- Terno_Olga

Содержание
- 2. ТИПЫ ПИТАНИЯ ЖИВЫХ ОРГАНИЗМОВ Жизнедеятельность всех организмов возможна только при наличии в них энергии. По способу
- 3. ФОТОСИНТЕЗ Фотосинтез(греч. phos — род. пад. photos — свет и synthesis — соединение) — образование при
- 4. СВЕТОВЫЕ РЕАКЦИИ ФОТОСИНТЕЗА Протекают в гранах тилакоидов хлоропластов .Кванты видимого света (фотоны) взаимодействуют с молекулами хлорофилла,
- 5. ТЕМНОВЫЕ РЕАКЦИИ, ИЛИ РЕАКЦИИ ФИКСАЦИИ УГЛЕРОДА Не связанные со светом, осуществляются в строме хлоропластов. Ключевое место
- 6. ХЕМОСИНТЕЗ (позднегреч. chemeta — химия и греч. synthesis — соединение) — автотрофный процесс создания органического вещества
- 7. КАТАБОЛИЗМ Катаболи́зм (от греч. καταβολή, «сбрасывание, разрушение»), также энергетический обмен, или диссимиляция, — это процесс метаболического
- 8. АНАБОЛИЗМ Анаболизм (пластический обмен) - совокупность химических процессов, составляющих одну из сторон обмена веществ в организме,
- 9. МЕТАБОЛИЗМ
- 11. Скачать презентацию
Слайд 2ТИПЫ ПИТАНИЯ ЖИВЫХ ОРГАНИЗМОВ
Жизнедеятельность всех организмов возможна только при наличии в них энергии.
ТИПЫ ПИТАНИЯ ЖИВЫХ ОРГАНИЗМОВ
Жизнедеятельность всех организмов возможна только при наличии в них энергии.

Слайд 3ФОТОСИНТЕЗ
Фотосинтез(греч. phos — род. пад. photos — свет и synthesis — соединение)
ФОТОСИНТЕЗ
Фотосинтез(греч. phos — род. пад. photos — свет и synthesis — соединение)

Слайд 4СВЕТОВЫЕ РЕАКЦИИ ФОТОСИНТЕЗА
Протекают в гранах тилакоидов хлоропластов .Кванты видимого света (фотоны) взаимодействуют
СВЕТОВЫЕ РЕАКЦИИ ФОТОСИНТЕЗА
Протекают в гранах тилакоидов хлоропластов .Кванты видимого света (фотоны) взаимодействуют

Слайд 5ТЕМНОВЫЕ РЕАКЦИИ, ИЛИ РЕАКЦИИ ФИКСАЦИИ УГЛЕРОДА
Не связанные со светом, осуществляются в строме
ТЕМНОВЫЕ РЕАКЦИИ, ИЛИ РЕАКЦИИ ФИКСАЦИИ УГЛЕРОДА
Не связанные со светом, осуществляются в строме

Слайд 6ХЕМОСИНТЕЗ
(позднегреч. chemeta — химия и греч. synthesis — соединение) — автотрофный процесс
ХЕМОСИНТЕЗ
(позднегреч. chemeta — химия и греч. synthesis — соединение) — автотрофный процесс

Слайд 7КАТАБОЛИЗМ
Катаболи́зм (от греч. καταβολή, «сбрасывание, разрушение»), также энергетический обмен, или диссимиляция, —
КАТАБОЛИЗМ
Катаболи́зм (от греч. καταβολή, «сбрасывание, разрушение»), также энергетический обмен, или диссимиляция, —

Слайд 8АНАБОЛИЗМ
Анаболизм (пластический обмен) - совокупность химических процессов, составляющих одну из сторон обмена веществ в
АНАБОЛИЗМ
Анаболизм (пластический обмен) - совокупность химических процессов, составляющих одну из сторон обмена веществ в

Слайд 9МЕТАБОЛИЗМ
МЕТАБОЛИЗМ

 Естественный отбор
Естественный отбор Обмен веществ (метаболизм)
Обмен веществ (метаболизм) Камеры, хрусталик, цилиарное тело
Камеры, хрусталик, цилиарное тело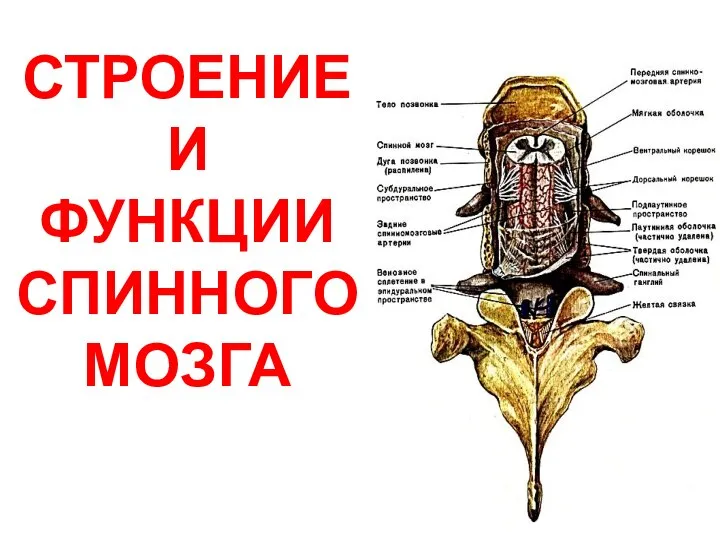 Строение и функции спинного мозга
Строение и функции спинного мозга Общие вопросы. Задания 27. Часть 2-3
Общие вопросы. Задания 27. Часть 2-3 Презентация на тему "Глаз и зрение. Близорукость и дальнозоркость" - презентации по Биологии
Презентация на тему "Глаз и зрение. Близорукость и дальнозоркость" - презентации по Биологии Организм – открытая живая система (биосистема)
Организм – открытая живая система (биосистема) 11.3 Индивидуальное развитие организмов. Биогенетический закон
11.3 Индивидуальное развитие организмов. Биогенетический закон ОРГАНОИДЫ
ОРГАНОИДЫ Первоцветы Жигулей
Первоцветы Жигулей Порушення функцій скелета та їх профілактика
Порушення функцій скелета та їх профілактика Игра-викторина. Части растений. Поверхность нашей планеты. Водоёмы
Игра-викторина. Части растений. Поверхность нашей планеты. Водоёмы Игра Юннатское движение 2 часть
Игра Юннатское движение 2 часть Связочный аппарат матки и ее придатков
Связочный аппарат матки и ее придатков Класс Амфибии, 7 класс
Класс Амфибии, 7 класс В царстве грибов
В царстве грибов Kletochnaya_teoria
Kletochnaya_teoria Размножение - свойство живых организмов
Размножение - свойство живых организмов Доклад о бегемоте
Доклад о бегемоте лекция4-5 ЦНС
лекция4-5 ЦНС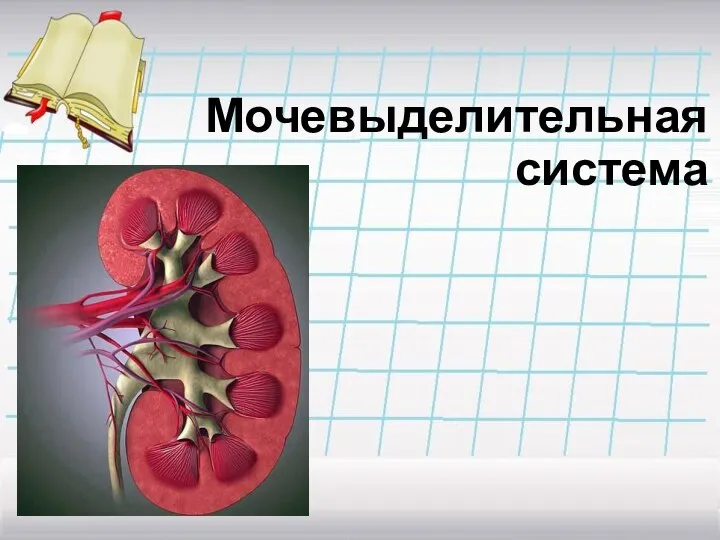 Мочевыделительная система
Мочевыделительная система Клетка и ее органоиды
Клетка и ее органоиды Строение вегетативная нервной системы человека
Строение вегетативная нервной системы человека Презентация на тему Губки. Тупиковая ветвь эволюции
Презентация на тему Губки. Тупиковая ветвь эволюции  Мутации и мутагенные факторы. Лекция № 3
Мутации и мутагенные факторы. Лекция № 3 Aýaklylar Görnüş: Aýaklylar
Aýaklylar Görnüş: Aýaklylar Гармония и Баланс
Гармония и Баланс Высшие споровые растения
Высшие споровые растения