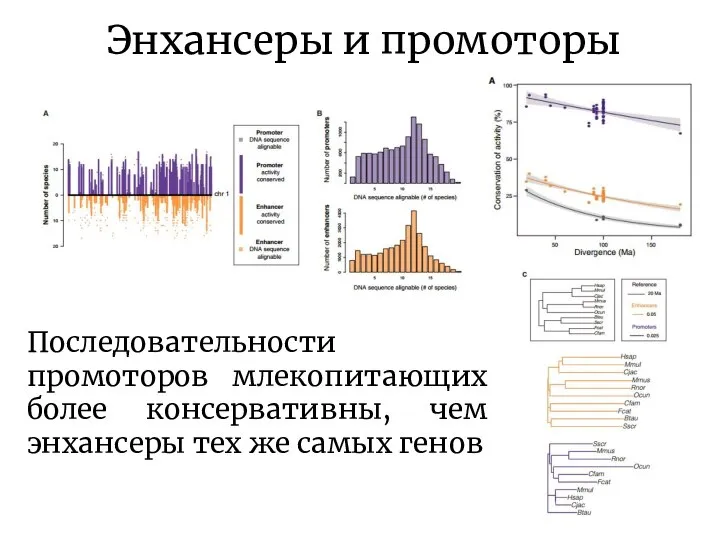
- 8. Энхансеры и промоторы Последовательности промоторов млекопитающих более консервативны, чем энхансеры тех же самых генов
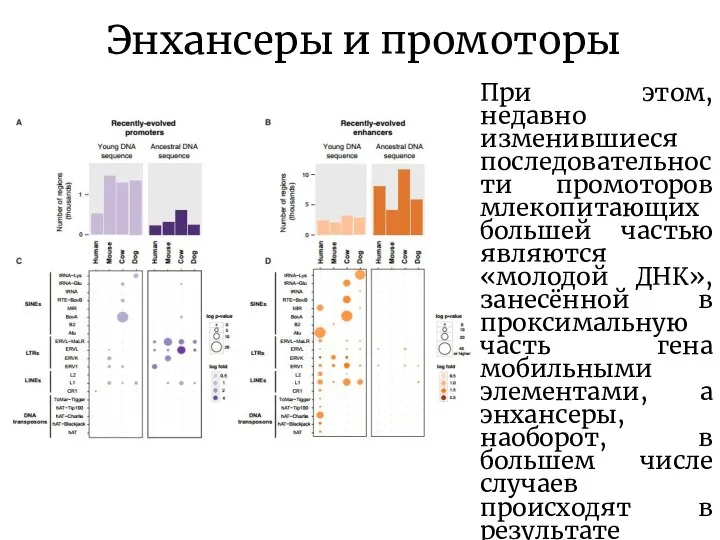
- 9. Энхансеры и промоторы При этом, недавно изменившиеся последовательности промоторов млекопитающих большей частью являются «молодой ДНК», занесённой
- 10. Возникновение энхансеров de novo Большинство энхансеров млекопитающих апоморфны
- 11. Промоторы млекопитающих
- 14. Гены, участвующих в развитии, ответе на стресс, регуляции клеточных процессов, обладают большим числом регуляторных элементов, чем
- 15. Эволюция системы транскрипционного фактора и гена-мишени
- 16. Эволюция системы транскрипционного фактора и гена-мишени
- 17. Глубокая гомология Формирование и дифференциация многих структур, таких как глаза, конечности и сердце, управляется одним и
- 18. Глубокая гомология
- 19. Модульность цис-регуляторных элементов Отдельная черта toolkit-генов — большие и сложные модульные цис-регуляторные регионы.
- 20. Модульность цис-регуляторных элементов Наличие большого числа цис-регуляторных элементов у toolkit-генов открывает несколько важных для эволюции формы
- 21. Большие генные сети
- 22. Регуляция 3’ и 5’ UTR
- 24. Скачать презентацию
Слайд 3Структурные базы данных
PDB (Brookhaven Protein DataBank) – коллекция экспериментально определенных 3D-структур биологических макромолекул. Раньше
Структурные базы данных
PDB (Brookhaven Protein DataBank) – коллекция экспериментально определенных 3D-структур биологических макромолекул. Раньше

Слайд 4Структурные базы данных
Структурные базы данных

Слайд 5Структурные базы данных
Структурные базы данных

Слайд 6Структурные базы данных
...
REMARK 465
REMARK 465 MISSING RESIDUES
REMARK 465 THE FOLLOWING
Структурные базы данных
...
REMARK 465
REMARK 465 MISSING RESIDUES
REMARK 465 THE FOLLOWING

Слайд 7Структурные базы данных
...
HELIX 1 1 THR A 87 SER A 91 5
Структурные базы данных
...
HELIX 1 1 THR A 87 SER A 91 5
Слайд 8Энхансеры и промоторы
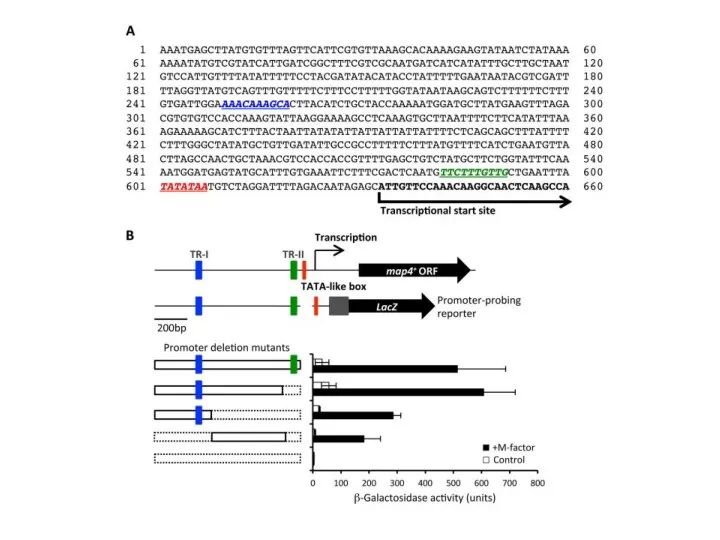
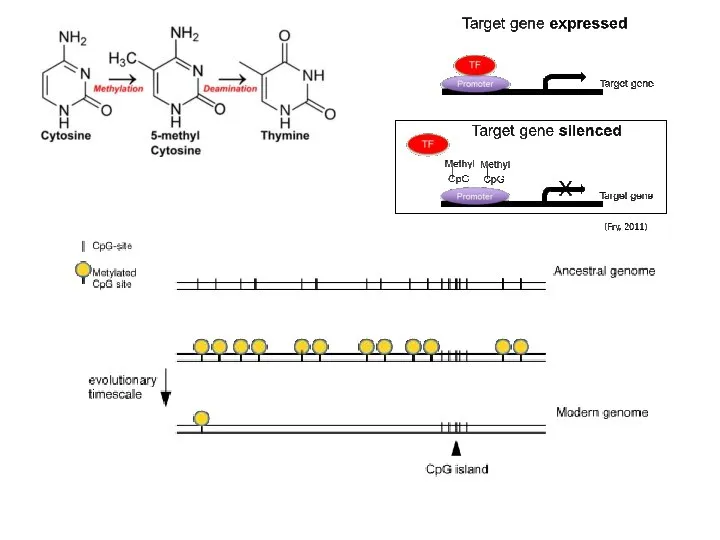
Последовательности промоторов млекопитающих более консервативны, чем энхансеры тех же самых
Энхансеры и промоторы
Последовательности промоторов млекопитающих более консервативны, чем энхансеры тех же самых
Слайд 9Энхансеры и промоторы
При этом, недавно изменившиеся последовательности промоторов млекопитающих большей частью являются
Энхансеры и промоторы
При этом, недавно изменившиеся последовательности промоторов млекопитающих большей частью являются
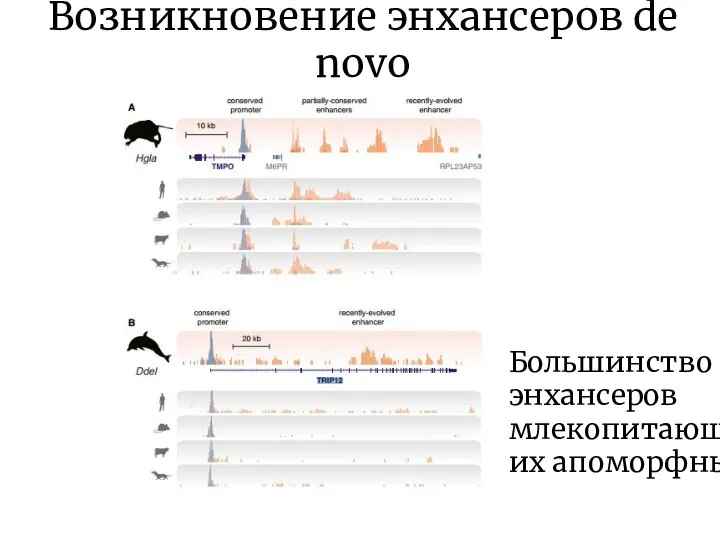
Слайд 10Возникновение энхансеров de novo
Большинство энхансеров млекопитающих апоморфны
Возникновение энхансеров de novo
Большинство энхансеров млекопитающих апоморфны
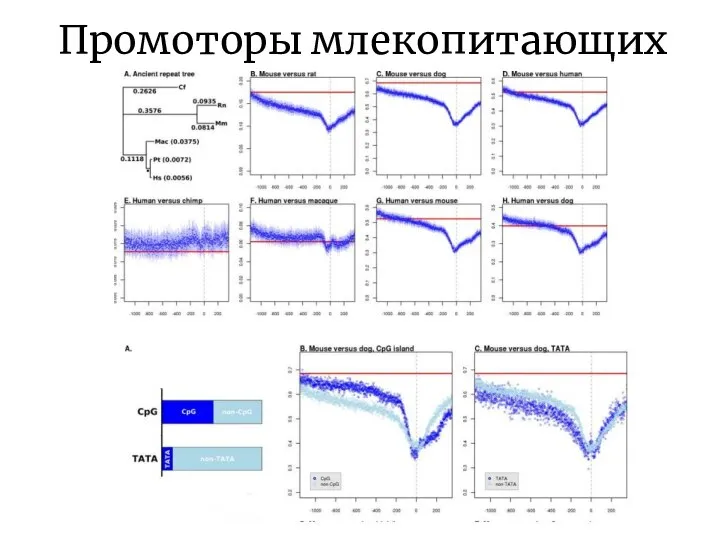
Слайд 11Промоторы млекопитающих
Промоторы млекопитающих
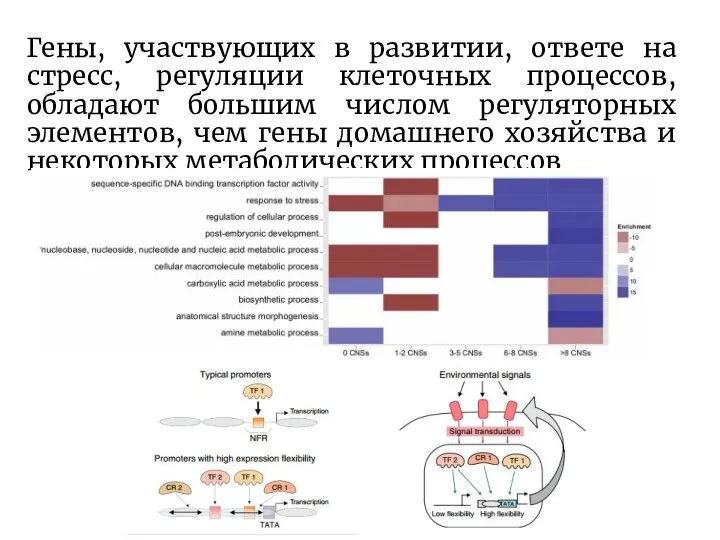
Слайд 14Гены, участвующих в развитии, ответе на стресс, регуляции клеточных процессов, обладают большим
Гены, участвующих в развитии, ответе на стресс, регуляции клеточных процессов, обладают большим
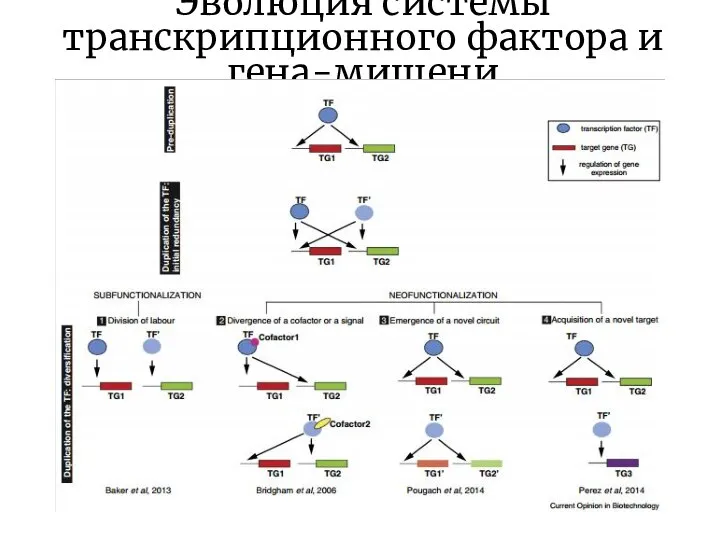
Слайд 15Эволюция системы транскрипционного фактора и гена-мишени
Эволюция системы транскрипционного фактора и гена-мишени
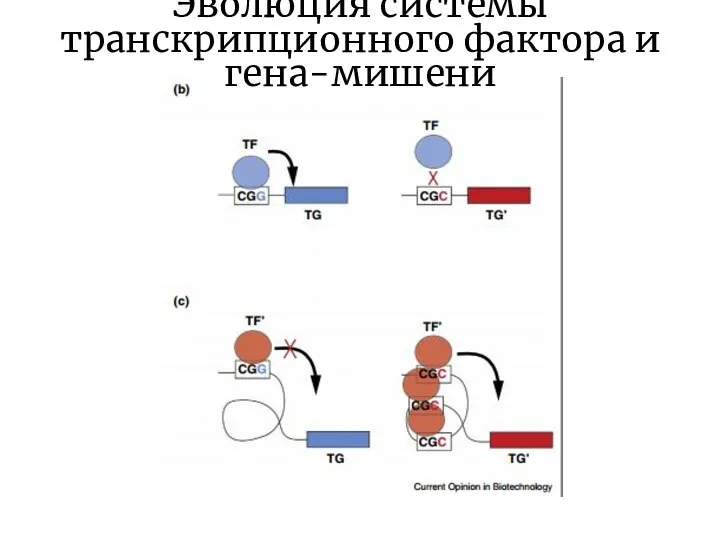
Слайд 16Эволюция системы транскрипционного фактора и гена-мишени
Эволюция системы транскрипционного фактора и гена-мишени
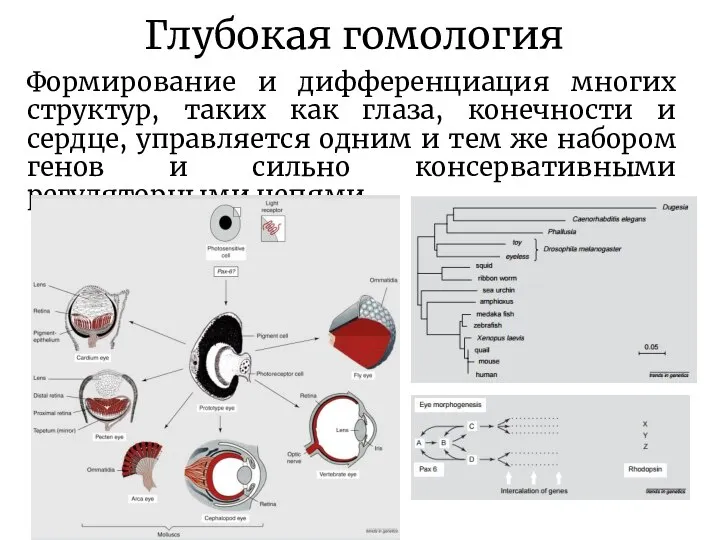
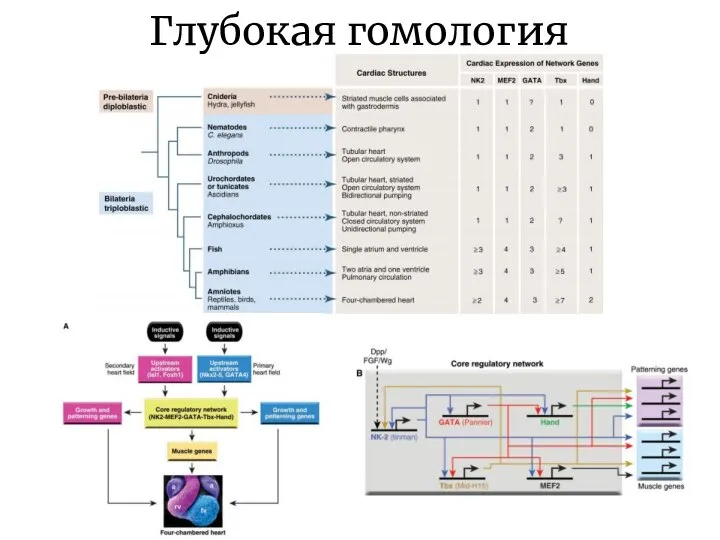
Слайд 17Глубокая гомология
Формирование и дифференциация многих структур, таких как глаза, конечности и сердце,
Глубокая гомология
Формирование и дифференциация многих структур, таких как глаза, конечности и сердце,
Слайд 18Глубокая гомология
Глубокая гомология
Слайд 19Модульность цис-регуляторных элементов
Отдельная черта toolkit-генов — большие и сложные модульные цис-регуляторные регионы.
Модульность цис-регуляторных элементов
Отдельная черта toolkit-генов — большие и сложные модульные цис-регуляторные регионы.

Слайд 20Модульность цис-регуляторных элементов
Наличие большого числа цис-регуляторных элементов у toolkit-генов открывает несколько важных
Модульность цис-регуляторных элементов
Наличие большого числа цис-регуляторных элементов у toolkit-генов открывает несколько важных

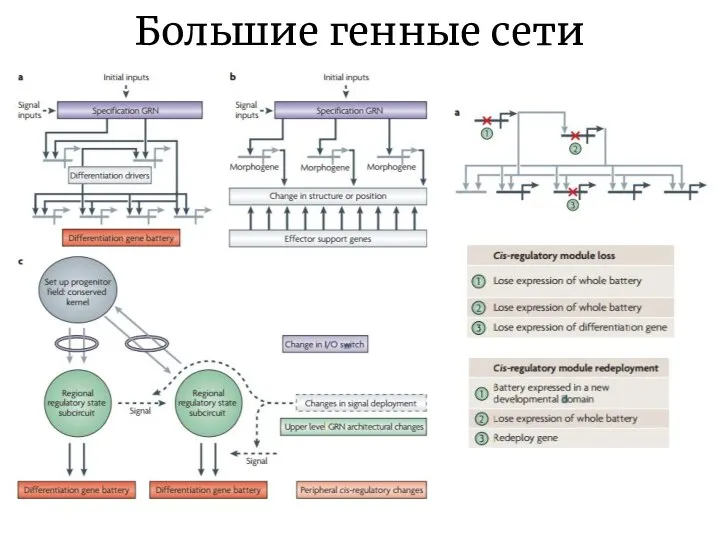
Слайд 21Большие генные сети
Большие генные сети
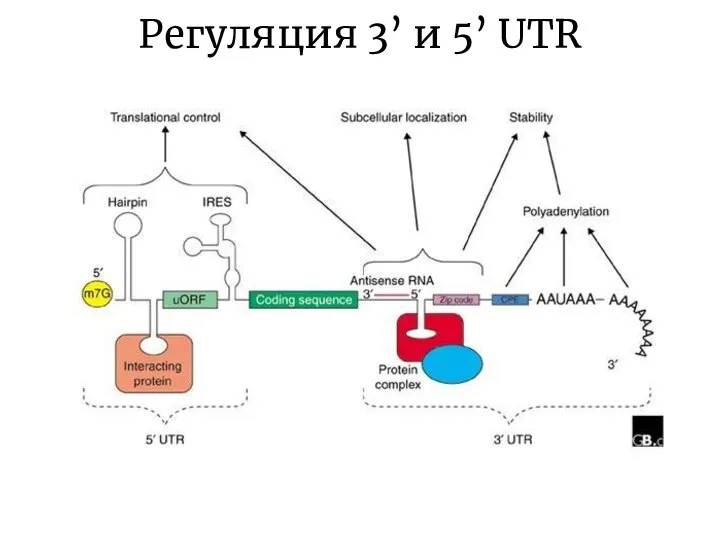
Слайд 22Регуляция 3’ и 5’ UTR
Регуляция 3’ и 5’ UTR
 Поджелудочная железа
Поджелудочная железа Иллюстрированный словарь народных названий растений Урала
Иллюстрированный словарь народных названий растений Урала Строение и физиологические функции кожного покрова пушных зверей
Строение и физиологические функции кожного покрова пушных зверей Опорно-двигательная система
Опорно-двигательная система Muskulýarny sistema
Muskulýarny sistema Парящие над облаками
Парящие над облаками Обмен липидов. Анаболизм липидов
Обмен липидов. Анаболизм липидов Энергетический обмен
Энергетический обмен ЭС 1ДЗ БАЗА
ЭС 1ДЗ БАЗА Метаболизм. Энергообмен
Метаболизм. Энергообмен Хвощи, плауны и папоротники
Хвощи, плауны и папоротники Уровни организации живой материи
Уровни организации живой материи Функции спинного мозга. Рефлекторная функция
Функции спинного мозга. Рефлекторная функция Белок - основа жизни
Белок - основа жизни Строение и функции зубов
Строение и функции зубов Органы кровообращения
Органы кровообращения Медведь
Медведь Метофазный метод учета хромосомных аберраций
Метофазный метод учета хромосомных аберраций 9. Транспорт веществ — в журнал
9. Транспорт веществ — в журнал Метаболізм ліпідів. Метаболізм білків. Стероїди. (Лекція 2)
Метаболізм ліпідів. Метаболізм білків. Стероїди. (Лекція 2) Презентация на тему Формы размножения организмов
Презентация на тему Формы размножения организмов  Ара. Ара пайдалану
Ара. Ара пайдалану Ә.Атнабаевның 90 еллык юбилеена багышлана
Ә.Атнабаевның 90 еллык юбилеена багышлана Особенности обмена отдельных аминокислот
Особенности обмена отдельных аминокислот Витамин H
Витамин H Строение ушной раковины
Строение ушной раковины Происхождение и развитие человека
Происхождение и развитие человека Гормоны мозгового слоя надпочечников
Гормоны мозгового слоя надпочечников