- Регуляция метаболизма у бактерий

Содержание
- 2. Существуют 2 способа регуляции метаболизма у бактерий: 1) на уровне активности ферментов, или регуляция активности ферментов;
- 3. Регуляция активности ферментов Аллостерические ферменты – белки с высокой молекулярной массой, состоящие в большинстве случаев из
- 4. Субъединица аллостерического фермента Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 5. Наиболее простой случай аллостерической регуляции – регу-ляция конечным продуктом активности первого или ключевого фермента неразветвленного биосинтетического
- 6. Превращение L-треонина в L-изолейцин включает пять ферментативных реакций: Первый фермент на пути синтеза L-изолейцина L-треонинде-заминаза является
- 7. На поверхности молекулы L-треониндезаминазы имеется два вида участков: каталитический – для связывания субстрата (L-треонина) и регуляторный
- 8. При накоплении в клетке L-изолейцина он связывается с аллостерическим центром фермента L-треониндезаминазы, подавляя его активность, и
- 9. В разветвленных метаболических путях активность аллостерических ферментов регулируется сложнее, так как от активности первого фермента зависит
- 10. - кумулятивное, или аддитивное ингибирование – присоединение к ферменту одного конечного продукта частично снижает его активность,
- 11. В некоторых разветвленных биосинтетических путях ингибирование первого фермента осуществляется не конечными продуктами каждой из ветвей, а
- 12. Примером такого ретроингибирования является ингибирование фермента 3-дезокси-D-арабиногептулозо-7-фосфатсинтазы (ДАГФ-синтазы) у бактерий B. subtilis префенатом: Лекции по курсу
- 13. Существуют разветвленные метаболические пути, в которых регуляция осуществляется таким образом, что одновременно существуют и активация и
- 14. В настоящее время в селекции микроорганизмов – продуцентов аминокислот и других биологически активных продуктов используются методы
- 15. Регуляцию активности ферментов таким способом можно рассмотреть на примере фермента глутаминсинтетазы, катализиру-ющего превращение у бактерий глутамата
- 16. При повышении концентрации этих ионов в среде, в которой культивируются бактерии, происходит подавление синтеза фермента глутаминсинтетазы
- 17. При удалении из среды ионов аммония в клетках бактерий создается недостаток глутамина и фермент глутаминсинтетаза снова
- 18. Регуляция на уровне генов, или регуляция синтеза ферментов Ф.Жакоб и Ж. Моно предположили, что хромосома бактерий
- 19. Рассмотрим строение оперона на примере Lac-оперона Lac-оперон состоит из кодирующей области, представленной тремя структурными генами, ответственными
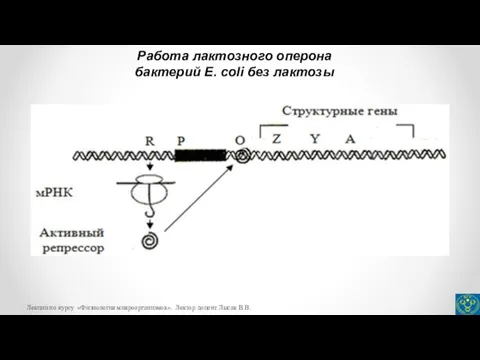
- 20. Оператор представляет собой небольшой участок ДНК, граничащий с первым структурным геном. С оператором может связываться белок-репрессор,
- 21. Таким образом, оперон – это транскрипционная единица, координированно экспрессируемая с общего промотора и контролируемая общим оператором.
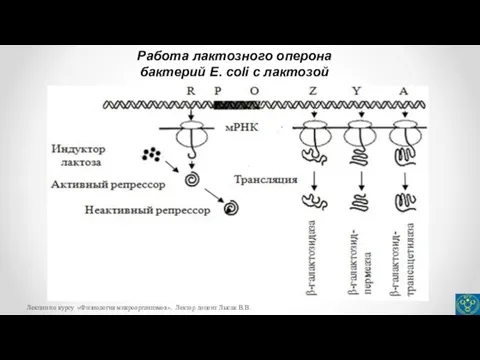
- 22. Для Lac-оперона индуктором является лактоза, которая связывается с репрессором, переводит его в неактивную форму, в результате
- 23. Работа лактозного оперона бактерий E. coli без лактозы Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак
- 24. Работа лактозного оперона бактерий E. coli c лактозой Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак
- 25. Репрессибельные опероны ответственны за синтез аминокислот аргинина, гистидина и триптофана. Максимальная транскрипция структурных генов этих оперонов
- 26. Работа триптофанового оперона бактерий E. coli (нет триптофана) Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак
- 27. Работа триптофанового оперона бактерий E. coli (избыток триптофана) Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак
- 28. Для того, чтобы понизить уровень транскрипции в присутствии триптофана, в еще большей степени, в клетках бактерий
- 29. Установлено, что аттенуация зависит не от самой аминокислоты триптофана, а от образования триптофанил-тРНК, т. е. активированной
- 30. Таким образом, и в случае индукции путем негативной регуляции (Lac-оперон) и в случае репрессии синтеза ферментов
- 31. Более высокий уровень регуляции характерен для модулона. Модулон – регуляторная система, в состав которой входят опероны
- 32. Такая регуляторная система реагирует на условия общего характера, например голодание или другие стрессовые условия, способные вызвать
- 33. Однако, как и в случае эукариотических клеток в составе мно-гоклеточного организма, регуляция одной прокариотической клет-ки не
- 35. Скачать презентацию

Слайд 3Регуляция активности ферментов
Аллостерические ферменты – белки с высокой молекулярной массой, состоящие в
Регуляция активности ферментов
Аллостерические ферменты – белки с высокой молекулярной массой, состоящие в

Слайд 4Субъединица аллостерического фермента
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Субъединица аллостерического фермента
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.

Слайд 5Наиболее простой случай аллостерической регуляции – регу-ляция конечным продуктом активности первого или
Наиболее простой случай аллостерической регуляции – регу-ляция конечным продуктом активности первого или

Слайд 6Превращение L-треонина в L-изолейцин включает пять ферментативных реакций:
Первый фермент на пути синтеза
Первый фермент на пути синтеза

Слайд 7На поверхности молекулы L-треониндезаминазы имеется два вида участков:
каталитический – для связывания
На поверхности молекулы L-треониндезаминазы имеется два вида участков:
каталитический – для связывания

Слайд 8При накоплении в клетке L-изолейцина он связывается с аллостерическим центром фермента L-треониндезаминазы,
При накоплении в клетке L-изолейцина он связывается с аллостерическим центром фермента L-треониндезаминазы,

Слайд 9В разветвленных метаболических путях активность аллостерических ферментов регулируется сложнее, так как от
В разветвленных метаболических путях активность аллостерических ферментов регулируется сложнее, так как от

Слайд 10- кумулятивное, или аддитивное ингибирование – присоединение к ферменту одного конечного продукта
- кумулятивное, или аддитивное ингибирование – присоединение к ферменту одного конечного продукта

Слайд 11В некоторых разветвленных биосинтетических путях ингибирование первого фермента осуществляется не конечными продуктами
В некоторых разветвленных биосинтетических путях ингибирование первого фермента осуществляется не конечными продуктами

Слайд 12Примером такого ретроингибирования является ингибирование фермента 3-дезокси-D-арабиногептулозо-7-фосфатсинтазы (ДАГФ-синтазы) у бактерий B. subtilis
Примером такого ретроингибирования является ингибирование фермента 3-дезокси-D-арабиногептулозо-7-фосфатсинтазы (ДАГФ-синтазы) у бактерий B. subtilis

Слайд 13Существуют разветвленные метаболические пути, в которых регуляция осуществляется таким образом, что одновременно
Существуют разветвленные метаболические пути, в которых регуляция осуществляется таким образом, что одновременно

Слайд 14В настоящее время в селекции микроорганизмов – продуцентов аминокислот и других биологически
В настоящее время в селекции микроорганизмов – продуцентов аминокислот и других биологически

Слайд 15Регуляцию активности ферментов таким способом можно рассмотреть на примере фермента глутаминсинтетазы, катализиру-ющего
Регуляцию активности ферментов таким способом можно рассмотреть на примере фермента глутаминсинтетазы, катализиру-ющего

Слайд 16При повышении концентрации этих ионов в среде, в которой культивируются бактерии, происходит
При повышении концентрации этих ионов в среде, в которой культивируются бактерии, происходит

Слайд 17При удалении из среды ионов аммония в клетках бактерий создается недостаток глутамина
При удалении из среды ионов аммония в клетках бактерий создается недостаток глутамина

Слайд 18Регуляция на уровне генов, или регуляция синтеза ферментов
Ф.Жакоб и Ж. Моно предположили,
Регуляция на уровне генов, или регуляция синтеза ферментов
Ф.Жакоб и Ж. Моно предположили,

Слайд 19Рассмотрим строение оперона на примере Lac-оперона
Lac-оперон состоит из кодирующей области, представленной тремя
Рассмотрим строение оперона на примере Lac-оперона
Lac-оперон состоит из кодирующей области, представленной тремя

Слайд 20Оператор представляет собой небольшой участок ДНК, граничащий с первым структурным геном. С
Оператор представляет собой небольшой участок ДНК, граничащий с первым структурным геном. С

Слайд 21Таким образом, оперон – это транскрипционная единица, координированно экспрессируемая с общего промотора
Таким образом, оперон – это транскрипционная единица, координированно экспрессируемая с общего промотора

Слайд 22Для Lac-оперона индуктором является лактоза, которая связывается с репрессором, переводит его в
Для Lac-оперона индуктором является лактоза, которая связывается с репрессором, переводит его в

Слайд 23Работа лактозного оперона
бактерий E. coli без лактозы
Лекции по курсу «Физиология микроорганизмов».
Работа лактозного оперона
бактерий E. coli без лактозы
Лекции по курсу «Физиология микроорганизмов».
Слайд 24Работа лактозного оперона
бактерий E. coli c лактозой
Лекции по курсу «Физиология микроорганизмов».
Работа лактозного оперона
бактерий E. coli c лактозой
Лекции по курсу «Физиология микроорганизмов».
Слайд 25Репрессибельные опероны ответственны за синтез аминокислот аргинина, гистидина и триптофана.
Максимальная транскрипция
Репрессибельные опероны ответственны за синтез аминокислот аргинина, гистидина и триптофана.
Максимальная транскрипция

Слайд 26Работа триптофанового оперона бактерий E. coli
(нет триптофана)
Лекции по курсу «Физиология микроорганизмов». Лектор
Работа триптофанового оперона бактерий E. coli
(нет триптофана)
Лекции по курсу «Физиология микроорганизмов». Лектор

Слайд 27Работа триптофанового оперона бактерий E. coli
(избыток триптофана)
Лекции по курсу «Физиология микроорганизмов». Лектор
Работа триптофанового оперона бактерий E. coli
(избыток триптофана)
Лекции по курсу «Физиология микроорганизмов». Лектор

Слайд 28Для того, чтобы понизить уровень транскрипции в присутствии триптофана, в еще большей
Для того, чтобы понизить уровень транскрипции в присутствии триптофана, в еще большей

Слайд 29Установлено, что аттенуация зависит не от самой аминокислоты триптофана, а от образования
Установлено, что аттенуация зависит не от самой аминокислоты триптофана, а от образования

Слайд 30Таким образом, и в случае индукции путем негативной регуляции (Lac-оперон) и в
Таким образом, и в случае индукции путем негативной регуляции (Lac-оперон) и в

Слайд 31Более высокий уровень регуляции характерен для модулона.
Модулон – регуляторная система, в
Более высокий уровень регуляции характерен для модулона.
Модулон – регуляторная система, в

Слайд 32Такая регуляторная система реагирует на условия общего характера, например голодание или другие
Такая регуляторная система реагирует на условия общего характера, например голодание или другие

Слайд 33Однако, как и в случае эукариотических клеток в составе мно-гоклеточного организма, регуляция
Однако, как и в случае эукариотических клеток в составе мно-гоклеточного организма, регуляция

 Митоз. Деление клеток
Митоз. Деление клеток Автономная нервная система
Автономная нервная система Презентация на тему Регенерация дождевого червя
Презентация на тему Регенерация дождевого червя  Trichomonas vaginalis
Trichomonas vaginalis Крылатые насекомые
Крылатые насекомые Движение в неживой и живой природе
Движение в неживой и живой природе Презентация на тему Основные методы селекции растений и животных
Презентация на тему Основные методы селекции растений и животных  Класс Споровики (sporozoa)
Класс Споровики (sporozoa) Строение клетки
Строение клетки Ptáci 1 - v poznávačce bude 7 zástupců!
Ptáci 1 - v poznávačce bude 7 zástupců! Презентация на тему Пингвины
Презентация на тему Пингвины  Углеводы. Липиды
Углеводы. Липиды Приставки и корни в биологии. Как легко запомнить и вспомнить термины
Приставки и корни в биологии. Как легко запомнить и вспомнить термины Особенности метаболизма азотистых оснований
Особенности метаболизма азотистых оснований Удивительные растения
Удивительные растения Горох. Строение растения и его особенности
Горох. Строение растения и его особенности Sloth
Sloth Биосинтез белка
Биосинтез белка Презентация на тему Человек как биологический вид
Презентация на тему Человек как биологический вид  Яблоко обыкновенное
Яблоко обыкновенное Микробиом. Многочисленные разнообразные ассоциации микроорганизмов
Микробиом. Многочисленные разнообразные ассоциации микроорганизмов Эволю́ция - необратимое историческое развитие живой природы. Вводная 11 класс
Эволю́ция - необратимое историческое развитие живой природы. Вводная 11 класс Проверочная работа Грибы
Проверочная работа Грибы Царство Растений
Царство Растений Одомашнивание собак
Одомашнивание собак Викторина по биологии Всеросс 2013-2014
Викторина по биологии Всеросс 2013-2014 Органы женского малого таза
Органы женского малого таза Рациональное использование луковичных растений на примере Tulipa hibrida
Рациональное использование луковичных растений на примере Tulipa hibrida