- Пример необратимого ингибирования – ЛВ аспирин

Содержание
- 2. Обратимое I связывается с E нековалентными связями → [IE] легко распадается, активность E при этом восстанавливается
- 3. Конкурентное (изостерическое) I - структурный аналог S I связывается с активным центром E → между I
- 4. Пример конкурентного ингибирования – торможение сукцинатдегидрогеназы малоновой кислотой
- 5. Для устранения действия конкурентного I необходимо увеличить концентрацию S или удалить I
- 6. Конкурентное ингибирование (1 – без I, 2 – с I) графически выражается с помощью кривой Михаэлиса
- 7. Т.о., в присутствии конкурентного I Vmax реакции не меняется, а Кm увеличивается → сродство E к
- 8. На принципе конкурентного ингибирования основано действие многих ЛВ, например, группа ацетилхолинэстеразных препаратов, являющихся конкурентными I АХЭ
- 9. Присоединение конкурентного I прозерина в акт.ц. АХЭ
- 10. Необратимо действуют фосфоорганические препараты: армин, нибуфин, хлорофос, зарин, зоман, фосфорилируя каталитический участок АХЭ
- 11. Сульфаниламид – структурный аналог парааминобензойной кислоты
- 12. Фолиевая кислота образуется в клетках бактерий, если они получают п-аминобензойную кислоту. Сульфаниламиды ингибируют Е, у которых
- 13. Эти Е могут использовать в качестве S сульфаниламиды → синтезируется не фолиевая кислота, а ее аналог,
- 14. → в бактериальных клетках возникает недостаток фолиевой кислоты, нарушаются все реакции, в которых она участвует, и
- 15. Неконкурентное (аллостерическое) Это такое ингибирование, при котором I взаимодействует с Е не в активном центре (А),
- 17. Неконкурентное ингибирование (1 – без I, 2 – с I) графически выражается с помощью кривой Михаэлиса
- 18. Т.о. под действием неконкурентного I Vmax уменьшается, а Кm не изменяется → сродство E к S
- 19. В роли регуляторов V ферментативных реакций наиболее часто выступают: гормоны, медиаторы, ионы металлов, коферменты, различные метаболиты
- 21. Активаторы ферментов 1. Ионы K+, Na+, Mg2+, Mn2+, Co2+, Zn2+, Fe2+, Cl-, SO42-, PO43- и др.
- 22. 2. Специфические агенты (н-р, HCl) и другие ферменты Регуляция сводится к превращению проферментов (неактивных предшественников Е)
- 23. Н-р, пепсиноген превращается в пепсин в результате ограниченного протеолиза: под действием HCl отщепляется пептид, который затрудняет
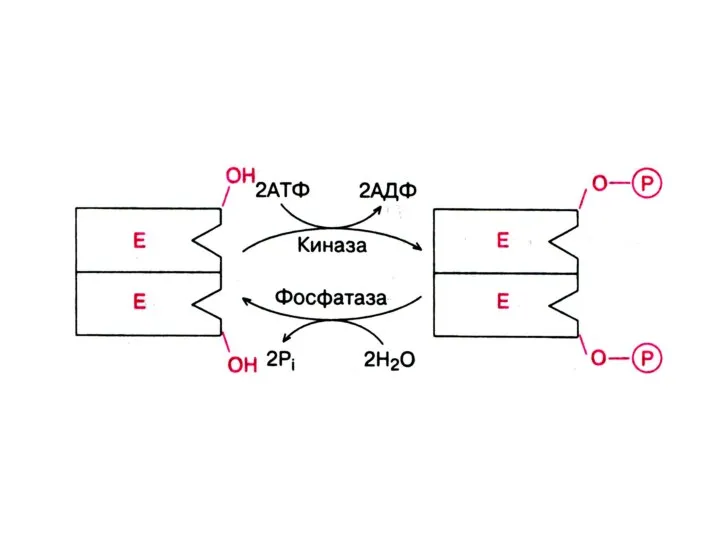
- 24. Активность некоторых Е может регулироваться с помощью химической модификации, н-р, путем фосфорилирования-дефорилирования:
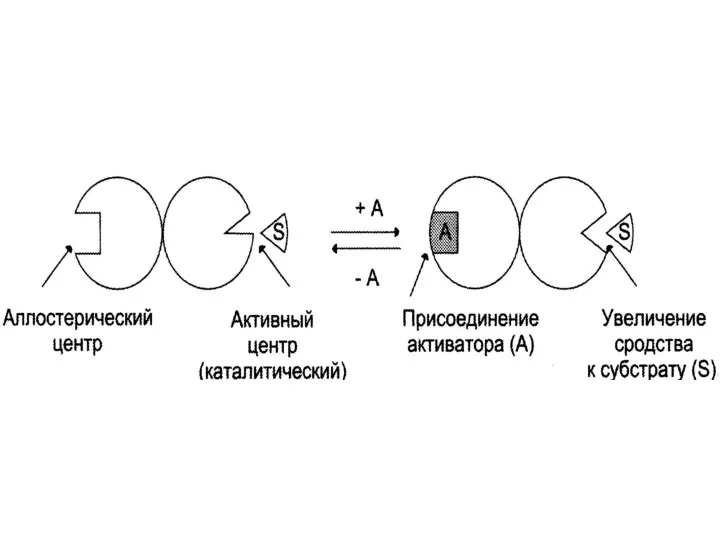
- 26. 3. Аллостерические активаторы Связываются с R, в результате чего конформация E изменяется т.о., что она оптимально
- 28. 4. Активаторы, способствующие объединению неактивных субъединиц E в активный надмолекулярный комплекс, имеющий четвертичную структуру
- 29. Изоферменты Это различные молекулярные формы одного и того же фермента, катализирующие одну и ту же реакцию,
- 30. Например, лактатдегидрогеназа (ЛДГ) имеет четвертичную структуру, содержит 2 типа субъединиц М и H (от heart -
- 31. ЛДГ1 – НННН (Н4) ЛДГ2 – НННМ (Н3М) ЛДГ3 – ННММ (Н2М2) ЛДГ4 – НМММ (НМ3)
- 33. ЛДГ на электрофореграмме в различных органах
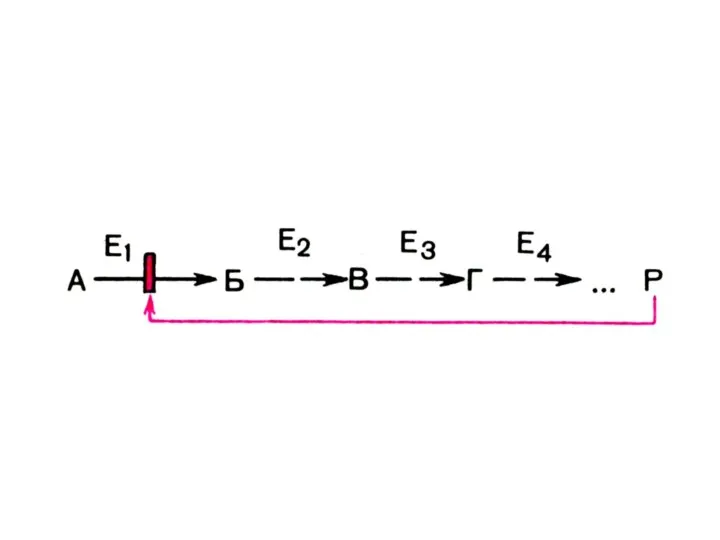
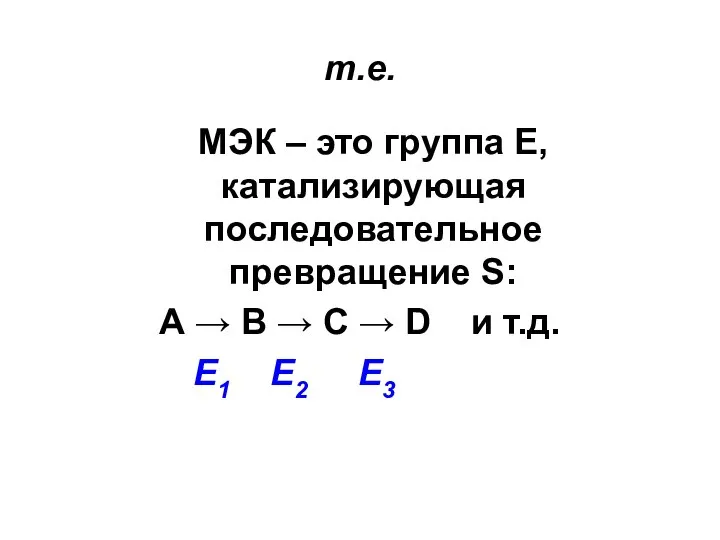
- 34. Мультиэнзимные комплексы (МЭК) - надмолекулярные ферментативные системы, состоящие из различных Е, катализирующих последовательные этапы одного метаболического
- 35. т.е. МЭК – это группа Е, катализирующая последовательное превращение S: А → В → С →
- 36. Существует несколько видов МЭК, в основе организации которых лежит единство: 1 – функциональное 2 – структурно-функциональное
- 37. 1 Отдельные Е объединены в полиферментную систему. Н-р, гликолиз: S1 → P1(S2) → P2 (S3) →...
- 38. 2 Е образуют структурные системы с определенной функцией при помощи Е-Е взаимодействий. Н-р, полиферментный комплекс –
- 39. Пируватдегидрогеназный комплекс
- 40. синтетаза высших жирных кислот, состоящая из 7 структурно связанных Е, в целом выполняющих общую функцию –
- 41. 3 Представляет комбинацию обоих типов организации. Н-р, цикл Кребса, в котором часть Е объединена в α-кетоглутаратдегидрогеназный
- 42. Биологическое значение Образуется «молекулярный конвейер», благодаря которому: значительно сокращается расстояние, на которое переносятся субстраты; Е работают
- 43. Медицинская энзимология Энзимопатология Энзимодиагностика Энзимотерапия
- 44. Энзимопатология изучает наследственные или приобретенные дефекты ферментных систем – энзимопатии. Различают энзимопатии: 1 – первичные (наследственные)
- 45. Н-р, 1: Фенилкетонурия – заболевание, при котором отсутствует Е гидроксилаза, превращающая фенилаланин в тирозин. В результате
- 46. Энзимодиагностика заключается в постановке диагноза заболевания на основе определения активности Е в биологических жидкостях
- 47. Например: При инфаркте миокарда увеличивается содержание ферментов ЛДГ1 и ЛДГ2, аспартатаминотрансферазы. При вирусном гепатите увеличивается содержание
- 48. Энзимотерапия – использование ферментов в качестве лекарственных средств. Имеет много ограничений вследствие высокой иммуногенности ферментов.
- 49. Имеет следующие направления: Заместительная терапия – использование ферментов в качестве лечебных препаратов в случае их недостаточности
- 50. Использование в качестве дополнительных терапевтических средств (н-р, различные гидролитические ферменты для ускорения заживления ран – пепсин,
- 51. Трудности в использовании ферментов: нестабильность антигенные свойства практически невозможность доставки к клеткам-мишеням
- 52. Для увеличения стабильности Е их связывают с различными инертными носителями (целлюлоза, крахмал). Для снижения антигенных свойств
- 53. Для направленного действия Е на мишень, на поверхности микрокапсулы прикрепляют векторную молекулу антитела, которое взаимодействует только
- 54. Ферменты широко используются для определения содержания различных веществ в биологических жидкостях Н-р, с помощью иммуноферментного анализа
- 56. Скачать презентацию
![Обратимое I связывается с E нековалентными связями → [IE] легко распадается, активность E при этом восстанавливается](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1015099/slide-1.jpg)
Слайд 3
Конкурентное (изостерическое)
I - структурный аналог S
I связывается с активным центром E
Конкурентное (изостерическое)
I - структурный аналог S
I связывается с активным центром E

Слайд 4Пример конкурентного ингибирования – торможение сукцинатдегидрогеназы малоновой кислотой
Пример конкурентного ингибирования – торможение сукцинатдегидрогеназы малоновой кислотой
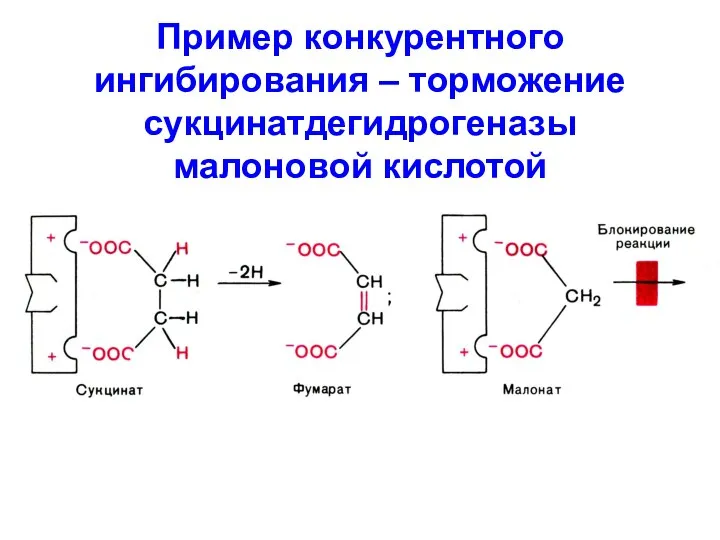
Слайд 5 Для устранения действия конкурентного I необходимо увеличить концентрацию S или удалить I
Для устранения действия конкурентного I необходимо увеличить концентрацию S или удалить I

Слайд 6Конкурентное ингибирование (1 – без I, 2 – с I) графически выражается
Конкурентное ингибирование (1 – без I, 2 – с I) графически выражается
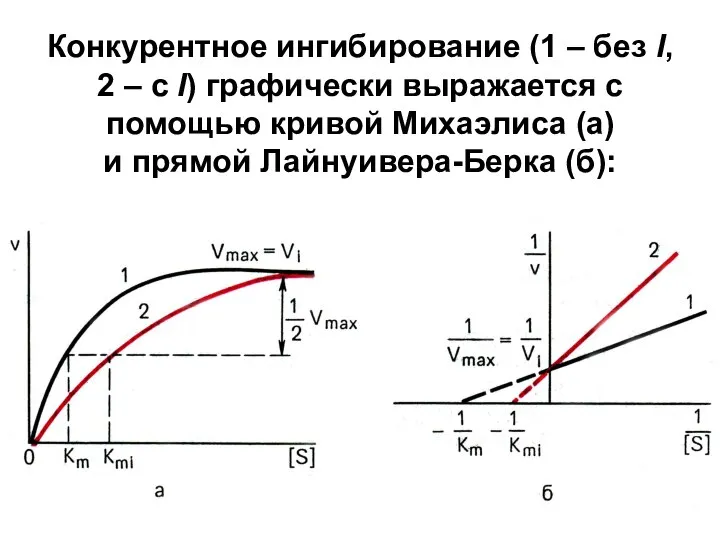
Слайд 7Т.о.,
в присутствии конкурентного I Vmax реакции не меняется, а Кm увеличивается →
Т.о.,
в присутствии конкурентного I Vmax реакции не меняется, а Кm увеличивается →

Слайд 8 На принципе конкурентного ингибирования основано действие многих ЛВ, например, группа ацетилхолинэстеразных препаратов,
На принципе конкурентного ингибирования основано действие многих ЛВ, например, группа ацетилхолинэстеразных препаратов,

Слайд 9Присоединение конкурентного I прозерина в акт.ц. АХЭ
Присоединение конкурентного I прозерина в акт.ц. АХЭ
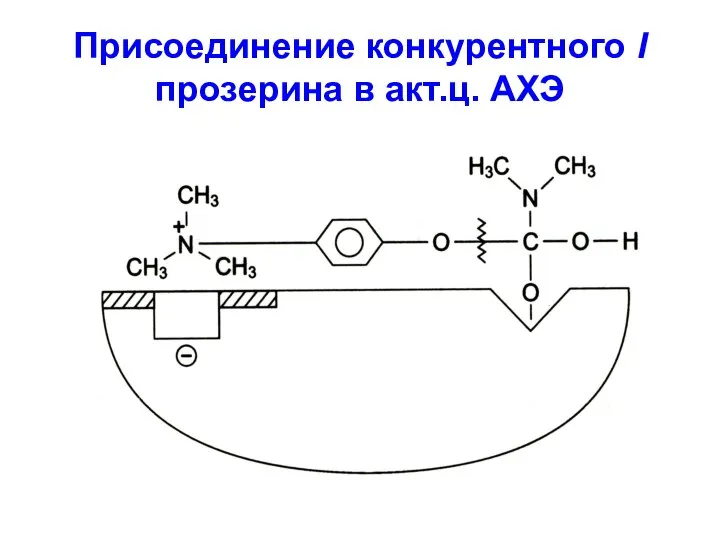
Слайд 10 Необратимо действуют фосфоорганические препараты: армин, нибуфин, хлорофос, зарин, зоман, фосфорилируя каталитический участок
Необратимо действуют фосфоорганические препараты: армин, нибуфин, хлорофос, зарин, зоман, фосфорилируя каталитический участок

Слайд 11Сульфаниламид – структурный аналог парааминобензойной кислоты
Сульфаниламид – структурный аналог парааминобензойной кислоты
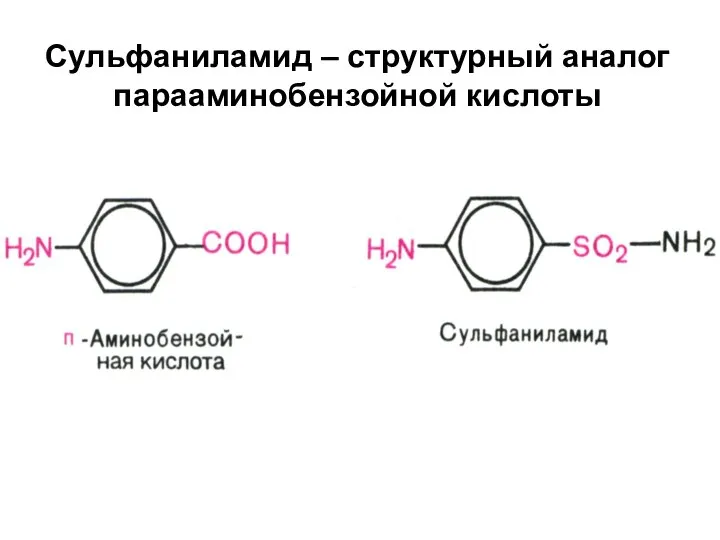
Слайд 12 Фолиевая кислота образуется в клетках бактерий, если они получают п-аминобензойную кислоту.
Сульфаниламиды
Фолиевая кислота образуется в клетках бактерий, если они получают п-аминобензойную кислоту.
Сульфаниламиды

Слайд 13 Эти Е могут использовать в качестве S сульфаниламиды → синтезируется не фолиевая
Эти Е могут использовать в качестве S сульфаниламиды → синтезируется не фолиевая

Слайд 14→ в бактериальных клетках возникает недостаток фолиевой кислоты, нарушаются все реакции, в
→ в бактериальных клетках возникает недостаток фолиевой кислоты, нарушаются все реакции, в

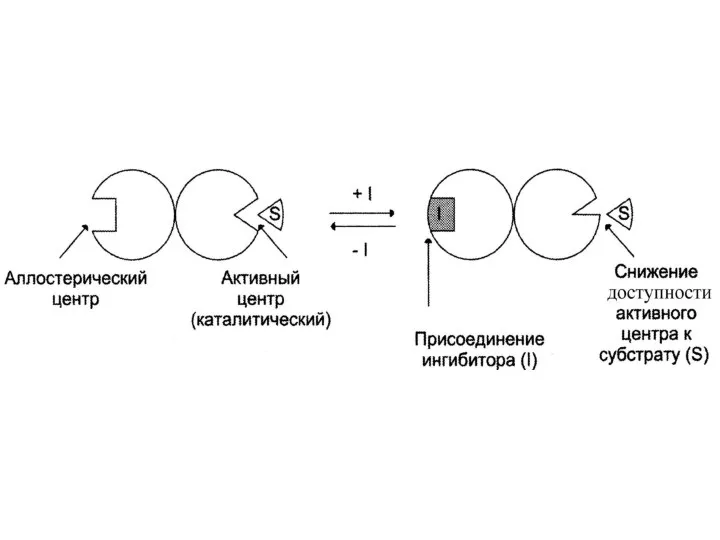
Слайд 15Неконкурентное (аллостерическое)
Это такое ингибирование, при котором I взаимодействует с Е не в
Неконкурентное (аллостерическое)
Это такое ингибирование, при котором I взаимодействует с Е не в

Слайд 17Неконкурентное ингибирование (1 – без I, 2 – с I) графически выражается
Неконкурентное ингибирование (1 – без I, 2 – с I) графически выражается
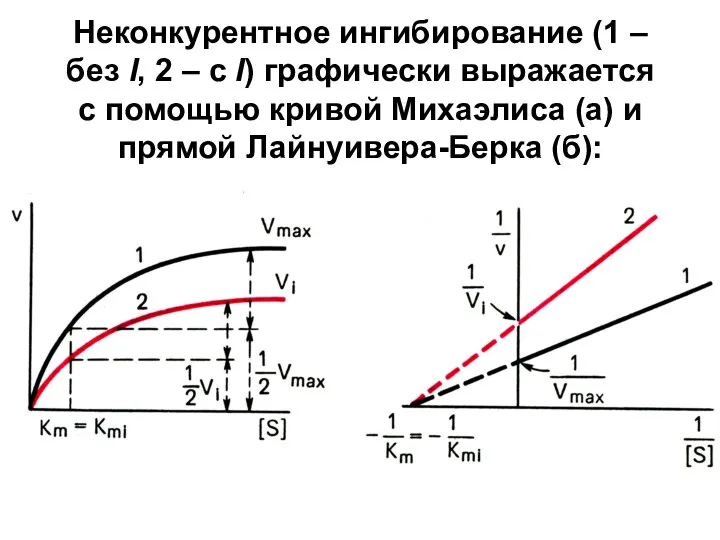
Слайд 18Т.о.
под действием неконкурентного I Vmax уменьшается, а Кm не изменяется → сродство
Т.о.
под действием неконкурентного I Vmax уменьшается, а Кm не изменяется → сродство

Слайд 19 В роли регуляторов V ферментативных реакций наиболее часто выступают: гормоны, медиаторы, ионы
В роли регуляторов V ферментативных реакций наиболее часто выступают: гормоны, медиаторы, ионы

Слайд 21
Активаторы ферментов
1. Ионы
K+, Na+, Mg2+, Mn2+, Co2+, Zn2+, Fe2+, Cl-, SO42-,
Активаторы ферментов
1. Ионы
K+, Na+, Mg2+, Mn2+, Co2+, Zn2+, Fe2+, Cl-, SO42-,

Слайд 22 2. Специфические агенты (н-р, HCl) и другие ферменты
Регуляция сводится к превращению проферментов
2. Специфические агенты (н-р, HCl) и другие ферменты
Регуляция сводится к превращению проферментов

Слайд 23Н-р, пепсиноген превращается в пепсин в результате ограниченного протеолиза:
под действием HCl отщепляется
Н-р, пепсиноген превращается в пепсин в результате ограниченного протеолиза:
под действием HCl отщепляется

Слайд 24Активность некоторых Е может регулироваться с помощью химической модификации, н-р, путем фосфорилирования-дефорилирования:
Активность некоторых Е может регулироваться с помощью химической модификации, н-р, путем фосфорилирования-дефорилирования:

Слайд 263. Аллостерические активаторы
Связываются с R, в результате чего конформация E изменяется
3. Аллостерические активаторы
Связываются с R, в результате чего конформация E изменяется

Слайд 28 4. Активаторы, способствующие объединению неактивных субъединиц E в активный надмолекулярный комплекс, имеющий
4. Активаторы, способствующие объединению неактивных субъединиц E в активный надмолекулярный комплекс, имеющий

Слайд 29
Изоферменты
Это различные молекулярные формы одного и того же фермента, катализирующие одну и
Изоферменты
Это различные молекулярные формы одного и того же фермента, катализирующие одну и

Слайд 30 Например, лактатдегидрогеназа (ЛДГ) имеет четвертичную структуру, содержит 2 типа субъединиц М и
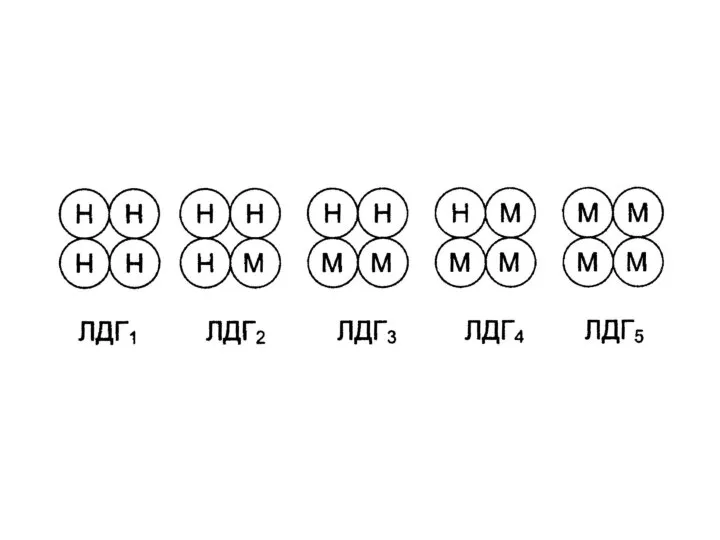
Например, лактатдегидрогеназа (ЛДГ) имеет четвертичную структуру, содержит 2 типа субъединиц М и

Слайд 31ЛДГ1 – НННН (Н4)
ЛДГ2 – НННМ (Н3М)
ЛДГ3 – ННММ (Н2М2)
ЛДГ4 – НМММ
ЛДГ1 – НННН (Н4)
ЛДГ2 – НННМ (Н3М)
ЛДГ3 – ННММ (Н2М2)
ЛДГ4 – НМММ
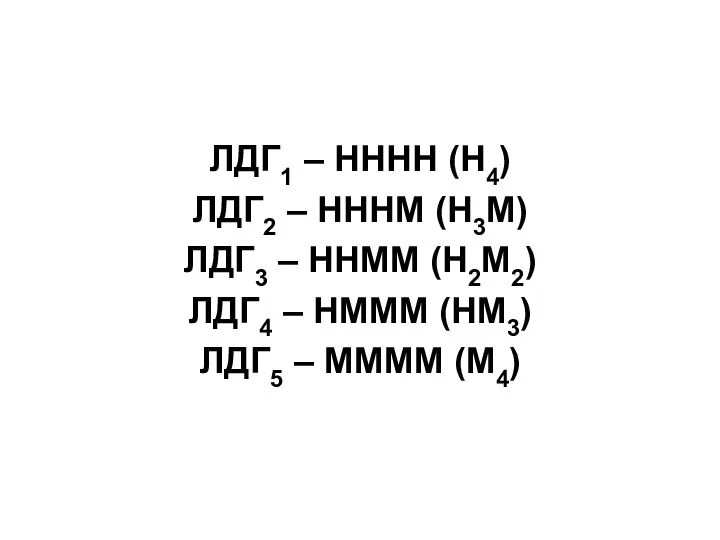
Слайд 33ЛДГ на электрофореграмме в различных органах
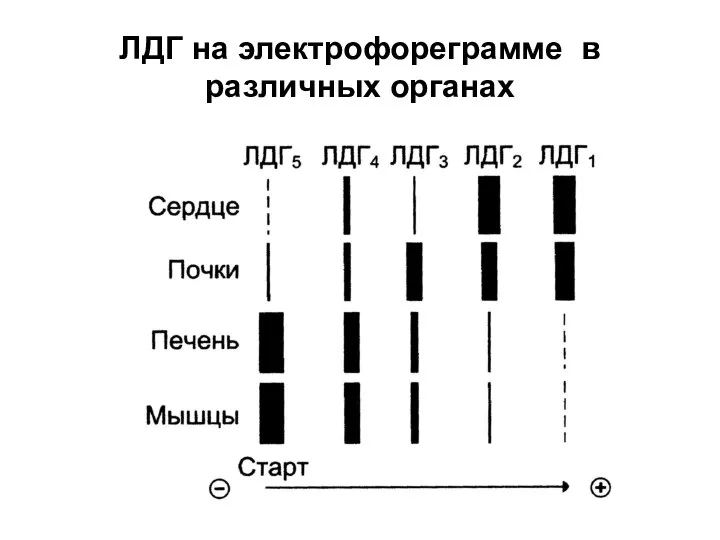
ЛДГ на электрофореграмме в различных органах
Слайд 34Мультиэнзимные комплексы (МЭК) -
надмолекулярные ферментативные системы, состоящие из различных Е, катализирующих последовательные
Мультиэнзимные комплексы (МЭК) -
надмолекулярные ферментативные системы, состоящие из различных Е, катализирующих последовательные

Слайд 35т.е.
МЭК – это группа Е, катализирующая последовательное превращение S:
А → В →
т.е.
МЭК – это группа Е, катализирующая последовательное превращение S:
А → В →
Слайд 36 Существует несколько видов МЭК, в основе организации которых лежит единство:
1 – функциональное
2
Существует несколько видов МЭК, в основе организации которых лежит единство:
1 – функциональное
2

Слайд 371
Отдельные Е объединены в полиферментную систему. Н-р, гликолиз: S1 → P1(S2) →
1
Отдельные Е объединены в полиферментную систему. Н-р, гликолиз: S1 → P1(S2) →

Слайд 382
Е образуют структурные системы с определенной функцией при помощи Е-Е взаимодействий. Н-р,
2
Е образуют структурные системы с определенной функцией при помощи Е-Е взаимодействий. Н-р,

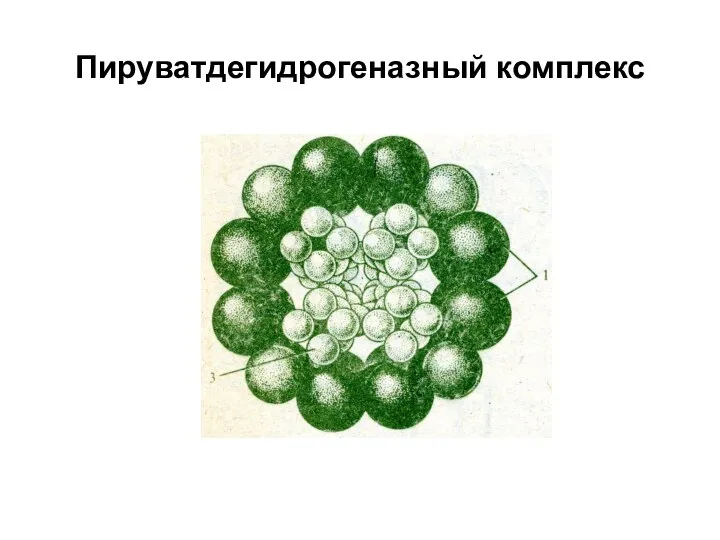
Слайд 39Пируватдегидрогеназный комплекс
Пируватдегидрогеназный комплекс
Слайд 40синтетаза высших жирных кислот, состоящая из 7 структурно связанных Е, в целом
синтетаза высших жирных кислот, состоящая из 7 структурно связанных Е, в целом

Слайд 413
Представляет комбинацию обоих типов организации. Н-р, цикл Кребса, в котором часть Е
3
Представляет комбинацию обоих типов организации. Н-р, цикл Кребса, в котором часть Е

Слайд 42Биологическое значение
Образуется «молекулярный конвейер», благодаря которому:
значительно сокращается расстояние, на которое переносятся
Биологическое значение
Образуется «молекулярный конвейер», благодаря которому:
значительно сокращается расстояние, на которое переносятся

Слайд 43Медицинская энзимология
Энзимопатология
Энзимодиагностика
Энзимотерапия
Медицинская энзимология
Энзимопатология
Энзимодиагностика
Энзимотерапия

Слайд 44Энзимопатология изучает наследственные или приобретенные дефекты ферментных систем – энзимопатии. Различают энзимопатии:
Энзимопатология изучает наследственные или приобретенные дефекты ферментных систем – энзимопатии. Различают энзимопатии:

Слайд 45Н-р, 1:
Фенилкетонурия – заболевание, при котором отсутствует Е гидроксилаза, превращающая фенилаланин в
Н-р, 1:
Фенилкетонурия – заболевание, при котором отсутствует Е гидроксилаза, превращающая фенилаланин в

Слайд 46Энзимодиагностика заключается в постановке диагноза заболевания на основе определения активности Е в
Энзимодиагностика заключается в постановке диагноза заболевания на основе определения активности Е в

Слайд 47Например:
При инфаркте миокарда увеличивается содержание ферментов ЛДГ1 и ЛДГ2, аспартатаминотрансферазы.
При вирусном гепатите
Например:
При инфаркте миокарда увеличивается содержание ферментов ЛДГ1 и ЛДГ2, аспартатаминотрансферазы.
При вирусном гепатите

Слайд 48Энзимотерапия – использование ферментов в качестве лекарственных средств.
Имеет много ограничений вследствие
Энзимотерапия – использование ферментов в качестве лекарственных средств.
Имеет много ограничений вследствие

Слайд 49Имеет следующие направления:
Заместительная терапия – использование ферментов в качестве лечебных препаратов в
Имеет следующие направления:
Заместительная терапия – использование ферментов в качестве лечебных препаратов в

Слайд 50Использование в качестве дополнительных терапевтических средств
(н-р, различные гидролитические ферменты для ускорения
Использование в качестве дополнительных терапевтических средств
(н-р, различные гидролитические ферменты для ускорения

Слайд 51
Трудности в использовании ферментов:
нестабильность
антигенные свойства
практически невозможность доставки к клеткам-мишеням
Трудности в использовании ферментов:
нестабильность
антигенные свойства
практически невозможность доставки к клеткам-мишеням

Слайд 52Для увеличения стабильности Е их связывают с различными инертными носителями (целлюлоза, крахмал).
Для
Для увеличения стабильности Е их связывают с различными инертными носителями (целлюлоза, крахмал).
Для

Слайд 53Для направленного действия Е на мишень, на поверхности микрокапсулы прикрепляют векторную молекулу
Для направленного действия Е на мишень, на поверхности микрокапсулы прикрепляют векторную молекулу

Слайд 54Ферменты широко используются для определения содержания различных веществ в биологических жидкостях
Н-р, с
Ферменты широко используются для определения содержания различных веществ в биологических жидкостях
Н-р, с

 tkani
tkani Класс Земноводные. Внешнее строение
Класс Земноводные. Внешнее строение Строение и функции спинного мозга
Строение и функции спинного мозга Голо- и Покрытосеменные
Голо- и Покрытосеменные Естественный отбор
Естественный отбор Законы Г. Менделя
Законы Г. Менделя Строение и функции растительного побега
Строение и функции растительного побега Морфологический анализ и содержание аскорбиновой кислоты в лекарственных растениях
Морфологический анализ и содержание аскорбиновой кислоты в лекарственных растениях Может ли растение дышать
Может ли растение дышать Презентация на тему Природа вокруг нас
Презентация на тему Природа вокруг нас  Папоротники. Разнообразие и распространение папоротников
Папоротники. Разнообразие и распространение папоротников Мышцы и пластика шеи
Мышцы и пластика шеи Внутреннее строение насекомых
Внутреннее строение насекомых Гром и молния
Гром и молния 9791 (1)
9791 (1) Растения в Библии
Растения в Библии Цитология
Цитология Japanese cherry flowers
Japanese cherry flowers Жизненный цикл клетки
Жизненный цикл клетки Физиология возбудимых тканей. Синапсы
Физиология возбудимых тканей. Синапсы Птицы Тамбовской области
Птицы Тамбовской области Формирование доз внешнего и внутреннего облучения ихтиофауны реки Теча в различные периоды онтогенеза
Формирование доз внешнего и внутреннего облучения ихтиофауны реки Теча в различные периоды онтогенеза Насекомоядные растения
Насекомоядные растения Взаимосвязанные изменения органов и систем органов в онто- и филогенезе
Взаимосвязанные изменения органов и систем органов в онто- и филогенезе Методы исследования в биологии. Урок № 2, 5 класс
Методы исследования в биологии. Урок № 2, 5 класс Презентация на тему СПОСОБНОСТИ МОЗГА
Презентация на тему СПОСОБНОСТИ МОЗГА  Тип плоские черви. Класс Ресничные черви
Тип плоские черви. Класс Ресничные черви Мышцы верхней конечности
Мышцы верхней конечности