- Свойства дыхательных газов

Содержание
- 2. Диффузия часто дополняется макропереносом (циркуляция крови), но диффузия – основа перемещения газов, участвующих в дыхании Важно
- 3. До h = 100 км - постоянство состава атмосферного воздуха поддерживается конвекционными токами; выше - +
- 4. С позиций физиологов, если исключить водяные пары, газовый состав атмосферы = сonst. Равновесие между потреблением О2,
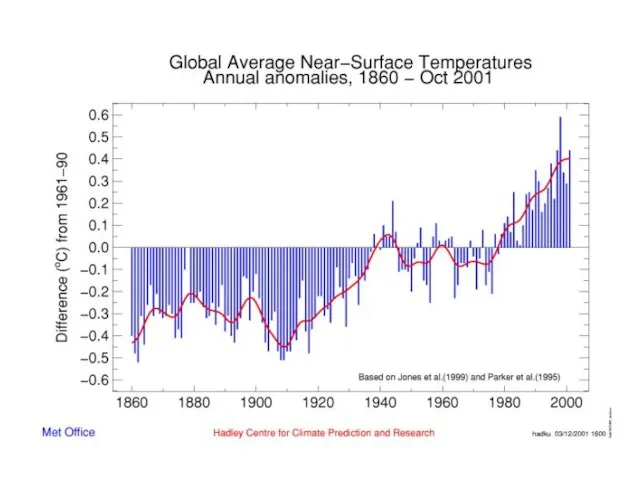
- 5. ?? Сжигание угля, нефти, природных газов ? истощение запасов О2 + возрастает % СО2 ?? Годы
- 6. Небольшое > %СО2 ? ∆ поглощения солнечной радиации ? парниковый эффект Испускаемое Землей длинноволновое излучение поглощается
- 7. ????????????????
- 9. Сильные вариации О2 в норах, внутри поверхностных слоев почвы: В почве много окисляющихся веществ (органика, сульфид
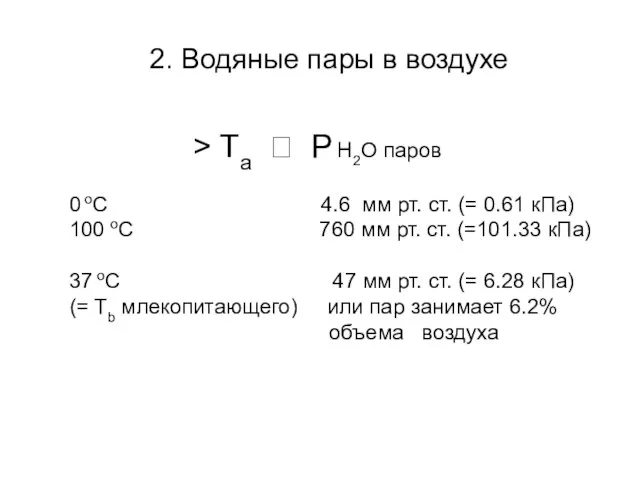
- 10. 2. Водяные пары в воздухе > Ta ? P H2O паров 0 оС 4.6 мм рт.
- 11. Воздух в легких насыщен парами при данной Tb Атмосферный воздух обычно не насыщен при данной Ta
- 12. 3. Высота и атмосферное давление h = 6000 m ? проблема с выживанием из-за недостатка О2,
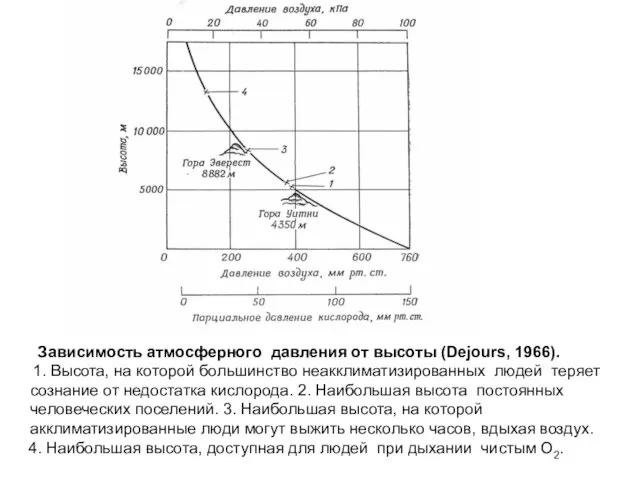
- 13. Зависимость атмосферного давления от высоты (Dejours, 1966). 1. Высота, на которой большинство неакклиматизированных людей теряет сознание
- 14. 4. Растворимость газов в воде В условиях равновесия между воздухом и открытой водной поверхностью количество газа,
- 15. 1) При определенных Р газа и Ta : О2 растворяется в 2 раза больше, чем N2
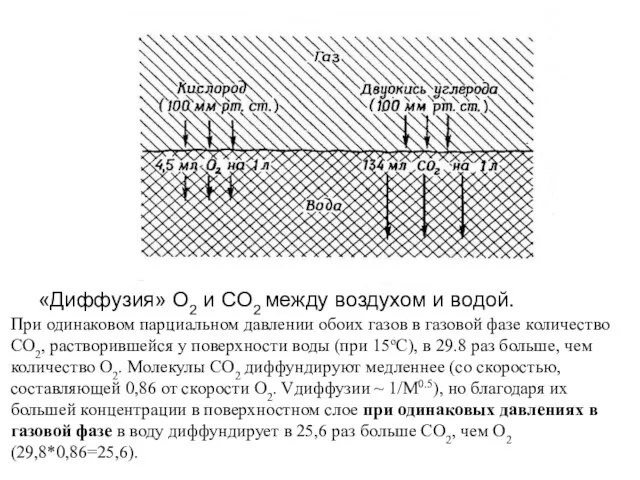
- 16. «Диффузия» O2 и CO2 между воздухом и водой. При одинаковом парциальном давлении обоих газов в газовой
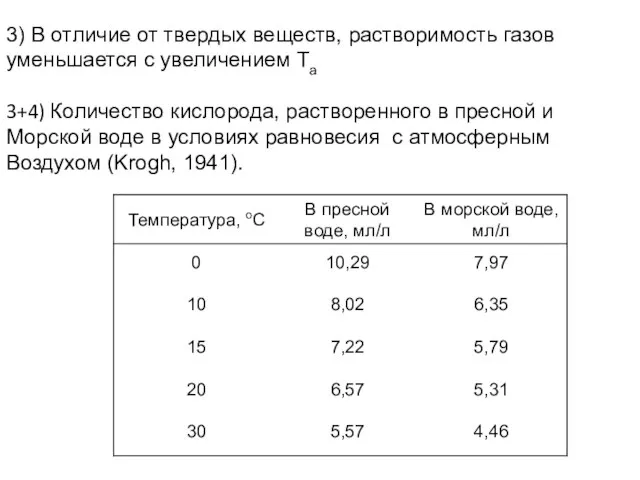
- 17. 3) В отличие от твердых веществ, растворимость газов уменьшается с увеличением Ta 3+4) Количество кислорода, растворенного
- 18. 5. Парциальное давление и напряжение газа Количество газа, растворенного в воде (жидкости), соответствует его парциальному давлению
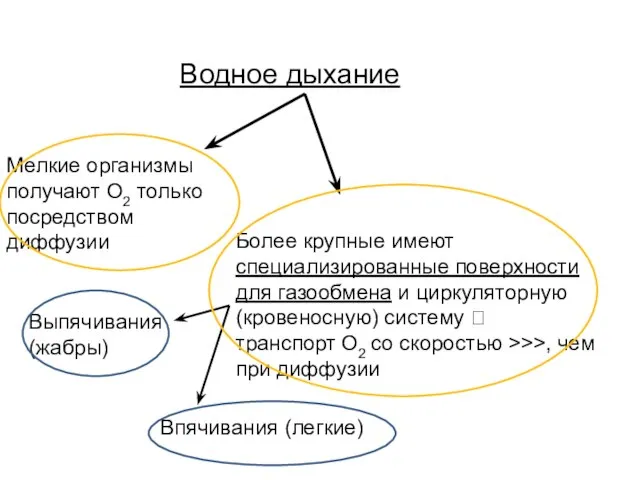
- 19. Водное дыхание Мелкие организмы получают О2 только посредством диффузии Более крупные имеют специализированные поверхности для газообмена
- 20. 1. Животные без специализированных органов дыхания Сфера: минимально возможная S при данном V R . R
- 21. Животные без специализированных органов дыхания Сфера: минимально возможная S при данном V R R – самый
- 22. Животные без специализированных органов дыхания Сфера: минимально возможная S при данном V R R – самый
- 23. Животные без специализированных органов дыхания Сфера: минимально возможная S при данном V R R – самый
- 24. Животные без специализированных органов дыхания Сфера: минимально возможная S при данном V R R – самый
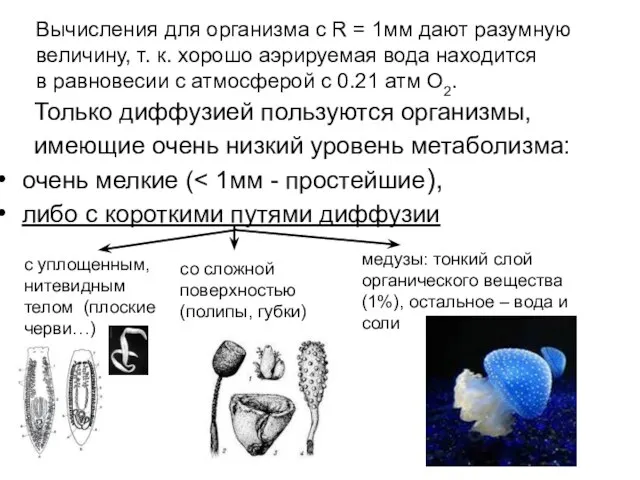
- 25. Вычисления для организма с R = 1мм дают разумную величину, т. к. хорошо аэрируемая вода находится
- 26. 2. Водные животные, обладающие органами дыхания Жабры исходно были вывернуты наружу, у рыб – вторично вошли
- 27. Вентиляция жабр Движение жабры в воде: «практично» только для небольших размеров R (для личинок поденок). Требуется
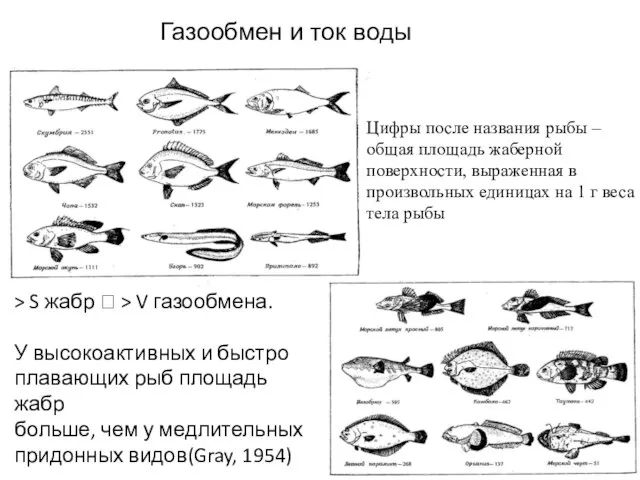
- 28. > S жабр ? > V газообмена. У высокоактивных и быстро плавающих рыб площадь жабр больше,
- 29. Жабра рыбы. Кончик одной жаберной нити форели. Вода течет между параллельно расположенными пластинками в направлении, перпендикулярном
- 30. Жабры рыб состоят из нескольких жаберных дуг. Каждая дуга несет на себе по два ряда жаберных
- 31. Противоток в жабрах рыб позволяет крови иметь на выходе из жабр почти такое же высокое напряжение
- 32. Эффективность обменных процессов между жидкостями, движущимися в одном и том же (А) и в противоположных направлениях
- 33. Воду через жабры рыб прогоняет двойная система насосов. При помощи соответствующих клапанов эти насосы обеспечивают однонаправленный
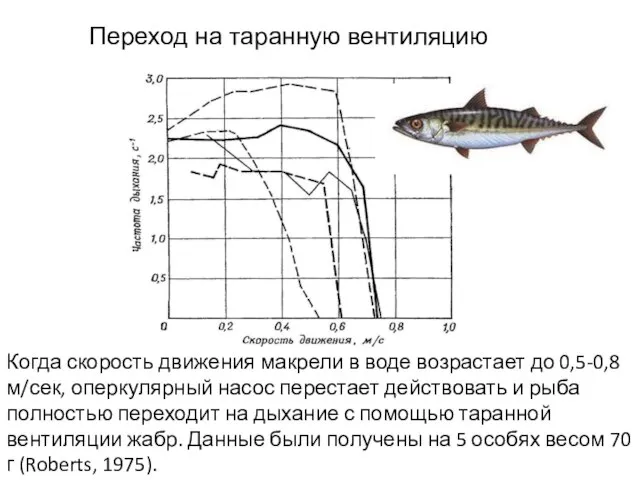
- 34. Когда скорость движения макрели в воде возрастает до 0,5-0,8 м/сек, оперкулярный насос перестает действовать и рыба
- 35. Дыхание на воздухе В широком масштабе успешная эволюционная адаптация к воздушному дыханию произошла у позвоночных, насекомых
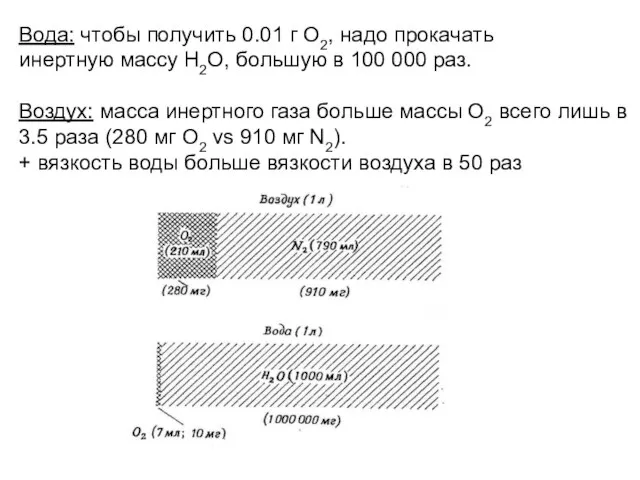
- 36. Вода: чтобы получить 0.01 г О2, надо прокачать инертную массу Н2О, большую в 100 000 раз.
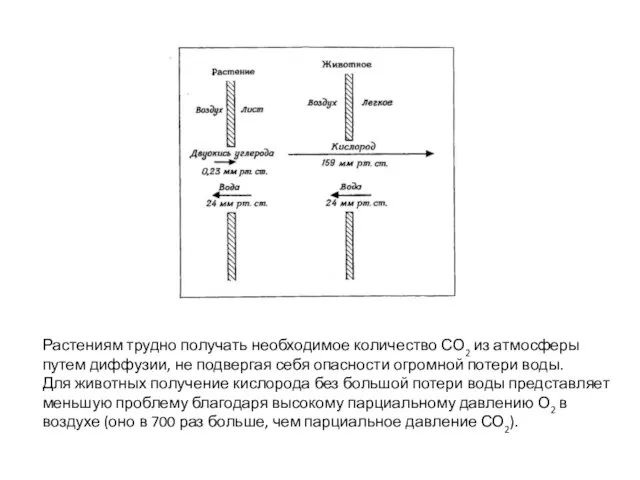
- 37. Растениям трудно получать необходимое количество СО2 из атмосферы путем диффузии, не подвергая себя опасности огромной потери
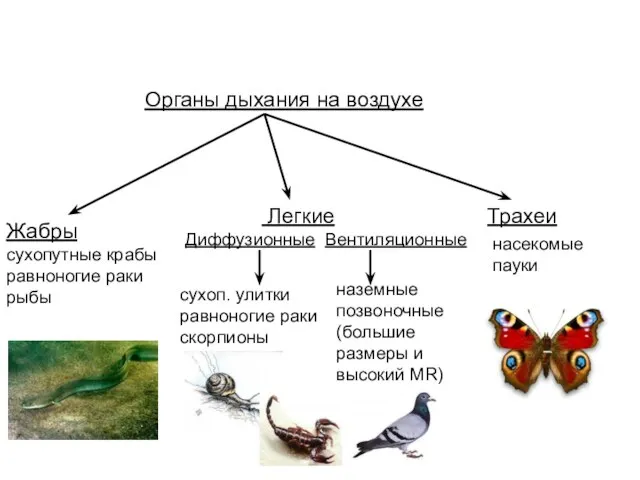
- 38. Органы дыхания на воздухе Жабры сухопутные крабы равноногие раки рыбы Трахеи Легкие Диффузионные Вентиляционные сухоп. улитки
- 39. Вентиляция легких у позвоночных Нагнетательный насос амфибий Всасывающий насос (рептилии, птицы, млекопитающие) Дыхательный цикл у лягушки
- 40. Рыбы, способные дышать воздухом Помимо двоякодышащих представители 17 родов рыб способны дышать воздухом. Экологические причины: нехватка
- 41. Рыба-ползун Anabas Индийский сом Saccobranchus Африканский шагающий сом Clarias Органы дополнительного воздушного дыхания у рыб. Варианты
- 43. Рыбы, для которых дыхание воздухом обязательно
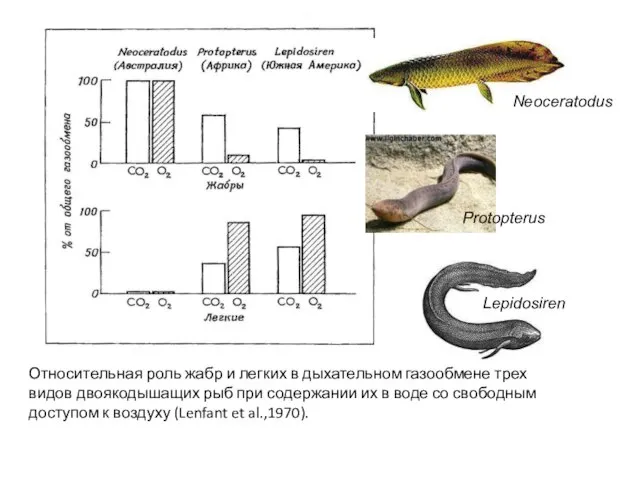
- 44. Относительная роль жабр и легких в дыхательном газообмене трех видов двоякодышащих рыб при содержании их в
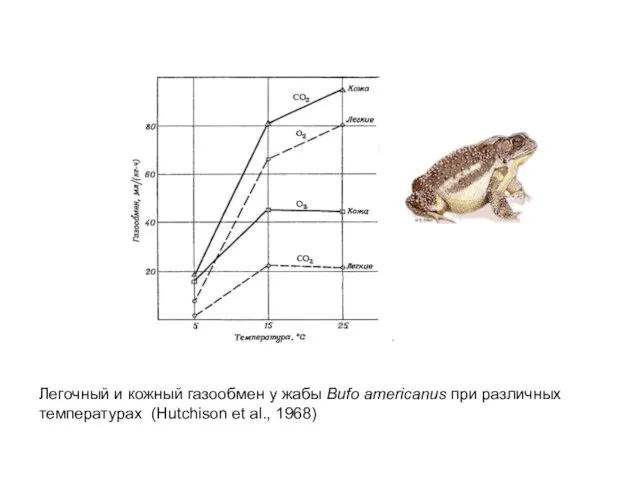
- 45. Легочный и кожный газообмен у жабы Bufo americanus при различных температурах (Hutchison et al., 1968)
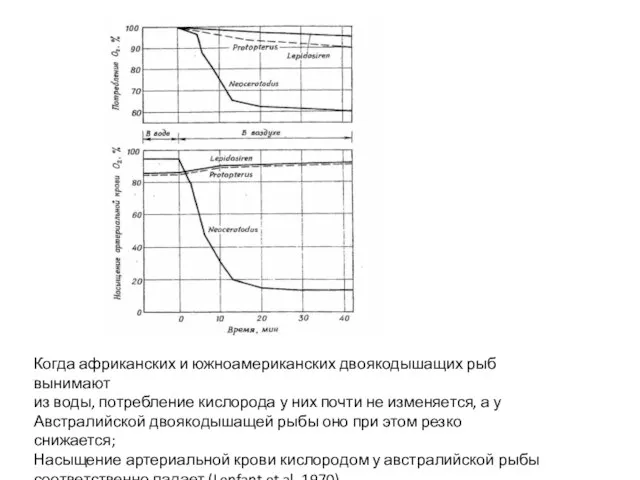
- 46. Когда африканских и южноамериканских двоякодышащих рыб вынимают из воды, потребление кислорода у них почти не изменяется,
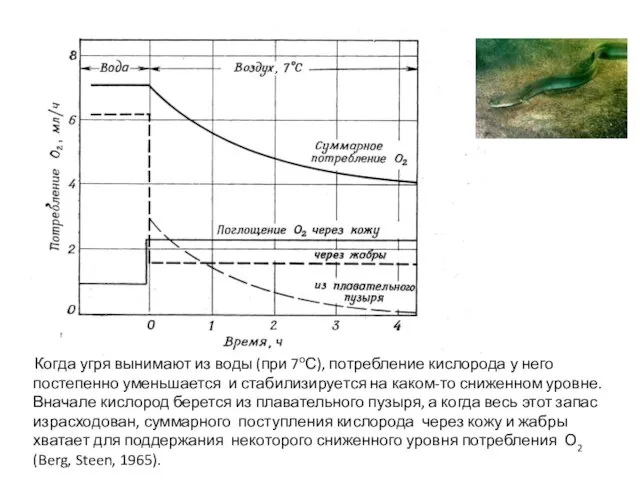
- 47. Когда угря вынимают из воды (при 7оС), потребление кислорода у него постепенно уменьшается и стабилизируется на
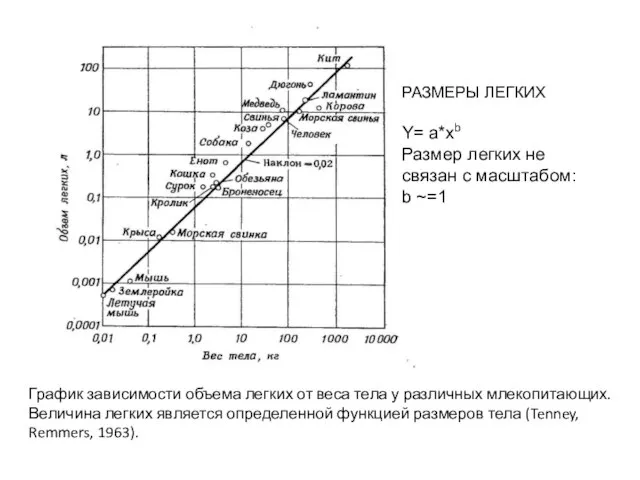
- 48. График зависимости объема легких от веса тела у различных млекопитающих. Величина легких является определенной функцией размеров
- 49. Дыхание птиц. Особенности строения и функционирования легких
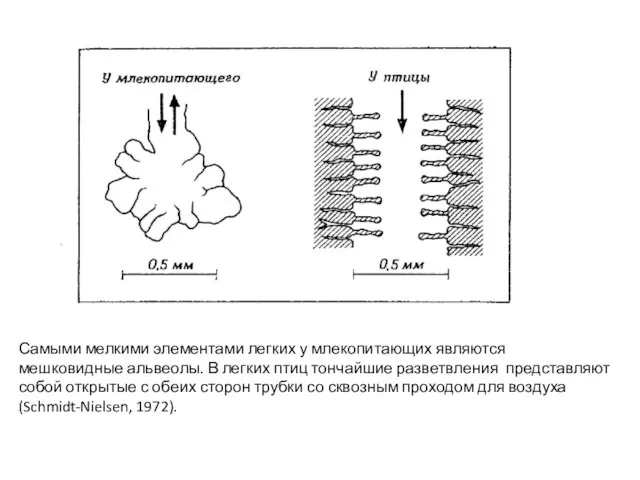
- 50. Самыми мелкими элементами легких у млекопитающих являются мешковидные альвеолы. В легких птиц тончайшие разветвления представляют собой
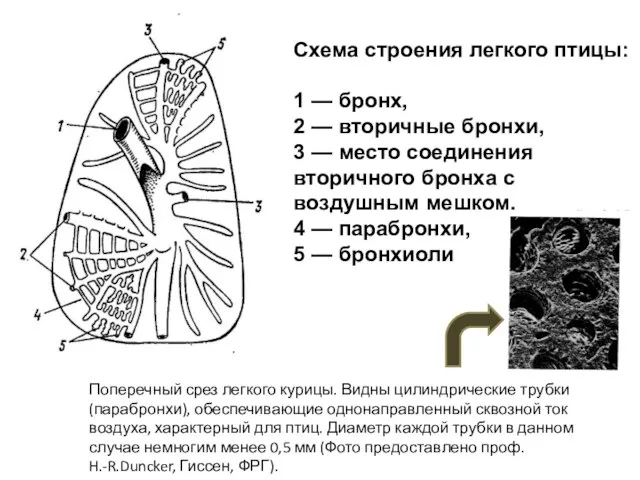
- 51. Схема строения легкого птицы: 1 — бронх, 2 — вторичные бронхи, 3 — место соединения вторичного
- 52. Объем дыхательной системы у типичной птицы и типичного млекопитающего, весом 1 кг (По данным Lasiewski, Calder,
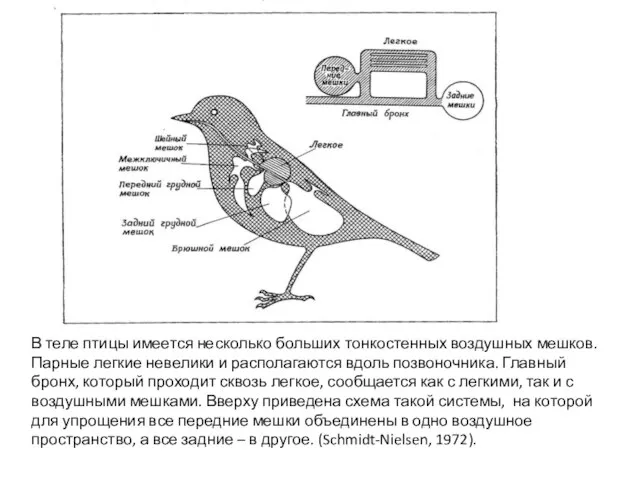
- 53. В теле птицы имеется несколько больших тонкостенных воздушных мешков. Парные легкие невелики и располагаются вдоль позвоночника.
- 54. Когда страусу дают один раз вдохнуть чистый кислород, а затем он дышит обычным воздухом, кислород играет
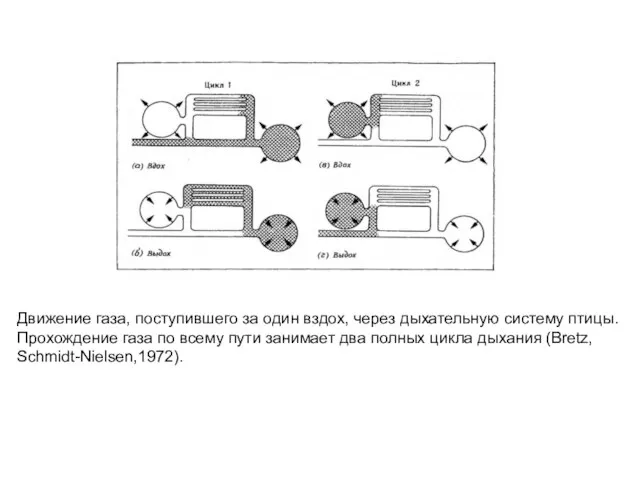
- 55. Движение газа, поступившего за один вздох, через дыхательную систему птицы. Прохождение газа по всему пути занимает
- 56. Газообмен в легком птицы на большой высоте (Schmidt-Nielsen, 1972). На упрощенной схеме показаны потоки крови и
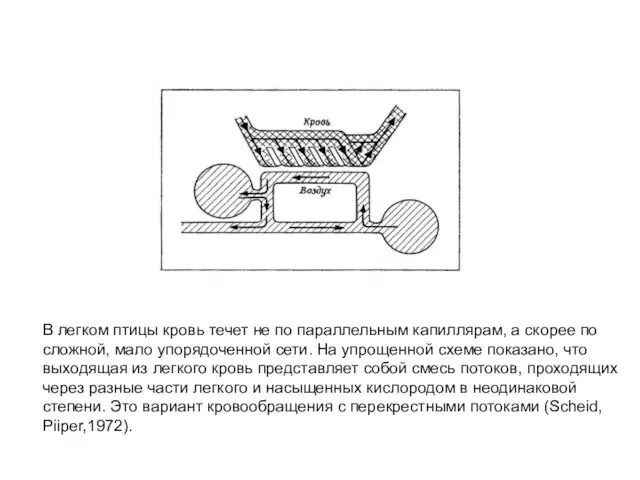
- 57. В легком птицы кровь течет не по параллельным капиллярам, а скорее по сложной, мало упорядоченной сети.
- 59. Скачать презентацию
Слайд 2Диффузия часто дополняется макропереносом (циркуляция крови),
но диффузия – основа перемещения газов, участвующих
Диффузия часто дополняется макропереносом (циркуляция крови),
но диффузия – основа перемещения газов, участвующих

Слайд 3До h = 100 км - постоянство состава атмосферного воздуха поддерживается конвекционными
До h = 100 км - постоянство состава атмосферного воздуха поддерживается конвекционными

Слайд 4С позиций физиологов,
если исключить водяные пары,
газовый состав атмосферы = сonst.
Равновесие между
С позиций физиологов,
если исключить водяные пары,
газовый состав атмосферы = сonst.
Равновесие между

Слайд 5?? Сжигание угля, нефти, природных газов ?
истощение запасов О2 + возрастает
?? Сжигание угля, нефти, природных газов ?
истощение запасов О2 + возрастает

Слайд 6Небольшое > %СО2 ? ∆ поглощения солнечной радиации ?
парниковый эффект
Испускаемое
Небольшое > %СО2 ? ∆ поглощения солнечной радиации ?
парниковый эффект
Испускаемое

Слайд 7????????????????
????????????????

Слайд 9Сильные вариации О2 в норах, внутри поверхностных
слоев почвы:
В почве много окисляющихся
Сильные вариации О2 в норах, внутри поверхностных
слоев почвы:
В почве много окисляющихся

Слайд 102. Водяные пары в воздухе
> Ta ? P H2O паров
0 оС
2. Водяные пары в воздухе
> Ta ? P H2O паров
0 оС
Слайд 11Воздух в легких насыщен парами при данной Tb
Атмосферный воздух обычно не
Воздух в легких насыщен парами при данной Tb
Атмосферный воздух обычно не

Слайд 123. Высота и атмосферное давление
h = 6000 m ? проблема с выживанием
3. Высота и атмосферное давление
h = 6000 m ? проблема с выживанием

Слайд 13 Зависимость атмосферного давления от высоты (Dejours, 1966).
1. Высота, на которой
Зависимость атмосферного давления от высоты (Dejours, 1966).
1. Высота, на которой
Слайд 144. Растворимость газов в воде
В условиях равновесия между воздухом и открытой водной
4. Растворимость газов в воде
В условиях равновесия между воздухом и открытой водной

Слайд 151) При определенных Р газа и Ta :
О2 растворяется в 2
1) При определенных Р газа и Ta :
О2 растворяется в 2

Слайд 16 «Диффузия» O2 и CO2 между воздухом и водой.
При одинаковом парциальном давлении
«Диффузия» O2 и CO2 между воздухом и водой.
При одинаковом парциальном давлении
Слайд 173) В отличие от твердых веществ, растворимость газов
уменьшается с увеличением Ta
3+4) Количество
3) В отличие от твердых веществ, растворимость газов
уменьшается с увеличением Ta
3+4) Количество
Слайд 185. Парциальное давление и напряжение газа
Количество газа, растворенного в воде (жидкости), соответствует
5. Парциальное давление и напряжение газа
Количество газа, растворенного в воде (жидкости), соответствует

Слайд 19Водное дыхание
Мелкие организмы
получают О2 только
посредством
диффузии
Более крупные имеют
специализированные
Водное дыхание
Мелкие организмы
получают О2 только
посредством
диффузии
Более крупные имеют
специализированные
Слайд 201. Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
1. Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V

Слайд 21Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R
Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R

Слайд 22Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R
Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R

Слайд 23Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R
Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R

Слайд 24Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R
Животные без специализированных органов дыхания
Сфера: минимально возможная S при данном V
R
R

Слайд 25 Вычисления для организма с R = 1мм дают разумную
величину,
Вычисления для организма с R = 1мм дают разумную
величину,
Слайд 262. Водные животные, обладающие органами дыхания
Жабры исходно были вывернуты наружу, у рыб
2. Водные животные, обладающие органами дыхания
Жабры исходно были вывернуты наружу, у рыб

Слайд 27Вентиляция жабр
Движение жабры в воде:
«практично» только для
небольших размеров R
(для личинок поденок).
Требуется
Вентиляция жабр
Движение жабры в воде:
«практично» только для
небольших размеров R
(для личинок поденок).
Требуется

Слайд 28> S жабр ? > V газообмена.
У высокоактивных и быстро
плавающих рыб
> S жабр ? > V газообмена.
У высокоактивных и быстро
плавающих рыб
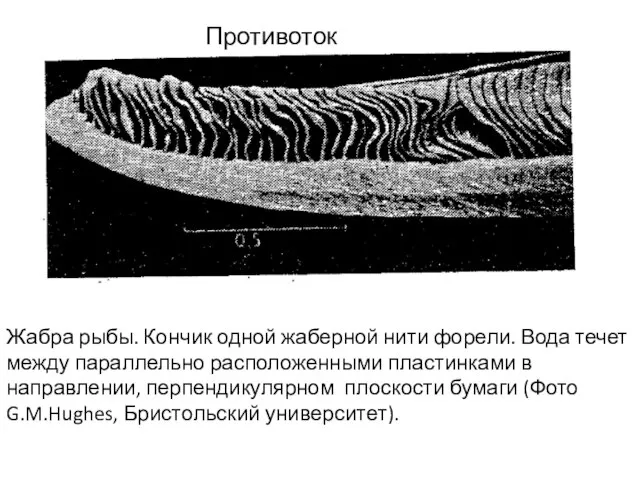
Слайд 29Жабра рыбы. Кончик одной жаберной нити форели. Вода течет
между параллельно расположенными
Жабра рыбы. Кончик одной жаберной нити форели. Вода течет
между параллельно расположенными
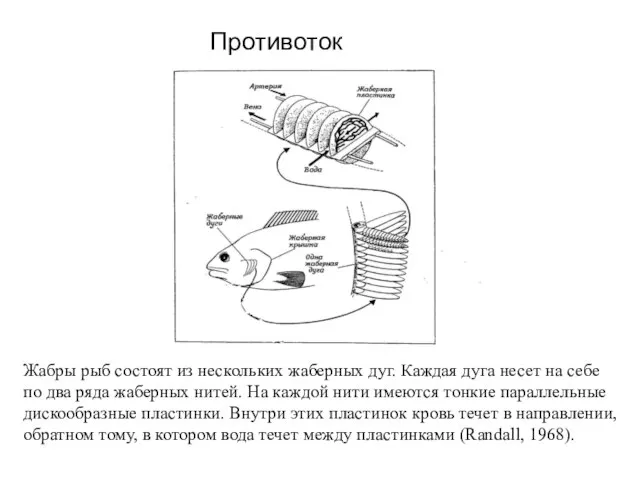
Слайд 30 Жабры рыб состоят из нескольких жаберных дуг. Каждая дуга несет на
Жабры рыб состоят из нескольких жаберных дуг. Каждая дуга несет на
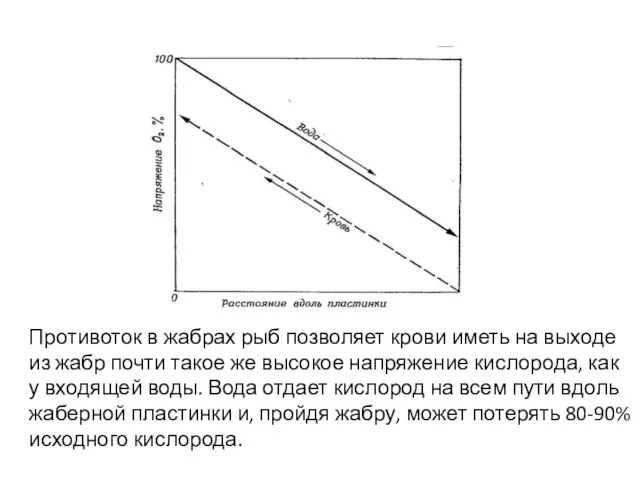
Слайд 31Противоток в жабрах рыб позволяет крови иметь на выходе из жабр почти
Противоток в жабрах рыб позволяет крови иметь на выходе из жабр почти
Слайд 32Эффективность обменных процессов между жидкостями, движущимися в
одном и том же (А) и
Эффективность обменных процессов между жидкостями, движущимися в
одном и том же (А) и

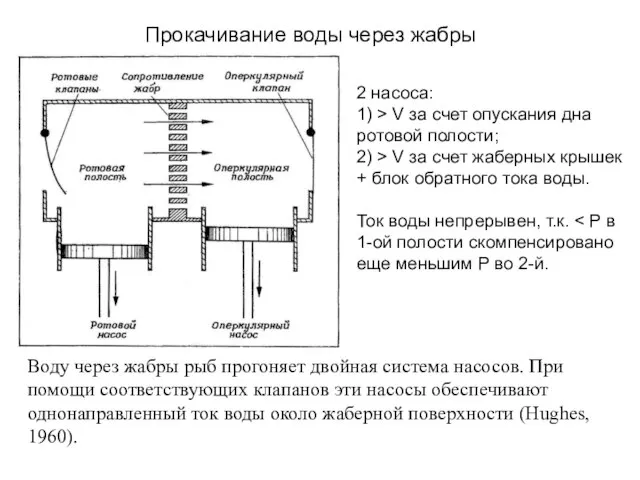
Слайд 33Воду через жабры рыб прогоняет двойная система насосов. При помощи соответствующих клапанов
Воду через жабры рыб прогоняет двойная система насосов. При помощи соответствующих клапанов
Слайд 34Когда скорость движения макрели в воде возрастает до 0,5-0,8 м/сек, оперкулярный насос
Когда скорость движения макрели в воде возрастает до 0,5-0,8 м/сек, оперкулярный насос
Слайд 35Дыхание на воздухе
В широком масштабе успешная эволюционная адаптация к воздушному дыханию произошла
Дыхание на воздухе
В широком масштабе успешная эволюционная адаптация к воздушному дыханию произошла

Слайд 36Вода: чтобы получить 0.01 г О2, надо прокачать
инертную массу Н2О, большую
Вода: чтобы получить 0.01 г О2, надо прокачать
инертную массу Н2О, большую
Слайд 37Растениям трудно получать необходимое количество СО2 из атмосферы
путем диффузии, не подвергая себя
Растениям трудно получать необходимое количество СО2 из атмосферы
путем диффузии, не подвергая себя
Слайд 38Органы дыхания на воздухе
Жабры
сухопутные крабы
равноногие раки
рыбы
Трахеи
Легкие
Диффузионные Вентиляционные
сухоп. улитки
равноногие раки
скорпионы
наземные
позвоночные
(большие размеры и
Органы дыхания на воздухе
Жабры
сухопутные крабы
равноногие раки
рыбы
Трахеи
Легкие
Диффузионные Вентиляционные
сухоп. улитки
равноногие раки
скорпионы
наземные
позвоночные
(большие размеры и
Слайд 39Вентиляция легких у позвоночных
Нагнетательный насос амфибий
Всасывающий насос
(рептилии, птицы, млекопитающие)
Дыхательный цикл
Вентиляция легких у позвоночных
Нагнетательный насос амфибий
Всасывающий насос
(рептилии, птицы, млекопитающие)
Дыхательный цикл
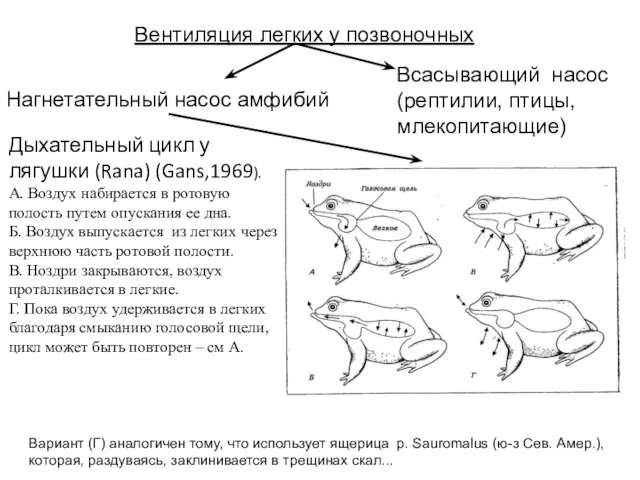
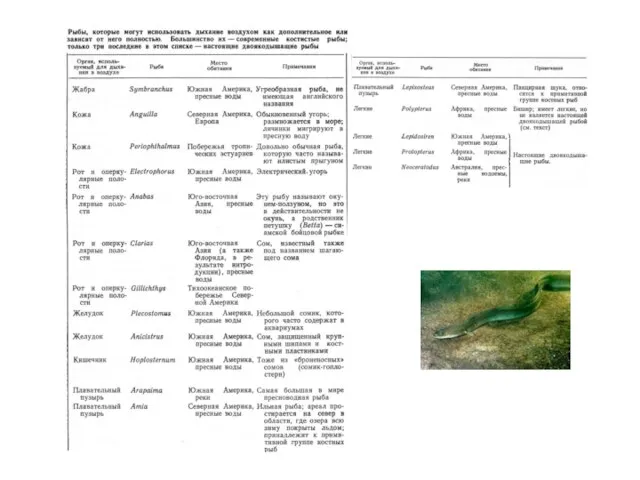
Слайд 40Рыбы, способные дышать воздухом
Помимо двоякодышащих представители 17 родов рыб
способны дышать воздухом.
Экологические
Рыбы, способные дышать воздухом
Помимо двоякодышащих представители 17 родов рыб
способны дышать воздухом.
Экологические

Слайд 41Рыба-ползун Anabas
Индийский сом
Saccobranchus
Африканский
шагающий сом
Clarias
Органы дополнительного воздушного
дыхания у рыб. Варианты васкуля-
ризации околожаберной
Рыба-ползун Anabas
Индийский сом
Saccobranchus
Африканский
шагающий сом
Clarias
Органы дополнительного воздушного
дыхания у рыб. Варианты васкуля-
ризации околожаберной
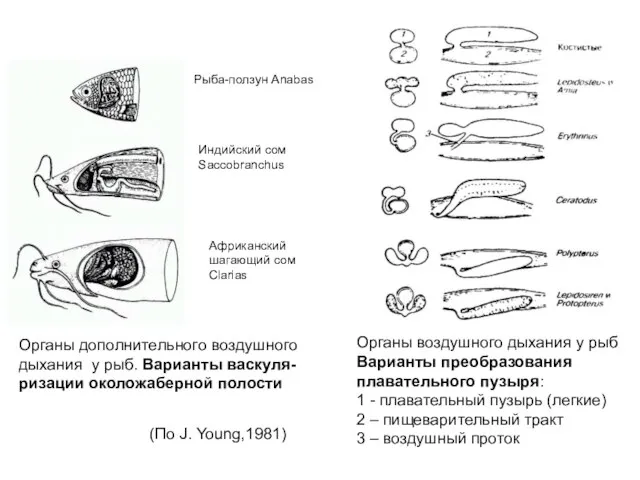
Слайд 43Рыбы, для которых дыхание воздухом обязательно
Рыбы, для которых дыхание воздухом обязательно
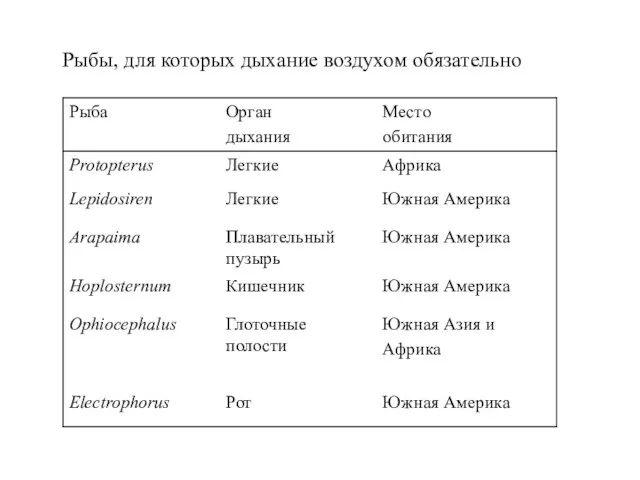
Слайд 44Относительная роль жабр и легких в дыхательном газообмене трех
видов двоякодышащих рыб при
Относительная роль жабр и легких в дыхательном газообмене трех
видов двоякодышащих рыб при
Слайд 45Легочный и кожный газообмен у жабы Bufo americanus при различных
температурах (Hutchison et
Легочный и кожный газообмен у жабы Bufo americanus при различных
температурах (Hutchison et
Слайд 46Когда африканских и южноамериканских двоякодышащих рыб вынимают
из воды, потребление кислорода у них
Когда африканских и южноамериканских двоякодышащих рыб вынимают
из воды, потребление кислорода у них
Слайд 47 Когда угря вынимают из воды (при 7оС), потребление кислорода у него
Когда угря вынимают из воды (при 7оС), потребление кислорода у него
Слайд 48График зависимости объема легких от веса тела у различных млекопитающих.
Величина легких является
График зависимости объема легких от веса тела у различных млекопитающих.
Величина легких является
Слайд 49Дыхание птиц.
Особенности строения и функционирования
легких
Дыхание птиц.
Особенности строения и функционирования
легких

Слайд 50Самыми мелкими элементами легких у млекопитающих являются
мешковидные альвеолы. В легких птиц тончайшие
Самыми мелкими элементами легких у млекопитающих являются
мешковидные альвеолы. В легких птиц тончайшие
Слайд 51Схема строения легкого птицы:
1 — бронх,
2 — вторичные бронхи,
3 —
Схема строения легкого птицы:
1 — бронх,
2 — вторичные бронхи,
3 —
Слайд 52Объем дыхательной системы у типичной птицы и типичного
млекопитающего, весом 1 кг (По
Объем дыхательной системы у типичной птицы и типичного
млекопитающего, весом 1 кг (По
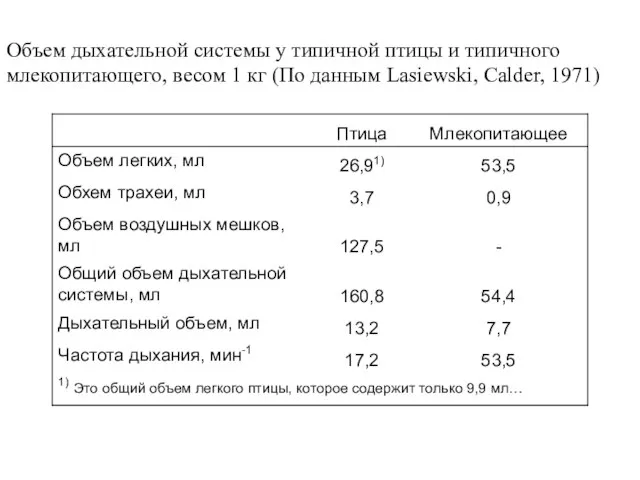
Слайд 53В теле птицы имеется несколько больших тонкостенных воздушных мешков.
Парные легкие невелики и
В теле птицы имеется несколько больших тонкостенных воздушных мешков.
Парные легкие невелики и
Слайд 54Когда страусу дают один раз вдохнуть чистый кислород, а затем он дышит
обычным
Когда страусу дают один раз вдохнуть чистый кислород, а затем он дышит
обычным

Слайд 55Движение газа, поступившего за один вздох, через дыхательную систему птицы.
Прохождение газа по
Движение газа, поступившего за один вздох, через дыхательную систему птицы.
Прохождение газа по
Слайд 56Газообмен в легком птицы на большой высоте (Schmidt-Nielsen, 1972).
На упрощенной схеме показаны
Газообмен в легком птицы на большой высоте (Schmidt-Nielsen, 1972).
На упрощенной схеме показаны

Слайд 57В легком птицы кровь течет не по параллельным капиллярам, а скорее по
сложной,
В легком птицы кровь течет не по параллельным капиллярам, а скорее по
сложной,
 Суперконтинент Пангея
Суперконтинент Пангея Презентация на тему: 1 и 2 закон Менделя Порхун Александры Группа 306 Сд 2013 год
Презентация на тему: 1 и 2 закон Менделя Порхун Александры Группа 306 Сд 2013 год Из гусеницы в бабочку. 3 класс
Из гусеницы в бабочку. 3 класс Физиология гимодинамики и ее возрастные особенности. Лекция 11
Физиология гимодинамики и ее возрастные особенности. Лекция 11 Кошки. Интересные факты
Кошки. Интересные факты Развитие коммуникативных УУД на уроках биологии и химии
Развитие коммуникативных УУД на уроках биологии и химии Возникновение адаптаций в результате естественного отбора
Возникновение адаптаций в результате естественного отбора Удивительная природа. Урок окружающего мира
Удивительная природа. Урок окружающего мира Жизнедеятельность организмов
Жизнедеятельность организмов Еловый лес
Еловый лес Area of distributijn of red wolves
Area of distributijn of red wolves Половое размножение. Эмбриональный период
Половое размножение. Эмбриональный период Виготовлення мікропрепаратів шкірки луски цибулі та розгляд її за допомогою оптичного мікроскопа
Виготовлення мікропрепаратів шкірки луски цибулі та розгляд її за допомогою оптичного мікроскопа Лекарственные растения
Лекарственные растения Генетика пола
Генетика пола Моллюски. Класс Брюхоногие
Моллюски. Класс Брюхоногие Я і навакольны свет. Надвор’е, з’явы прыроды
Я і навакольны свет. Надвор’е, з’явы прыроды Животные, имеющие суперспособности
Животные, имеющие суперспособности Бассет хаунд
Бассет хаунд Зоркий глаз
Зоркий глаз Биосинтез белка
Биосинтез белка Липиды и их производные
Липиды и их производные Введение в вирусологию
Введение в вирусологию Пищевые продукты и питательные вещества
Пищевые продукты и питательные вещества Тема: Влияние температуры на живые организмы Цели: Дать понятие абиотическому фактору, тепловому балансу Проклассифицировать ра
Тема: Влияние температуры на живые организмы Цели: Дать понятие абиотическому фактору, тепловому балансу Проклассифицировать ра Klassifikation von Viren
Klassifikation von Viren Мейоз/Meiosis/إنقسام منصف
Мейоз/Meiosis/إنقسام منصف Способности человека
Способности человека