- Секреторная мембранная система. Аппарат Гольджи. Лекция 9

Содержание
- 2. Секреторная мембранная система В клетках эукариот белки, предназначенные для выведения из клетки, для внутриклеточного использования (гидролитические
- 3. Общая схема строения секреторной мембранной системы и направления транспортных потоков в клетке (антероградный и ретроградный транспорт)
- 4. Везикулярный транспорт между разными мембранными компартментами В ходе везикулярного транспорта происходит постоянный обмен компонентами между закрытыми
- 5. Нобелевскую премию по медицине в 2013 году присудили за исследования, посвященные механизмам везикулярного транспорта. Джеймс Ротман,
- 6. Использование разных белковых покрытий для везикулярного транспорта Разные покрывающие белки «выбирают» разные транспортные везикулы/переносчики в биосинтетическом,
- 7. Как выглядят разные белковые покрытия на транспортных везикулах Molecular Cell Biology, 2014 Cell Biology, 2016 Клатриновое
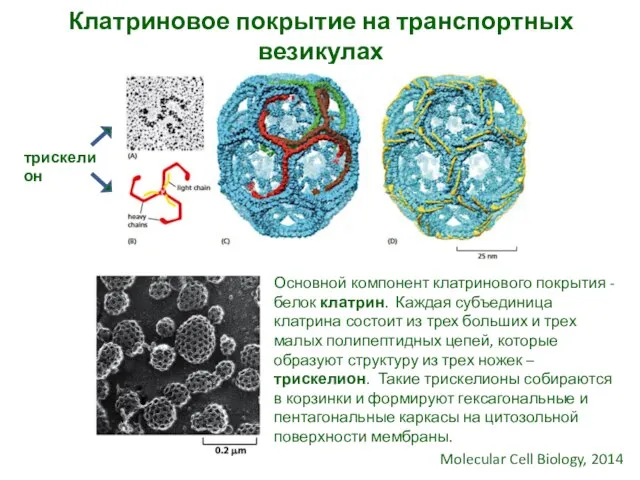
- 8. Клатриновое покрытие на транспортных везикулах Molecular Cell Biology, 2014 Основной компонент клатринового покрытия - белок клатрин.
- 9. Сборка и разборка клатринового покрытия Рецепторы «собирают» молекулы карго (белков) для упаковки в везикулу. Сборка покрытия
- 10. Сборка клатринового, COPI и COPII покрытий нужна для сортировки рецепторов, связанных с карго, и контролируется белками
- 11. Формирование COPII покрытия на везикулах, отщепляющихся от ERES (ER Exit Sites) Molecular Cell Biology, 2014 1
- 12. При образовании везикул-переносчиков, на донорских мембранах собирается белковое покрытие из клатрина, COPI или COPII, которое сортирует
- 13. Rab белки относятся к семейству малых ГТФаз Rab-ГТФазы обеспечивают взаимодействие между транспортными везикулами и «docking complexes»
- 14. Локализация белков семейства Rab на клеточных мембранах (Bhuin and Roy, 2014)
- 15. Эффекторные белки для Rab (Bhuin and Roy, 2014)
- 16. Эффекторные белки на примере ”tethering proteins” Эффекторные белки (гольджины GMAP-120) отходят от таргетной мембраны и захватывают
- 17. Непосредственное слияние мембран катализируется белками семейства SNARE В функциональном плане SNARE белки существуют в виде комплементарных
- 18. Белки семейства SNARE - SNAP (Soluble NSF Attachment Protein) REceptor") [NSF -N-ethylmaleimide-sensitive factor/N-ethylmaleimide sensitive fusion proteins]
- 19. R-SNARE работают как v-SNARE и Q-SNARE работают как t-SNARE
- 20. Как Rab белки, эффекторные белки и комплекс SNARE участвуют в узнавании, прикреплении и слиянии транспортных везикул
- 21. Mechanisms of vesicle budding and fusion (Bonifacino and Glick, 2004) После завершения слияния везикул, коровый комплекс
- 22. Антероградный и ретроградный транспорт между ЭПР и аппаратом Гольджи Molecular Cell Biology, 2014 Возвращение резидентных белков
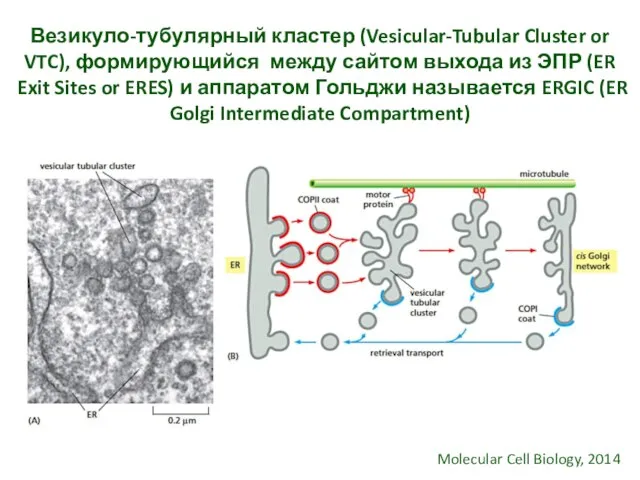
- 23. Везикуло-тубулярный кластер (Vesicular-Tubular Cluster or VTC), формирующийся между сайтом выхода из ЭПР (ER Exit Sites or
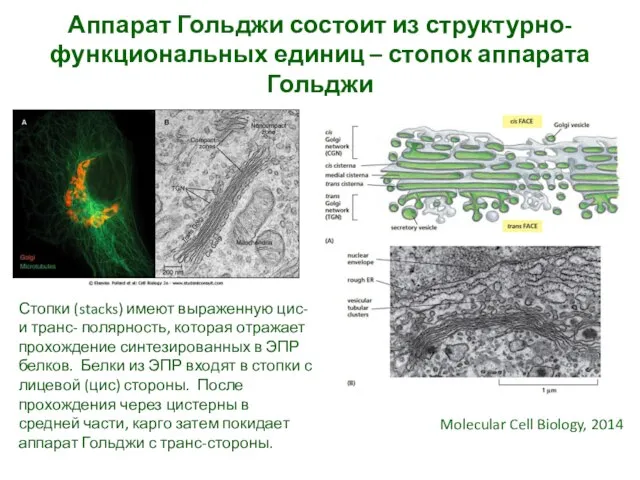
- 24. Аппарат Гольджи состоит из структурно-функциональных единиц – стопок аппарата Гольджи Molecular Cell Biology, 2014 Стопки (stacks)
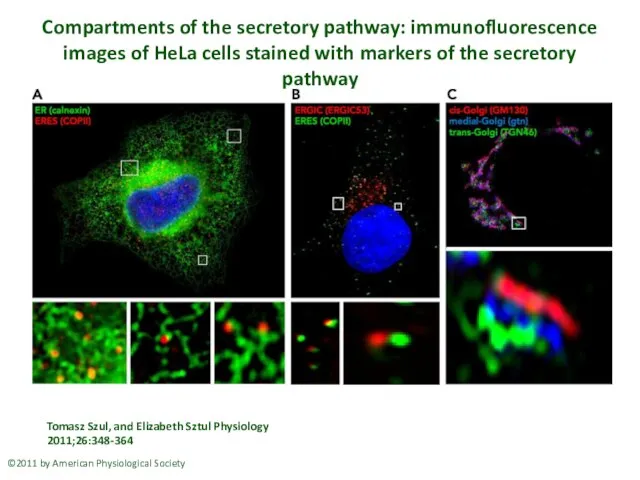
- 25. Compartments of the secretory pathway: immunofluorescence images of HeLa cells stained with markers of the secretory
- 26. Функции комплекса Гольджи 1) В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование); соединение синтезированных олигосахаридов
- 27. Функциональная компартментализация комплекса Гольджи подразумевает то, что функции, связанные с модификацией, сортировкой и адресованием карго, выполняются
- 28. ФУНКЦИИ АППАРАТА ГОЛЬДЖИ: 1. ПРОЦЕССИНГ ГЛИКОПРОТЕИДОВ (ВТОРИЧНОЕ ГЛИКОЗИЛИРОВАНИЕ) И СИНТЕЗ ПРОТЕОГЛИКАНОВ
- 29. В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование) Олигосахаридный предшественник добавленный к белку в ходе
- 30. Первичный тримминг олигосахаридных остатков в ЭПР Molecular Cell Biology, 2003 (ed. Lodish H.) Присоединение сахарных остатков
- 31. Два основных класса N-связанных олигосахаридов, обнаруженных среди зрелых гликопротеидов млекопитающих Molecular Cell Biology, 2014
- 32. Образование высокоманнозных и сложных N-связанных олигосахаридов начинается в среднем участке, и завершается в транс-сети комплекса Гольджи
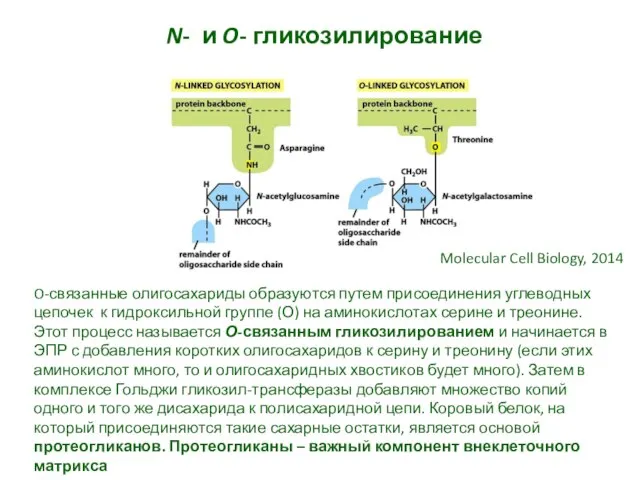
- 33. N- и O- гликозилирование Molecular Cell Biology, 2014 O-связанные олигосахариды образуются путем присоединения углеводных цепочек к
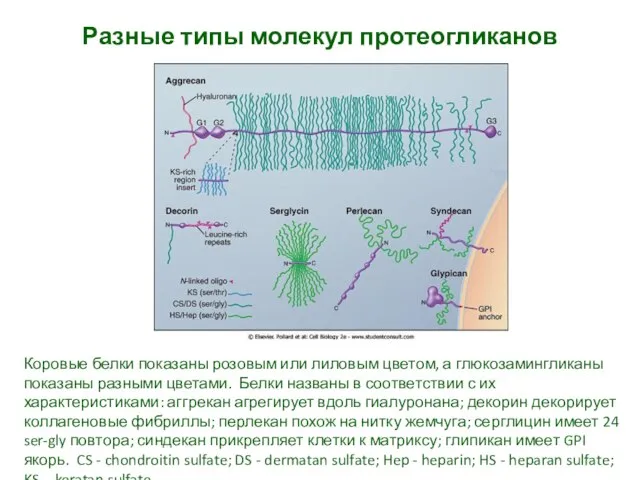
- 34. Разные типы молекул протеогликанов Коровые белки показаны розовым или лиловым цветом, а глюкозамингликаны показаны разными цветами.
- 35. Биосинтез и метаболизм липидов в комплексе Гольджи Одной из функций комплекса Гольджи является синтез сфинголипидов. Основой
- 37. Скачать презентацию
Слайд 2Секреторная мембранная система
В клетках эукариот белки, предназначенные для выведения из клетки, для
Секреторная мембранная система
В клетках эукариот белки, предназначенные для выведения из клетки, для

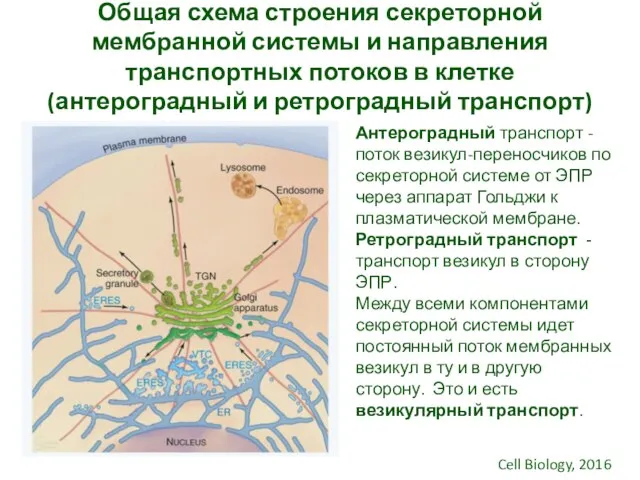
Слайд 3Общая схема строения секреторной мембранной системы и направления транспортных потоков в клетке
Общая схема строения секреторной мембранной системы и направления транспортных потоков в клетке
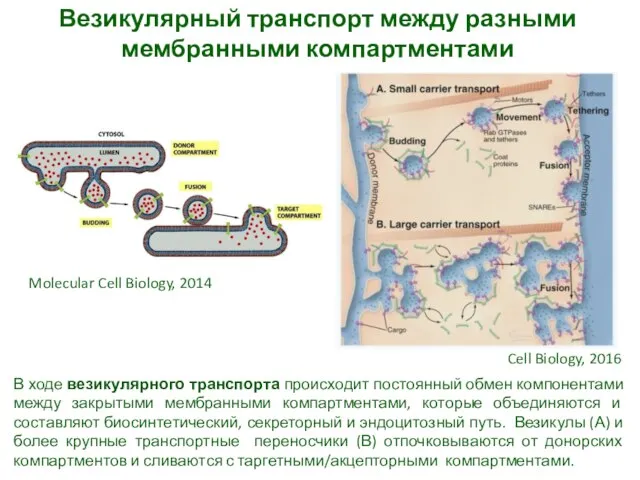
Слайд 4Везикулярный транспорт между разными мембранными компартментами
В ходе везикулярного транспорта происходит постоянный обмен
Везикулярный транспорт между разными мембранными компартментами
В ходе везикулярного транспорта происходит постоянный обмен
Слайд 5Нобелевскую премию по медицине в 2013 году присудили за исследования, посвященные механизмам
Нобелевскую премию по медицине в 2013 году присудили за исследования, посвященные механизмам

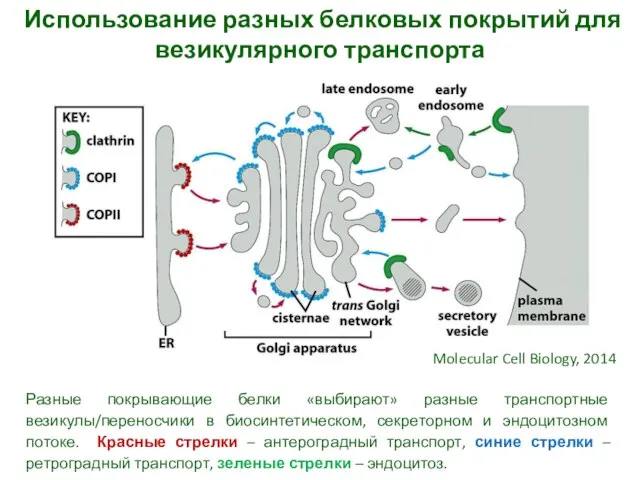
Слайд 6Использование разных белковых покрытий для везикулярного транспорта
Разные покрывающие белки «выбирают» разные транспортные
Использование разных белковых покрытий для везикулярного транспорта
Разные покрывающие белки «выбирают» разные транспортные
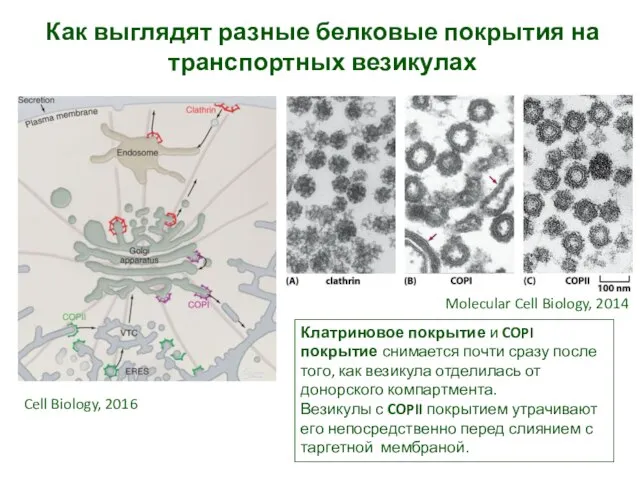
Слайд 7Как выглядят разные белковые покрытия на транспортных везикулах
Molecular Cell Biology, 2014
Cell Biology,
Как выглядят разные белковые покрытия на транспортных везикулах
Molecular Cell Biology, 2014
Cell Biology,
Слайд 8Клатриновое покрытие на транспортных везикулах
Molecular Cell Biology, 2014
Основной компонент клатринового покрытия -
Клатриновое покрытие на транспортных везикулах
Molecular Cell Biology, 2014
Основной компонент клатринового покрытия -
Слайд 9Сборка и разборка клатринового покрытия
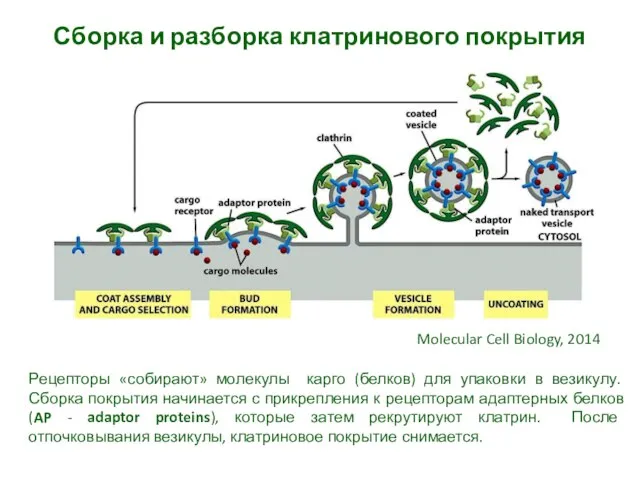
Рецепторы «собирают» молекулы карго (белков) для упаковки в
Сборка и разборка клатринового покрытия
Рецепторы «собирают» молекулы карго (белков) для упаковки в
Слайд 10Сборка клатринового, COPI и COPII покрытий нужна для сортировки рецепторов, связанных с
Сборка клатринового, COPI и COPII покрытий нужна для сортировки рецепторов, связанных с

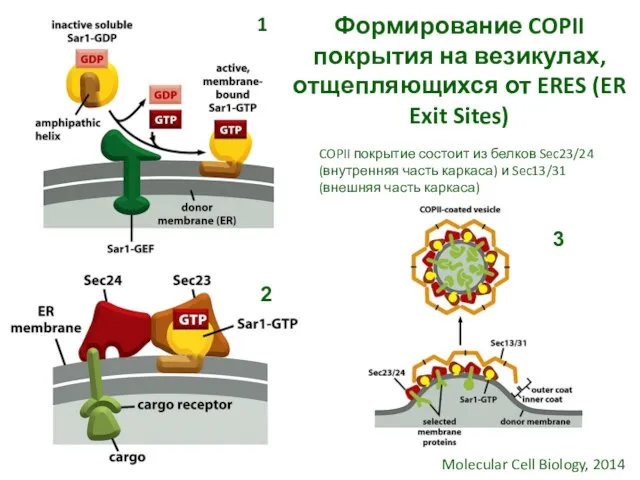
Слайд 11Формирование COPII покрытия на везикулах, отщепляющихся от ERES (ER Exit Sites)
Molecular Cell
Формирование COPII покрытия на везикулах, отщепляющихся от ERES (ER Exit Sites)
Molecular Cell
Слайд 12При образовании везикул-переносчиков, на донорских мембранах собирается белковое покрытие из клатрина, COPI
При образовании везикул-переносчиков, на донорских мембранах собирается белковое покрытие из клатрина, COPI

Слайд 13Rab белки относятся к семейству малых ГТФаз
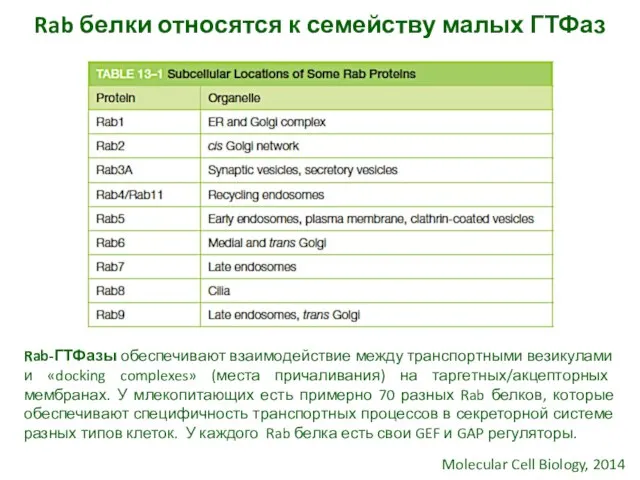
Rab-ГТФазы обеспечивают взаимодействие между транспортными везикулами
Rab белки относятся к семейству малых ГТФаз
Rab-ГТФазы обеспечивают взаимодействие между транспортными везикулами
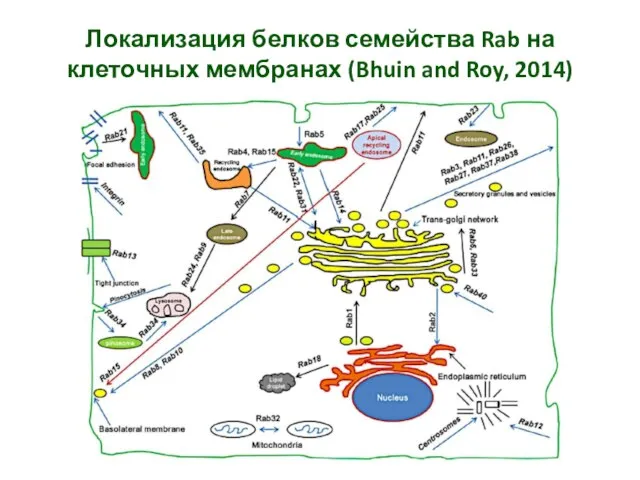
Слайд 14Локализация белков семейства Rab на клеточных мембранах (Bhuin and Roy, 2014)
Локализация белков семейства Rab на клеточных мембранах (Bhuin and Roy, 2014)
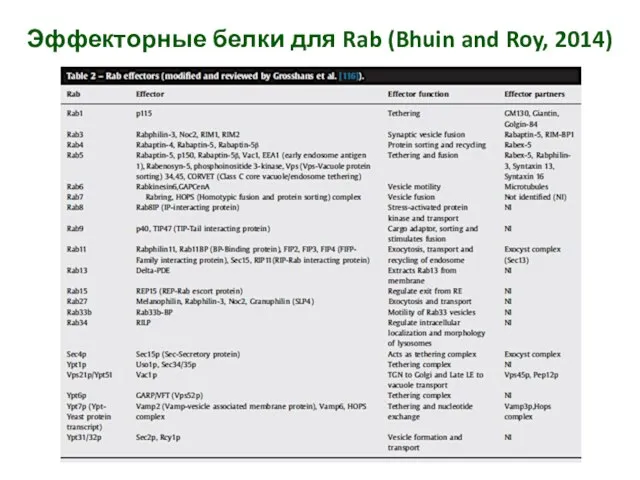
Слайд 15Эффекторные белки для Rab (Bhuin and Roy, 2014)
Эффекторные белки для Rab (Bhuin and Roy, 2014)
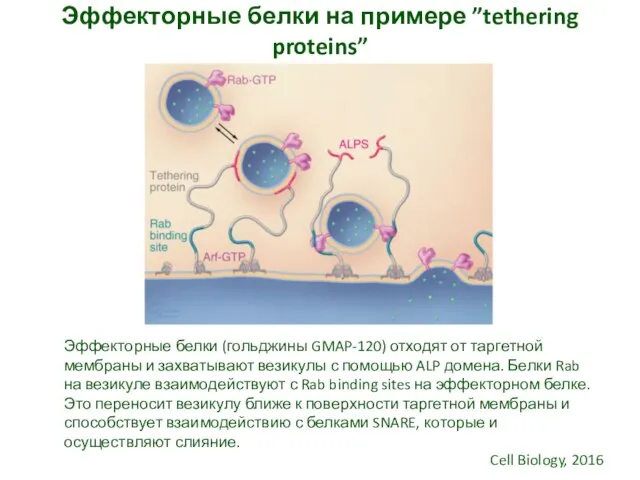
Слайд 16Эффекторные белки на примере ”tethering proteins”
Эффекторные белки (гольджины GMAP-120) отходят от таргетной
Эффекторные белки на примере ”tethering proteins”
Эффекторные белки (гольджины GMAP-120) отходят от таргетной
Слайд 17Непосредственное слияние мембран катализируется белками семейства SNARE
В функциональном плане SNARE белки
Непосредственное слияние мембран катализируется белками семейства SNARE
В функциональном плане SNARE белки

Слайд 18Белки семейства SNARE - SNAP (Soluble NSF Attachment Protein) REceptor") [NSF -N-ethylmaleimide-sensitive
Белки семейства SNARE - SNAP (Soluble NSF Attachment Protein) REceptor") [NSF -N-ethylmaleimide-sensitive
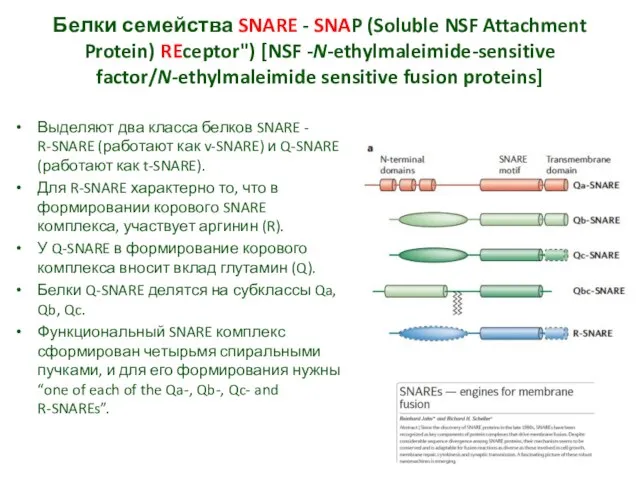
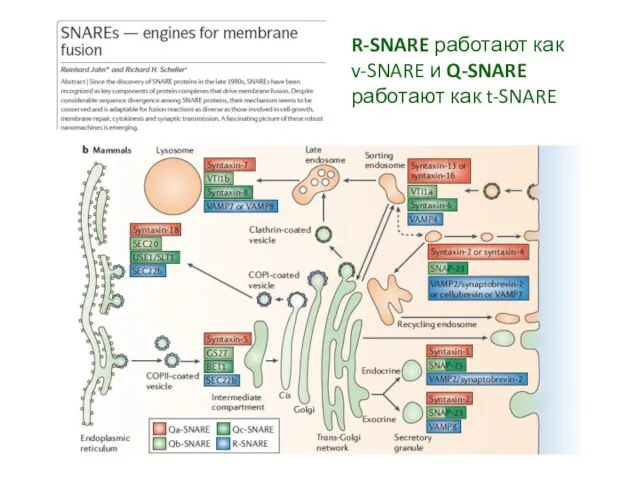
Слайд 19R-SNARE работают как v-SNARE и Q-SNARE работают как t-SNARE
R-SNARE работают как v-SNARE и Q-SNARE работают как t-SNARE
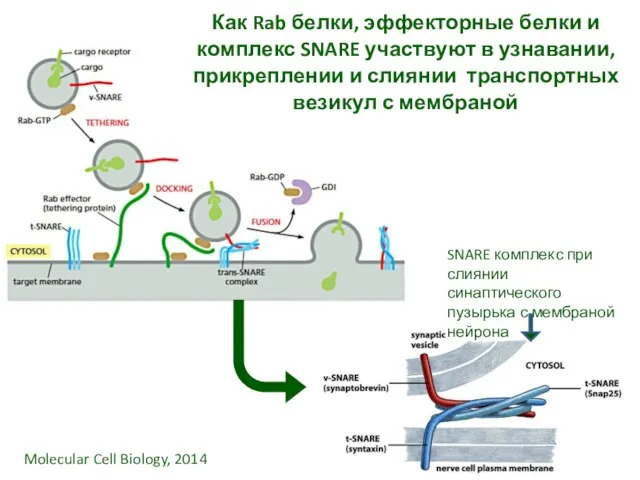
Слайд 20Как Rab белки, эффекторные белки и комплекс SNARE участвуют в узнавании, прикреплении
Как Rab белки, эффекторные белки и комплекс SNARE участвуют в узнавании, прикреплении
Слайд 21
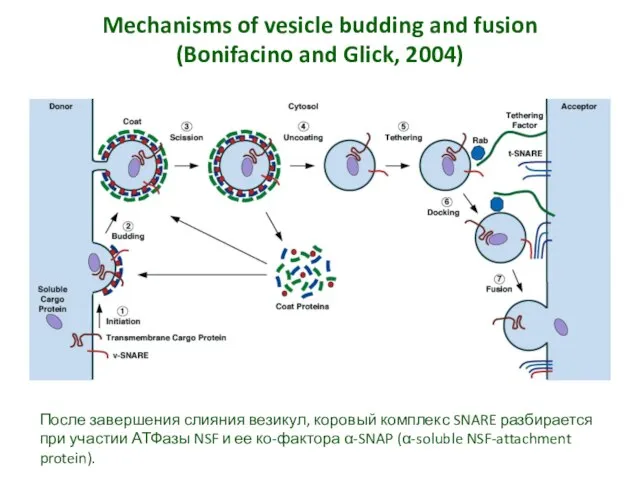
Mechanisms of vesicle budding and fusion
(Bonifacino and Glick, 2004)
После завершения слияния везикул,
Mechanisms of vesicle budding and fusion
(Bonifacino and Glick, 2004)
После завершения слияния везикул,
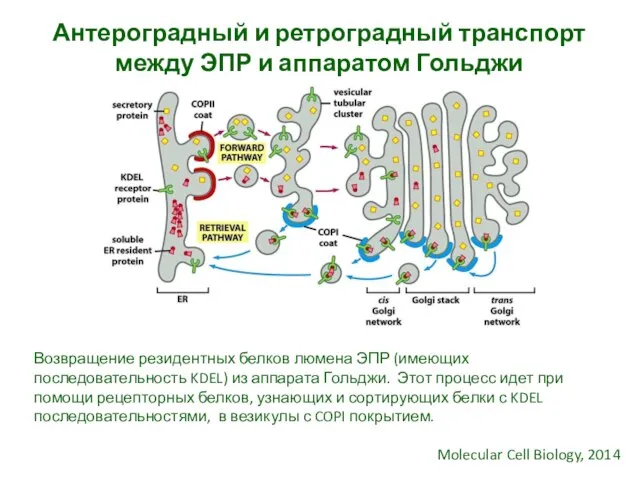
Слайд 22Антероградный и ретроградный транспорт между ЭПР и аппаратом Гольджи
Molecular Cell Biology, 2014
Возвращение
Антероградный и ретроградный транспорт между ЭПР и аппаратом Гольджи
Molecular Cell Biology, 2014
Возвращение
Слайд 23Везикуло-тубулярный кластер (Vesicular-Tubular Cluster or VTC), формирующийся между сайтом выхода из ЭПР
Везикуло-тубулярный кластер (Vesicular-Tubular Cluster or VTC), формирующийся между сайтом выхода из ЭПР
Слайд 24Аппарат Гольджи состоит из структурно-функциональных единиц – стопок аппарата Гольджи
Molecular Cell Biology,
Аппарат Гольджи состоит из структурно-функциональных единиц – стопок аппарата Гольджи
Molecular Cell Biology,
Слайд 25Compartments of the secretory pathway: immunofluorescence images of HeLa cells stained with
Compartments of the secretory pathway: immunofluorescence images of HeLa cells stained with
Слайд 26Функции комплекса Гольджи
1) В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование);
Функции комплекса Гольджи
1) В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование);

Слайд 27Функциональная компартментализация комплекса Гольджи подразумевает то, что функции, связанные с модификацией, сортировкой
Функциональная компартментализация комплекса Гольджи подразумевает то, что функции, связанные с модификацией, сортировкой
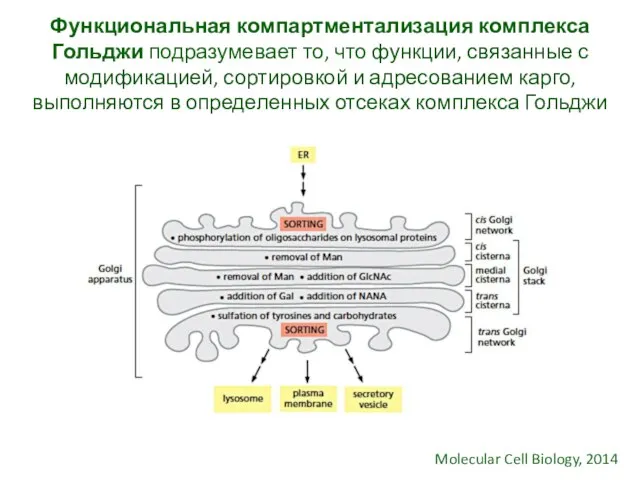
Слайд 28ФУНКЦИИ АППАРАТА ГОЛЬДЖИ:
1. ПРОЦЕССИНГ ГЛИКОПРОТЕИДОВ (ВТОРИЧНОЕ ГЛИКОЗИЛИРОВАНИЕ) И СИНТЕЗ ПРОТЕОГЛИКАНОВ
ФУНКЦИИ АППАРАТА ГОЛЬДЖИ:
1. ПРОЦЕССИНГ ГЛИКОПРОТЕИДОВ (ВТОРИЧНОЕ ГЛИКОЗИЛИРОВАНИЕ) И СИНТЕЗ ПРОТЕОГЛИКАНОВ

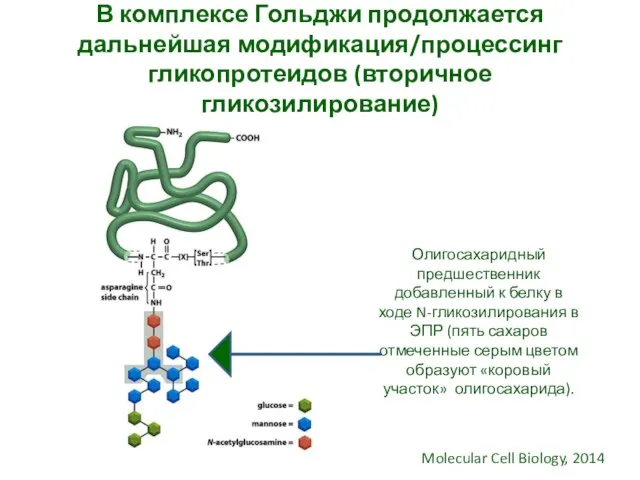
Слайд 29В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование)
Олигосахаридный предшественник добавленный к
В комплексе Гольджи продолжается дальнейшая модификация/процессинг гликопротеидов (вторичное гликозилирование)
Олигосахаридный предшественник добавленный к
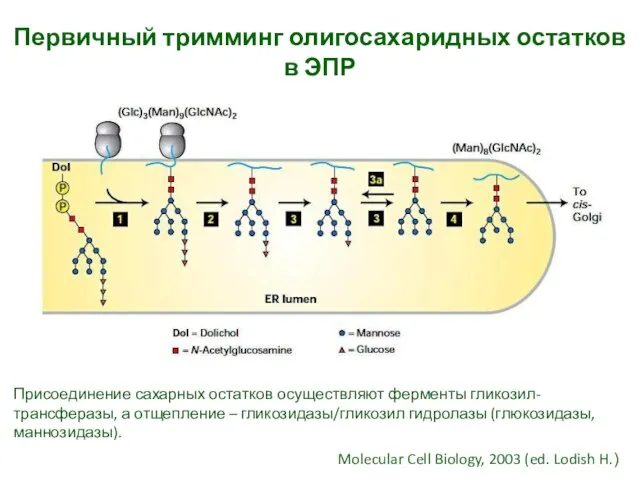
Слайд 30Первичный тримминг олигосахаридных остатков в ЭПР
Molecular Cell Biology, 2003 (ed. Lodish H.)
Присоединение
Первичный тримминг олигосахаридных остатков в ЭПР
Molecular Cell Biology, 2003 (ed. Lodish H.)
Присоединение
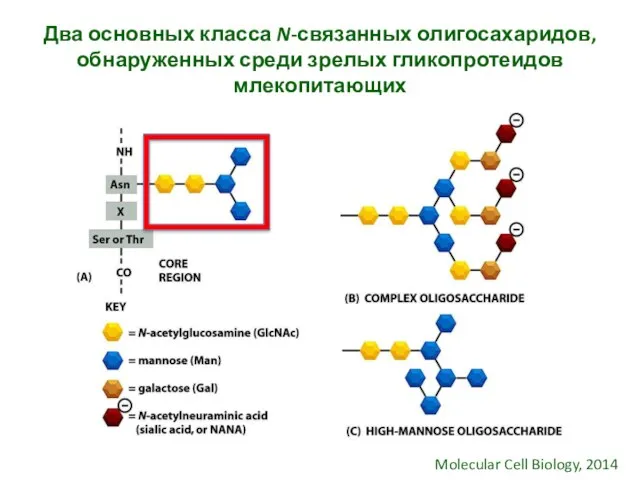
Слайд 31Два основных класса N-связанных олигосахаридов, обнаруженных среди зрелых гликопротеидов млекопитающих
Molecular Cell Biology,
Два основных класса N-связанных олигосахаридов, обнаруженных среди зрелых гликопротеидов млекопитающих
Molecular Cell Biology,
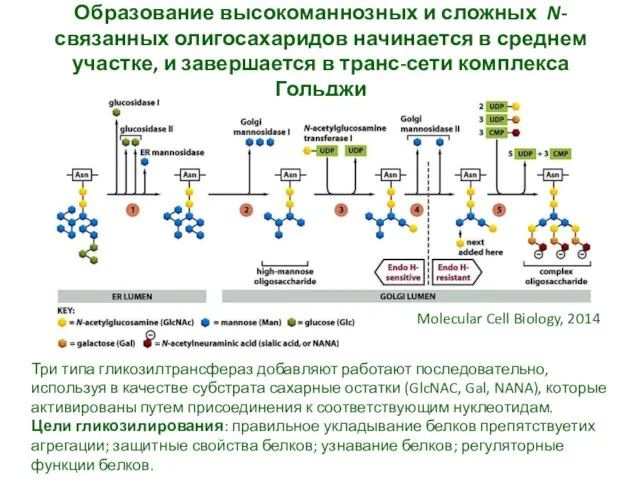
Слайд 32Образование высокоманнозных и сложных N-связанных олигосахаридов начинается в среднем участке, и завершается
Образование высокоманнозных и сложных N-связанных олигосахаридов начинается в среднем участке, и завершается
Слайд 33N- и O- гликозилирование
Molecular Cell Biology, 2014
O-связанные олигосахариды образуются путем присоединения углеводных
N- и O- гликозилирование
Molecular Cell Biology, 2014
O-связанные олигосахариды образуются путем присоединения углеводных
Слайд 34Разные типы молекул протеогликанов
Коровые белки показаны розовым или лиловым цветом, а глюкозамингликаны
Разные типы молекул протеогликанов
Коровые белки показаны розовым или лиловым цветом, а глюкозамингликаны
Слайд 35Биосинтез и метаболизм липидов в комплексе Гольджи
Одной из функций комплекса Гольджи является
Биосинтез и метаболизм липидов в комплексе Гольджи
Одной из функций комплекса Гольджи является

 Живые барометры природы
Живые барометры природы Почему люди любят динозавров
Почему люди любят динозавров Гаметогенез и дробление
Гаметогенез и дробление Пищевод ≈ 25 см
Пищевод ≈ 25 см Породы голубей
Породы голубей Орган слуха
Орган слуха Борьба за существование
Борьба за существование Движущие силы эволюции
Движущие силы эволюции Презентация по биологии на тему _Этапы развития жизни на Земле_
Презентация по биологии на тему _Этапы развития жизни на Земле_ Презентация на тему РАСТЕНИЯ-ПАРАЗИТЫ
Презентация на тему РАСТЕНИЯ-ПАРАЗИТЫ  Движущие силы психического развития
Движущие силы психического развития Кожа
Кожа Биосфера - это оболочка Земли
Биосфера - это оболочка Земли Фотоотчёт по строению ракообразных
Фотоотчёт по строению ракообразных Внешнее строение рыб
Внешнее строение рыб Зимующие птицы
Зимующие птицы Рептилии
Рептилии Клеточные и неклеточные формы жизни
Клеточные и неклеточные формы жизни Біологічна вікторина
Біологічна вікторина Влияние гормонов на животных и растения
Влияние гормонов на животных и растения Наследственность. Строение клетки
Наследственность. Строение клетки Изучение механизма наследственности
Изучение механизма наследственности Кто такие звери
Кто такие звери Сколько живут растения
Сколько живут растения Роль ЦНС в регуляции физиологических функций. Строение и организация спинного и головного мозга
Роль ЦНС в регуляции физиологических функций. Строение и организация спинного и головного мозга Вегетативная функция ЦНС
Вегетативная функция ЦНС Механизмы нервной и гуморальной регуляции сна
Механизмы нервной и гуморальной регуляции сна Обмен белков. Декарбоксилированние карбоновых кислот, обмен по амино-группе
Обмен белков. Декарбоксилированние карбоновых кислот, обмен по амино-группе