- Травлення. Переробка і перетворення харчових продуктів

Содержание
- 2. Функції травної системи секреторна – утворення і виділення залозистими клітинами травних соків (слина, шлунковий, підшлунковий і
- 3. захисна (бактерицидна, бактеріостатична, дезінтоксикаційна) – здійснюється завдяки бар’єрним системам шлунково-кишкового тракту і рефлекторним механізмам; рецепторна (аналізаторна)
- 5. ХАРАКТЕРИСТИКА РІЗНИХ ТИПІВ ТРАВЛЕННЯ Власний: організм використовує власні ферменти для асиміляції їжі. Ділиться на позаклітинне (порожнинне),
- 6. Особливості основних типів травлення Внутрішньоклітинне. Еволюційно та філогенетично найстаріше: лізосомальні ферменти гідролізують харчові речовини, які попадають
- 7. 2) Мембранне - здійснюється ферментами, фіксованими на апікальній мембрані ентероцитів , є основним механізмом гідролізу дрібних
- 8. Рис. Схема позаклітинного (А), внутрішньоклітинного (Б) і мембранного (В) гідролізів харчових речовин. 1 — позаклітинне середовище;
- 9. Рис. Локалізація процесів порожнинного і мембранного травлення тонкої кишки та ферментів, які їх здійснюють.
- 10. Рис. Схема деполімеризації часточок харчових речовин на поверхні мікроворсинок ентероцита, апікальному та латеральному глікокаліксі. 1—3 —
- 11. Травні залози і секреція Секреція – це складний процес, під час якого секреторна клітина отримує з
- 12. Секреторний цикл - процеси, які послідовно повторюються в секреторних клітинах. Може бути виміряний в одиницях часу,
- 13. 1 фаза секреторного циклу– надходження первинних речовин Основні механізми проникнення речовин через плазматичну мембрану: дифузія, пасивний
- 14. 2 фаза секреторного циклу – синтез
- 15. Рис. Механізм спрямування поліпептиду в ендоплазматичний ретикулум: 1 – мРНК; 2 – тРНК; 3 – вільна
- 16. 3 фаза –1) транспорт секреторних продуктів Сигнальна гіпотеза транспорту синтезованих секреторних білків.
- 17. 3 фаза секреторного циклу – 2) дозрівання секрету Модифікація поліпептидів секрету в ендоплазматичному ретикулумі і в
- 18. 5 фаза секреторного процесу - виведення (екструзія) секрету з клітин Сегрегаційна теорія
- 19. Cекреторні клітини за типом виділення секрету: голокринові (клітини епітелію шлунка), які характеризуються тим, що вся клітина
- 20. Рис. Типи виділення секрету травними клітинами: 1 – голокриновий; 2 – мікро- і макроапокриновий тип (а,б);
- 21. Речовини, які поступають в клітину, використовуються як вихідний матеріал для секреторного продукту і для забезпечення обміну
- 22. Секретовані речовини виводяться з клітини за конститутивним та регульованим механізмами (молекули зберігаються у секреторних пухирцях, які
- 23. 5 етапів регульованого екзоцитозу: 1) спряження стимул-секреція – це послідовність процесів, що викликаються дією стимула на
- 24. Захисні механізми травного тракту Органи травного тракту виконують захисну функцію завдяки центральним рефлекторним механізмам і місцевим
- 25. Нудота – неприємне, суб’єктивне відчуття, формується в ЦНС при значному підвищенні збудження нейронів ретикулярної формації, викликане
- 26. Форми нудоти: 1) мозкова; 2) рефлекторна виникає під час механічного або токсичного подразнення кореня язика, тильної
- 27. Блювота – це захисна складнокоординована системна рефлекторна реакція. Вона виникає при збудженні центру блювоти, локалізованого в
- 28. Аферентні впливи. На блювотний рефлекс впливають вищі відділи стовбуру мозку і кора. Окремі запахи, смакові відчуття,
- 29. Ефекторні впливи по доцентровим волокнам в складі блукаючого і черевного нервів передаються до кишечника, шлунка, стравоходу,
- 30. Акту блювоти передує виникнення нудоти і антиперистальтики. Антиперистальтика в кишечнику фізіологічно обумовлена надмірними подразненнями або розтягненням
- 31. НЕСПЕЦИФІЧНІ МЕХАНІЗМИ ЗАХИСТУ З харчовими речовинами в травну систему попадають різноманітні шкідливі речовини (токсини, канцерогени, солі
- 32. нормальна рухова і секреторна активність тонкого кишечника. Постійна секреція рідини в криптах ворсинок вимиває бактерії в
- 33. Захисні механізми ентероцита: ендоплазматична сітка, комплекс Гольджі, в яких частина харчових речовин піддається метаболічним перетворенням і
- 34. Механічна або пасивна система захисної системи ШКТ: обмеження проникності слизової оболонки для водорозчинних молекул, молекулярна маса
- 35. СПЕЦИФІЧНІ ІМУННІ МЕХАНІЗМИ ЗАХИСТУ Специфічні імунні механізми забезпечуються імунокомпетентною “лімфоїдною тканиною”: інтраепітеліальні Т і В-лімфоцити, які
- 36. Вплив імуноглобулінів: антивірусний; антитоксичний; протимікробний; місцевий імунітет; антипаразитарний. Первинна імунна нейтралізація антигенів відбувається в пристінковому шарі
- 37. SIgA складається з підкласів SIgA1 (25-30%), SIgA2 (60-70%), SIgA – мономер (5%). В шлунковому соці химерний
- 38. Недиференційовані лімфоцити, розташовані у власному шарі або пейєрових бляшках продукують секреторні антитіла Ig A і Ig
- 39. Імунна система організму, кишечні гідролітичні ферменти і система тканинної детоксикації (апарат окислювального гідроксилювання і лізосомальну систему
- 40. ТРАВЛЕННЯ В РОТОВІЙ ПОРОЖНИНІ
- 41. Ротова порожнина, глотка і стравохід - функціональний комплекс, який забезпечує первинну обробку їжі. Цей комплекс забезпечує:
- 42. СКЛАД І ВЛАСТИВОСТІ СЛИНИ Слина – сукупний зовнішньо-секреторний продукт привушних, підщелепних, під’язикових великих слинних залоз і
- 43. Сухий залишок містить: неорганічні та органічні речовини (›50%).Неорганічні компоненти: Na+, K+, Ca2+, Fe2+, Li+, Mg2+ .
- 44. РЕГУЛЯЦІЯ СЛИНОВИДІЛЕННЯ Секреторна функція слинних залоз і слиновиділення регулюються рефлекторно. Розрізняють безумовно- і условнорефлекторні впливи на
- 45. До слинних залоз імпульси передаються парасимпатичними і симпатичними нервовими волокнами. Парасимпатична іннервація привушних слинних залоз здійснюється
- 46. Симпатична іннервація слинних залоз здійснюється волокнами, що відходять від верхніх шийних симпатичних гангліїв. Подразнення цих волокон
- 47. Умовно-рефлекторне слиновиділення викликають вид і запах їжі, а також звуки, пов'язані з годівлею. Умовно-рефлекторне слиновиділення у
- 48. МЕХАНІЧНІ ПРОЦЕСИ: ЖУВАННЯ І КОВТАННЯ Жування — це процес механічної обробки їжі, що полягає в її
- 49. У людини і більшості тварин верхня щелепа нерухома, тому жування здійснюється рухами нижньої щелепи зверху вниз
- 50. Жування відбувається мимоволі, однак включає і довільні компоненти. Регуляція здійснюється на різних рівнях ЦНС, у тому
- 51. Ковтання — складний рефлекторний акт, у процесі якого їжа переходить з ротової порожнини в стравохід, а
- 52. У глоткову фазу: харчова грудка на корені язика подразнює чутливі нервові закінчення трійничних, верхніх і нижніх
- 53. Стравохідна фаза: потрапивши в стравохід, харчову грудку проходить в шлунок, що складає стравохідну фазу ковтання і
- 54. Перистальтика стравоходу Рухи стравоходу взаємопов’язані з рухами ковтального апарату і викликаються рефлекторно при кожному ковтальному акті.
- 55. Швидкість поширення перистальтичної хвилі залежить від консистенції харчової грудки і положення тіла. При вертикальному положенні тіла
- 56. ПОРУШЕННЯ НОРМАЛЬНОЇ ФУНКЦІЇ СТРАВОХОДУ При ахалазії (спазм кардіального відділу стравоходу і розширення його відділів, розташованих вище)
- 57. Скорочення м'язів стравоходу пов'язані зі збудженням холінергічних нейронів. Їх розслаблення не обумовлене ні холінергічним, ні адренергічним
- 58. ТРАВЛЕННЯ В ШЛУНКУ
- 59. Шлунок – це відділ травної системи, в якому накопичується проковтнута їжа. Вона змішана зі слиною і
- 60. ФУНКЦІЇ ШЛУНКУ: Депонування їжі: в проксимальному відділі шлунка відсутній ритм збудження і перистальтики. В залежності від
- 61. Секреторна функція: виділення соку шлунковими залозами. Сік містить іони і макромолекули, забезпечує хімічну обробку їжі. Кардіальний
- 62. Рухова функція: перемішування їжі з шлунковим соком і її просування в дванадцятипалу кишку. В тілі шлунка
- 63. Інкреторна: утворення біологічно активних речовин (гормонів): гастрину, гістаміну, соматостатину, мотиліну й інших речовин, що беруть участь
- 64. СКЛАД І ВЛАСТИВОСТІ ШЛУНКОВОГО СОКУ Шлунковий сік — продукт зовнішньо секреторної і екскреторної діяльності залоз шлунка.
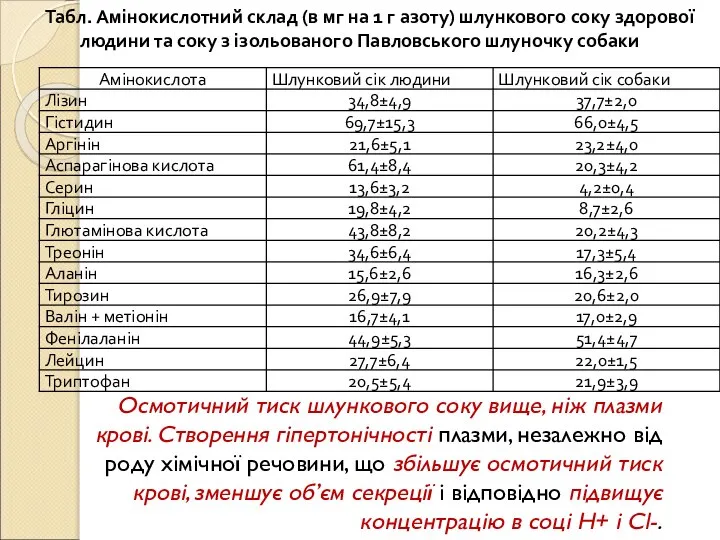
- 65. Неорганічні речовини: хлориди, сульфати, фосфати, бікарбонати натрію, калію, кальцію, магнію. Соляна кислота майже вся дисоційована і
- 66. Табл. Амінокислотний склад (в мг на 1 г азоту) шлункового соку здорової людини та соку з
- 67. Ферменти шлункового соку. Виявлено щонайменше 8 протеолітичних ферментів. 5 швидко мігруючих пепсиногенів складають групу I і
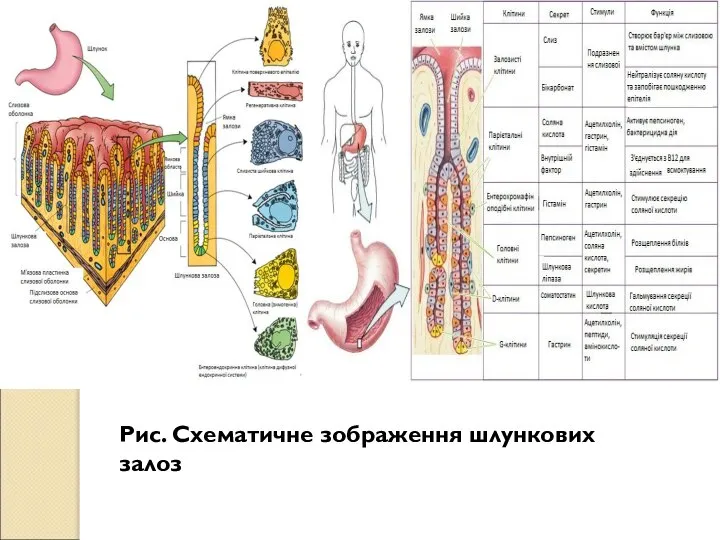
- 68. Важливий компонент шлункового соку — слиз, що захищає слизову від механічного і хімічного ушкодження. Основною складовою
- 69. Рис. Схематичне зображення шлункових залоз
- 70. РЕГУЛЯЦІЯ ШЛУНКОВОЇ СЕКРЕЦІЇ Переривчастість або безперервність шлункової секреції залежить від умов харчування, що властиві різним тваринам.
- 71. Шлункова секреція регулюється комплексом ендокринних, паракринних, нейрональних і місцевих механізмів (парасимпатических, симпатичних, метасимпатических рефлекторних дуг). На
- 72. ПЕРША ЦЕФАЛІЧНА (СКЛАДНОРЕФЛЕКТОРНА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ
- 73. Важливу роль у нейрогенній стимуляції секреторних процесів відіграє гастрин. Видалення пілоричного відділу шлунка у собак викликало
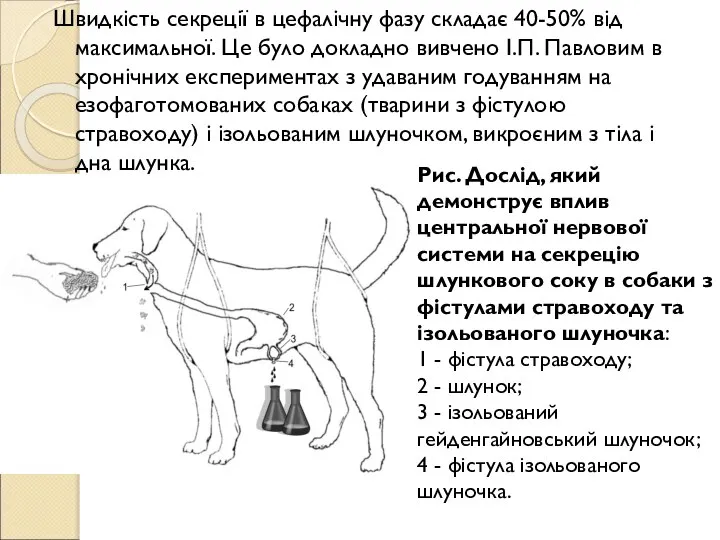
- 74. Швидкість секреції в цефалічну фазу складає 40-50% від максимальної. Це було докладно вивчено І.П. Павловим в
- 75. Удаване годування, що викликає збудження блукаючих нервів, здійснює як гальмівний, так і стимулюючий вплив на вагально
- 76. Усі гальмівні нейропептиди, окрім бомбезина, справляють свій вплив на шлункову секрецію через систему блукаючих нервів (через
- 77. ДРУГА (НЕЙРОГУМОРАЛЬНА) ШЛУНКОВА ФАЗА СЕКРЕЦІЇ
- 78. Фактори регуляції шлункової секреції у другу (нейрогуморальну) фазу
- 79. ЗАХИСНІ МЕХАНІЗМИ ДЛЯ СЕКРЕТОРНОГО АПАРАТУ ШЛУНКА
- 80. ВПЛИВ РОЗТЯГНЕННЯ ШЛУНКУ НА СЕКРЕЦІЮ ШЛУНКОВОГО СОКУ
- 81. Участь капсаїцин-чутливої системи у регуляції шлункової секреції
- 82. ШЛУНКОВА ФАЗА ГАЛЬМУВАННЯ Це гальмування звільнення гастрина при омиванні антральної слизової кислим розчином. Закислення антральної слизової
- 83. Дія соматостатину: гальмує виділення гормонів гіпофіза; гальмує виділення інсуліну і глюкагона, стимульоване усіма видами їжі; зменшує
- 84. Шлунковий соматостатин – паракринний гальмівний регулятор секреції кислоти в шлунку. Виділяється в присутності кислоти у фундальному
- 85. ІІІ-ТЯ (КИШКОВА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ Ентеро-гастральний рефлекс
- 86. Гальмування шлункової секреції продовжується під дією секретину, що звільняється S-клітинами слизової оболонки проксимального відділу дванадцятипалої кишки
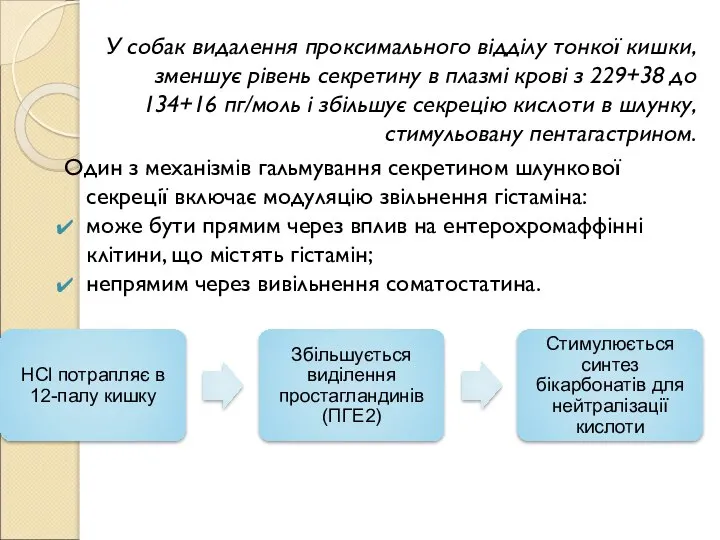
- 87. У собак видалення проксимального відділу тонкої кишки, зменшує рівень секретину в плазмі крові з 229+38 до
- 88. Гальмівний вплив секретину не залежить від вагальної іннервації фундального відділу шлунка. Однак, гальмування секретином секреції кислоти,
- 89. Жир, введений у тонкий кишечник, гальмує секрецію кислоти в шлунку. Механізм гальмування жиром шлункової секреції включає:
- 90. До ентерогастронів відносять: гастроінгібуючий поліпептид, секретин, глюкагоно-подібні пептиди 1 і 2, пептид YY, холецистокінін, нейротензин, аполіпротеїн
- 91. ШЛУНКОВИЙ СЛИЗ: УТВОРЕННЯ І ВИДІЛЕННЯ Слиз – складна динамічна система колоїдних розчинів високомолекулярних біополімерів, що включає
- 92. До складу слизу входять нейтральні мукополісахариди, сіаломуцини, глікопротеїди і протеоглікани (кислі мукополісахариди). Слиз покриває всю внутрішню
- 93. При значеннях рН між 7,0 і 7,9 шлунковий слиз має мінімальну в'язкість; по мірі зниження рН
- 94. ОСНОВНІ ЕТАПИ БІОСИНТЕЗУ І СЕКРЕЦІЇ СЛИЗУ
- 95. Шлунковий слиз утворюється в результаті взаємодії мукоїдних речовин, що надходять у просвіт шлунка, з іншими компонентами
- 96. Мукоцити одночасно з муцином продукують бікарбонат. Муцин взаємодіє з бікарбонатом, в результаті чого утворюється мукозо-бікарбонатний бар'єр,
- 97. На підставі гістохімічного аналізу виділено чотири основних класи біополімерів, що утворюють нерозчинний і розчинний слиз: 1.
- 98. 3. Сіаломуцини беруть участь у синтезі соляної кислоти. Сиалові сульфоглікопротеїни синтезуються поверхневим епітелієм тільки у м'ясоїдних.
- 99. Шлунковий слиз, бікарбонати, фосфоліпіди і вода утворюють гель, що створює фізико-хімічний бар'єр для епітеліальних клітин шлунку.
- 100. МЕХАНІЗМИ РЕГУЛЯЦІЇ ВИДІЛЕННЯ СЛИЗУ Фактори, які стимулюють синтез пепсину і соляної кислоти, одночасно стимулюють і виділення
- 101. МЕХАНІЗМ ВПЛИВУ ІНСУЛІНУ НА СЕКРЕЦІЮ ТА ВИДІЛЕННЯ ШЛУНКОВОГО СЛИЗУ
- 102. Інсулін стимулює також секрецію внутрішнього фактора Кастла за механізмом, аналогічним стимуляції секреції пепсину: в результаті прямого
- 103. Місцеве механічне подразнення значно стимулює секрецію слизу мукоїдними клітинами антрального і фундального відділів шлунку. Це пов'язано
- 104. ГОРМОНАЛЬНІ ФАКТОРИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СЛИЗУ Істотну роль відіграють гормони гіпофізарно-адреналової системи. АКТГ стимулює інкрецію глюкокортикоїдів, що
- 105. Серотонін (5-гідрокситриптамін) є однією з ланок гормональної регуляції секреції мукоїдних речовин. Він стимулює секрецію слизу в
- 106. Участь місцевого подразнення слизової оболонки шлунку Механічне і хімічне (гормони, нейротропні речовини, ліки) подразнення: етиловий спирт,
- 107. МЕХАНІЗМИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СОЛЯНОЇ КИСЛОТИ І ЇЇ РОЛЬ В ТРАВЛЕННІ Обкладові клітини мають унікальну здатність до
- 108. БУДОВА ОБКЛАДОВИХ КЛІТИН Обкладові клітини містять численні великі мітохондрії, характерні внутрішньоклітинні канальці, вистелені мікроворсинками, що відкриваються
- 109. Рис. Обкладова клітина в спокої (А) і при стимуляції (Б): Я – ядро, Р – гладкий
- 110. В мікроворсинках канальців локалізована H+,K+-АТФаза, що транспортує іони Н+. Обкладові клітини утворюють соляну кислоту за участю
- 111. Рис. Секреція НСІ обкладовою клітиною
- 112. В секреторній фазі секретуєтся більше іонів Cl-, ніж іонів Н+. У стані спокою секреція іонів Сl-
- 113. Активація секреторного процесу в обкладових клітинах є результатом інтеграції нейрогуморальних механізмів. Медіатори-первинні посередники в індукції секреції:
- 114. Гуморальні чинники регуляції шлункової секреції На обкладовій клітині знаходиться три види рецепторів, збудження яких стимулює секрецію
- 115. Слизова оболонка містить піридоксальзалежний фермент гістидиндекарбоксилазу, необхідний для синтезу гістамину. Гістидиндекарбоксилаза активується гастрином. Вихід з клітин
- 116. У шлунку ссавців виділяють два депо гістаміну: гладкі і ентерохромафіно-подібні (ECL) клітини. Крім того, він міститься
- 117. В нормальних регуляторних процесах бере участь гістамін ЕСL-клітин. ECL-клітини — це маленькі аргірофільні клітини, у великій
- 118. Рецептори, що гальмують активність на ECL-клітинах: SST2 чутливі до соматостатину CGRP, чутливі до кальцитонін ген-зв'язаного пептиду
- 119. Гістамін опосередковує свою дію в організмі через взаємодію з гістаміновими рецепторами Н1–, Н2–, Н3– і Н4–
- 120. Н2-блокатори гальмують стимульовану гістаміном секрецію кислоти в шлунку людини, собак, кішок, щурів, мурчаків, жаб. Н1-рецептори не
- 121. В міжхарчову фазу секреторного процесу гістамін зберігається, головним чином, в секреторних гранулах ECL-клітин, однак невелика кількість
- 122. Ацетилхолін виділяється з терміналей постгангліонарних холінергічних нейронів і впливає на обкладові клітини прямо й опосредковано. Пряма
- 123. Непрямий вплив ацетилхоліну реалізується через його дію на ECL- клітини і G- клітини (гастриноцити), що призводить
- 124. Феномен потенціації Ефект дії між гістаміном і гастрином, між гістаміном і ацетилхоліном, згідно з яким відповідь
- 125. Роль соляної кислоти у травленні: – сприяє перетворенню пепсиногену в пепсин шляхом дисоціації інгібуючого білкового комплексу;
- 126. Патології, викликані порушеннями секреції соляної кислоти Гіперсекреція соляної кислоти є однією з важливих ланок патогенезу виразкової
- 127. РУХОВА АКТИВНІСТЬ ШЛУНКА І ЕВАКУАЦІЯ ЙОГО ВМІСТУ Шлунок виконує важливу транспортну функцію: накопичує, перетирає і переміщує
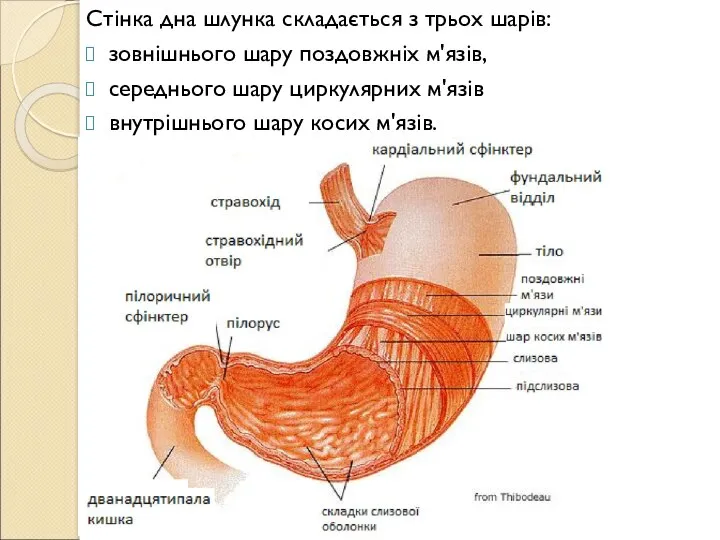
- 128. Стінка дна шлунка складається з трьох шарів: зовнішнього шару поздовжніх м'язів, середнього шару циркулярних м'язів внутрішнього
- 129. Збудливими медіаторами неадренергічних, нехолінергічних нейронів є вазоактивный інтестинальний пептид (ВІП), оксид азоту (NO) і аденозинтрифосфат (АТФ).
- 130. Властивість шлунка зберігати постійний рівень внутрішньопорожнинного тиску, незважаючи на прогресивне збільшення або зменшення об’єму шлунка називається
- 131. На відміну від пластичного тонусу, тонічні хвилі є фазними скороченнями, сила і тривалість яких варіюють у
- 132. Рис. Пейсмекерні потенціали шлунка (повільні хвилі): 1 – 3 скорочення за хвилину, хвилі, які реєструються електрогастрографічним
- 133. Пейсмекерні потенціали: контролюють в шлунку кількість скорочень за хвилину; впливають на поріг скорочень циркулярних м'язів: не
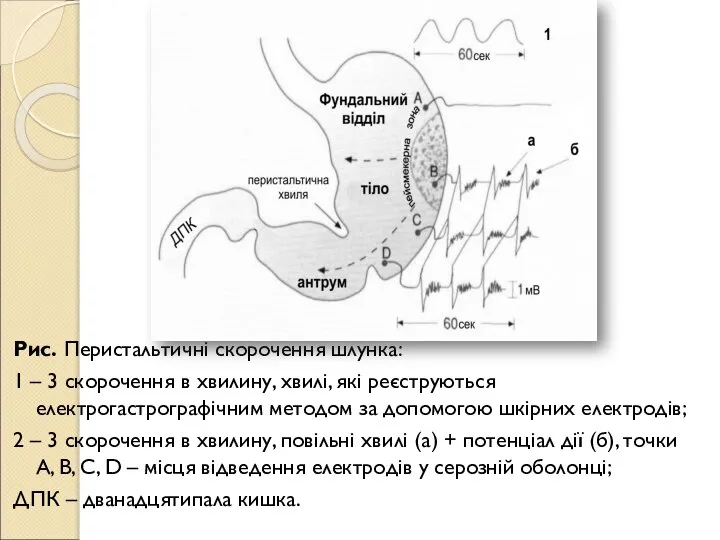
- 134. Рис. Перистальтичні скорочення шлунка: 1 – 3 скорочення в хвилину, хвилі, які реєструються електрогастрографічним методом за
- 135. Скорочення виникає в результаті надходження імпульсів в антрум, але не кожна хвиля деполяризації призводить до скорочення.
- 136. Частота появи перистальтичної хвилі у шлунку відносно стала серед представників одного виду. Для виникнення перистальтики вирішальне
- 137. Внутрішньопорожнинний тиск у пілоричному відділі шлунка в стані спокою в голодних собак коливається від +3 до
- 138. Перистальтичні хвилі виникають і передаються убік пілоруса. Пілорус є воротами шлунка і працює як м'язове кільце,
- 139. Коли хімус готовий до евакуації, кожна перистальтична хвиля евакуює 3-4 мол химуса в дванадцятипалу кишку. Евакуація
- 140. Концепція, що пояснює, чому ефективному стисканню піддаються лише великі частки їжі: вони мають більшу інерцію, ніж
- 141. Рідини евакуюються зі шлунка значно швидше, ніж тверда їжа, яка звичайно залишається в шлунку доти, поки
- 142. У багатьох роботах, присвячених фізіології шлунка, показано, що скорочення дна шлунка обумовлює евакуацію зі шлунка рідин,
- 143. Довгий час наявність градієнта тиску між шлунком і дванадцятипалою кишкою, що є причиною евакуації рідин зі
- 144. На швидкість евакуації рідин впливає їхня кількість і хімічний склад. Збільшення усередині шлунка рідкого вмісту приводить
- 145. Уповільнення евакуації зі шлунка не завжди здатне вберегти від потрапляння в дванадцятипалу кишку надлишкових на даний
- 146. Структура і функції рецепторів кишки, зв'язаних з евакуаторною функцією шлунка складні. Існують спеціальні рецептори для глюкози,
- 147. Під час відсутності імпульсів від блукаючого нерва (наприклад, при його перерізанні) перистальтика шлунка значно слабшає і
- 148. Уповільнення евакуації їжі зі шлунка називається гастропарезом. Він може виникнути внаслідок порушення нейрогуморальної регуляції, патології гладенької
- 149. ТРАВЛЕННЯ В ТОНКОМУ КИШЕЧНИКУ Ферментативне перетравлення їжі (корму) завершується в тонкому кишечнику. Всі білки, жири і
- 150. СЕКРЕЦІЯ ПІДШЛУНКОВОЇ ЗАЛОЗИ Підшлункова залоза синтезує і секретує у дванадцятипалу кишку більш 25 травних ферментів і
- 151. Характеристика панкреатичного соку: не має запаху, ізоосмотичний плазмі крові, його питома вага ~ 1,008, Неорганічні компоненти:
- 152. Основні катіони підшлункового соку: Na+, концентрація якого в соці відрізняється від концентрації в плазмі крові. Натрій
- 153. В основі секреції електролітів лежить два активних транспортних механізми: пов'язаний з транспортом натрію — натрій-калієвий насос,
- 154. Підшлункова залоза синтезує і виділяє багато ферментів, що беруть участь у розкладанні полісахаридів (амілаза), білків (трипсин,
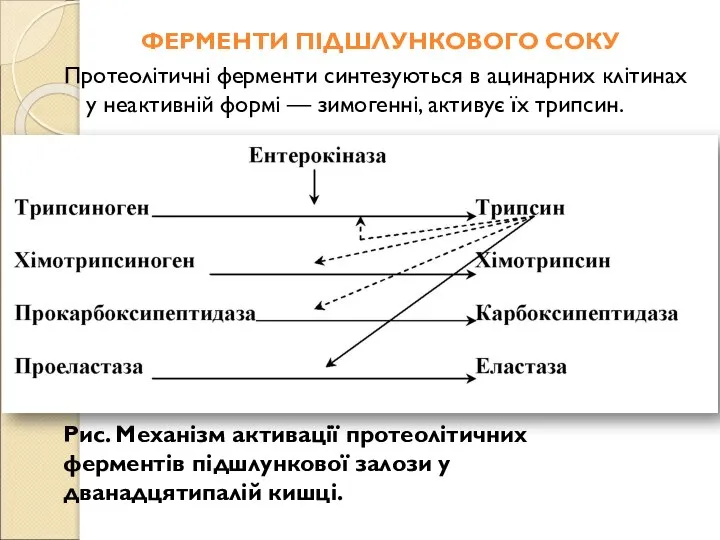
- 155. ФЕРМЕНТИ ПІДШЛУНКОВОГО СОКУ Протеолітичні ферменти синтезуються в ацинарних клітинах у неактивній формі — зимогенні, активує їх
- 156. Процеси активації є високоспецифічними й в основному складаються з розщеплення пептидного зв'язку у визначеному місці поліпептиду.
- 157. Еластаза (панкреатопептидаза) має меншу специфічність, ніж два попередні ферменти. Kарбоксипептидази гідролізують С-кінцеві амінокислотні залишки в молекулах
- 158. Один з цих білків— інгібітор Kазаля — активний тільки стосовно трипсину. Він локалізований в ацинарних клітинах
- 159. РЕГУЛЯЦІЯ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ Регуляція поживними речовинами Підшлункова залоза людини і тварин бере участь у періодичному
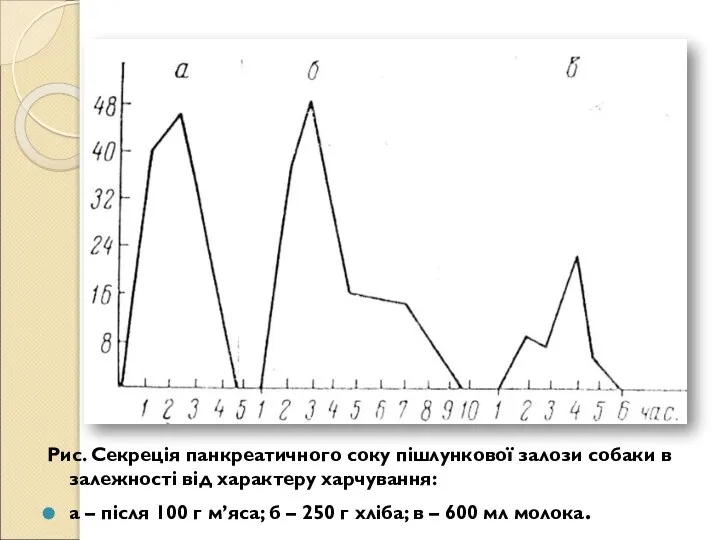
- 160. Рис. Секреція панкреатичного соку пішлункової залози собаки в залежності від характеру харчування: а – після 100
- 161. Склад ферментів соку також залежить від складу їжі. У соці, виділеному на жири, концентрація ліпаз у
- 162. Виробляється така реакція (зміни ферментативного складу соку) на харчові речовини протягом індивідуального життя тварини по типу
- 163. Ферментативна адаптація підшлункової залози починає розвиватися з перших днів змін в харчуванні тварини. Повного розвитку такі
- 164. У собак здатність підшлункової залози адаптуватися до характеру їжі різко знижується при недостатній кількості білку в
- 165. Таким чином, зміна секреції ферментів відповідно до вмісту харчових речовин у кормі забезпечує найбільш повне їхнє
- 166. НЕРВОВИЙ КОНТРОЛЬ ПАНКРЕАТИЧНОЇ СЕКРЕЦІЇ Більш 100 років тому І.П.Павлов вперше встановив, що блукаючі нерви регулюють секрецію
- 167. Антихолінергичні речовини (атропін, бантін, пробантін, ізопропамід) справляють гальмівний ефект на секрецію ферментів на стадії трансляції в
- 168. Наявність першої фази секреції підшлункової залози свідчить про участь вищих відділів ЦНС у регуляції панкреатичної секреції.
- 170. Шлункова фаза секреції підшлункової залози починається при надходженні їжі в шлунок, має нервові і гуморальні компоненти.
- 171. У кишковій фазі секреції нейрогуморальні впливи на підшлункову залозу сприяють більш інтенсивній секреції ферментів, ніж в
- 172. Ваготомія й атропін знижують секрецію ферментів, стимульовану невеликим навантаженням амінокислотами і жирними кислотами. Значне ж навантаження
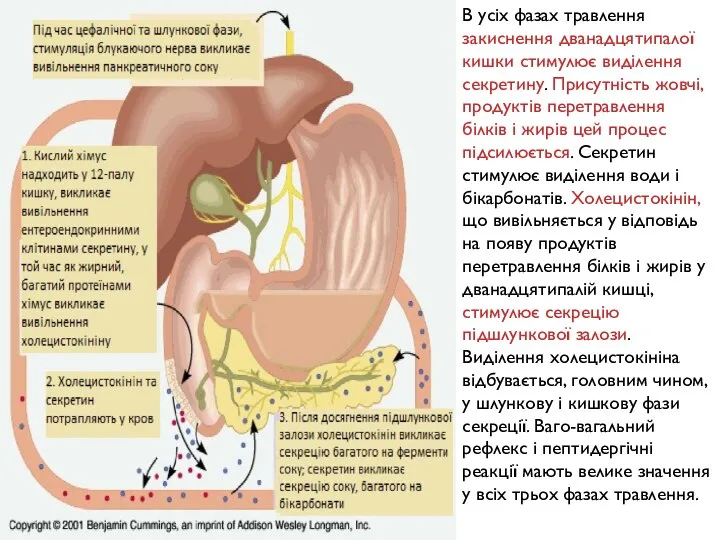
- 173. В усіх фазах травлення закиснення дванадцятипалої кишки стимулює виділення секретину. Присутність жовчі, продуктів перетравлення білків і
- 174. ГУМОРАЛЬНІ МЕХАНІЗМИ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ Гуморальні механізми забезпечують функціональну єдність усіх компонентів гастропанкреодуоденальної зони. Гуморальна регуляція
- 175. Основна маса ендокринних клітин підшлункової залози зосереджена в острівцях Лангерганса. Частина таких клітин розсіяна по паренхімі
- 176. Однак на підшлункову залозу впливають гормони й інших ендокринних клітин й органів. Це гормони гіпофіза, щитовидної
- 177. Секретин виділяється S-клітинами слизової оболонки тонкого кишечнику і стимулює клітини, що вистилають протоки і секретують головним
- 178. Тироліберин-рилізинг-фактор гіпоталамуса гальмує секрецію підшлункової залози. У ранньому неонатальному періоді його в підшлунковій залозі більше, ніж
- 179. Соматостатин також проявляє різнонаправлені ефекти на підшлункову залозу, можливо, тому, що соматостатинсекретуючі нейрони мають численні контакти
- 180. Соматостатин діє на ЦНС (гальмує нейронну активність і виділення ацетилхоліну, є агоністом опіатних рецепторів), на функцію
- 181. Енкефаліни мають широкий спектр дії і, крім участі в больовій чутливості і регуляції секреції шлунка, є
- 182. Введення лей-енкефаліна в периферичний кровотік знижує кількість панкреатичного соку в два рази. Мікроін'єкція в хвостате ядро
- 183. Бомбезин, що знаходиться в нервовій тканині, в епітеліальних клітинах кишки і шлунка, в ендокринних клітинах шлунка
- 184. Гастрин за структурою близький до холецистокініну, але має більш слабку дію на секрецію підшлункової залози. Гастрин,
- 185. Панкреатичний поліпептид (ПП) виявлений у ендо- і екзокринних клітинах підшлункової залози і, як багато регуляторних пептидів,
- 186. Ацетилхолін. Механізм цієї активації — звільнення іонів кальцію з ендоплазматичного ретикулума з наступним екзоцитозом ферментів. Однак
- 187. У механізмах регуляції секреції підшлункової залози важливе місце належить неорганічним іонам, у першу чергу іонам кальцію.
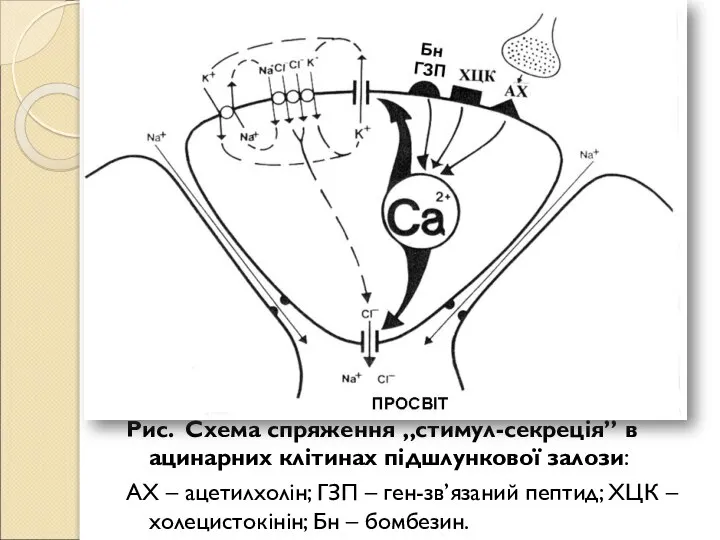
- 188. Рис. Схема спряження „стимул-секреція” в ацинарних клітинах підшлункової залози: АХ – ацетилхолін; ГЗП – ген-зв’язаний пептид;
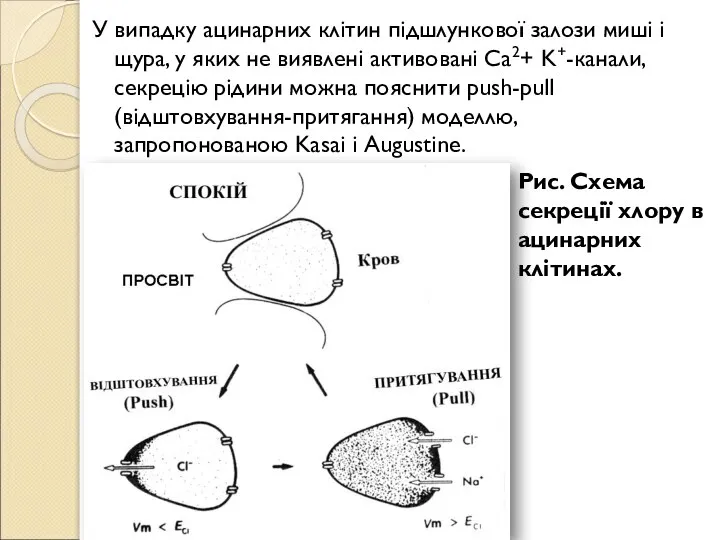
- 189. У випадку ацинарних клітин підшлункової залози миші і щура, у яких не виявлені активовані Ca2+ K+-канали,
- 190. ВЗАЄМОДІЯ ЕКЗО- І ЕНДОКРИННИХ ФУНКЦІЙ ПІДШЛУНКОВОЇ ЗАЛОЗИ Перше зазначення існування екзо- і ендокринних функцій підшлункової залози
- 191. Також, ендокринна функція підшлункової залози залежить від її зовнішньосекреторної активності. Про це свідчить той факт, що
- 192. ЖОВЧОУТВОРЮВАЛЬНА ФУНКЦІЯ ПЕЧІНКИ Печінка виконує в організмі важливі функції, серед яких: участь у травленні; кровотворення в
- 193. Бар'єрна функція печінки складається в детоксикації (знешкодженні) продуктів обміну (наприклад, продуктів розпаду білків з утворенням сечовини),
- 194. СКЛАД, ВЛАСТИВОСТІ Й УТВОРЕННЯ ЖОВЧІ Таблиця. Склад, pH та питома вага печінкової та міхурової жовчі
- 195. Через присутність білірубіна (продукт розпаду гемоглобіну) жовч має золотостий колір. Склад електролітів жовчі приблизно такий же,
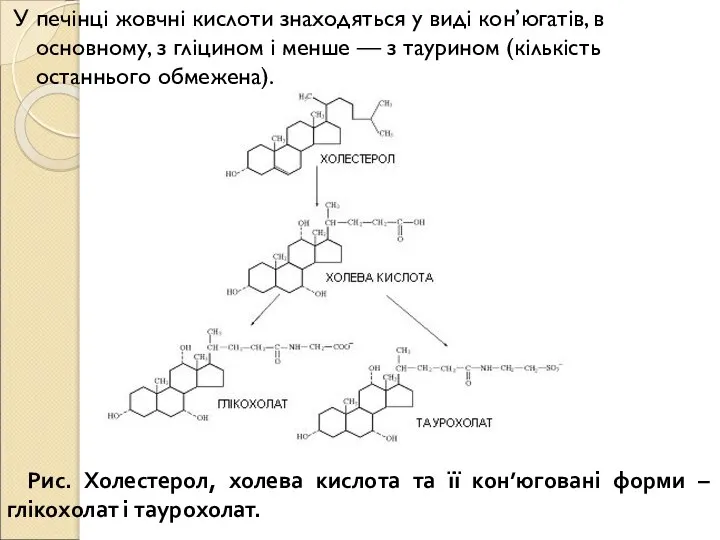
- 196. У печінці жовчні кислоти знаходяться у виді кон’югатів, в основному, з гліцином і менше — з
- 197. Завдяки амфіпатичним властивостям, жовчні кислоти, їхні солі і кон’югати є ефективними детергентами і емульгаторами. Залишки гліцину
- 198. Гідрофільні (гідроксильні і карбоксильні) групи молекул жовчних кислот знаходяться на одній стороні молекули, а гідрофобна частина
- 199. Міцели часто містять і холестерол, і фосфоліпіди (змішані міцели). Якщо концентрація холестерола перевищує ємність міцел (або
- 200. За добу печінка дорослої людини секретує 600–1200 мл (зі швидкістю близько 0,4-0,8 мл/хв) жовчі. Середня інтенсивність
- 201. У жовчі людини, собаки і кішки переважає оранжево-коричневий пігмент білірубін; у жовчі травоїдних і птахів —
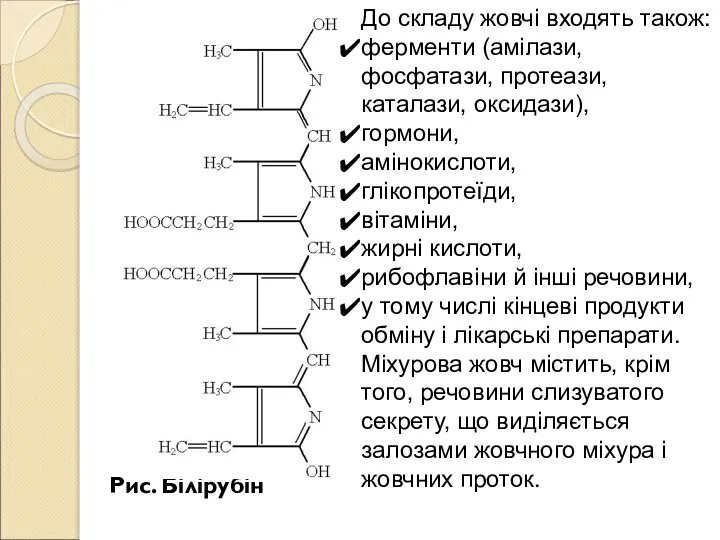
- 202. Рис. Білірубін До складу жовчі входять також: ферменти (амілази, фосфатази, протеази, каталази, оксидази), гормони, амінокислоти, глікопротеїди,
- 203. СЕКРЕЦІЯ І ФУНКЦІЇ ЖОВЧІ Близько 70% добової жовчі надходить з канальців печінки, а близько 30% —
- 205. Близько 10% (іноді до 20%) жовчних кислот виводиться з організму. Жовчні кислоти відіграють у функціонуванні кишечнику
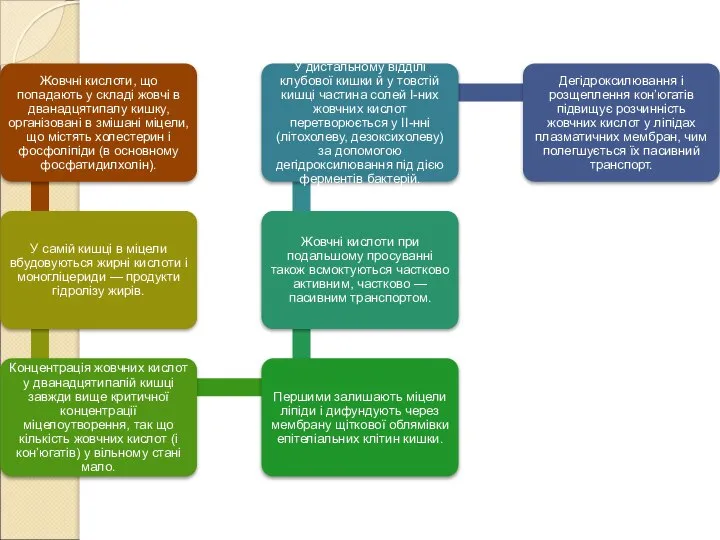
- 206. Білірубін (як і інші жовчні пігменти) реабсорбується в жовчному міхурі і кишечнику в дуже невеликій кількості.
- 207. Рис. Реабсорбція і секреція у жовчному міхурі
- 208. У кишечнику жовч: сприяє розщепленню, омиленню, емульгуванню й усмоктуванню жирів, підсилює перистальтику кишки, що підвищує адсорбцію
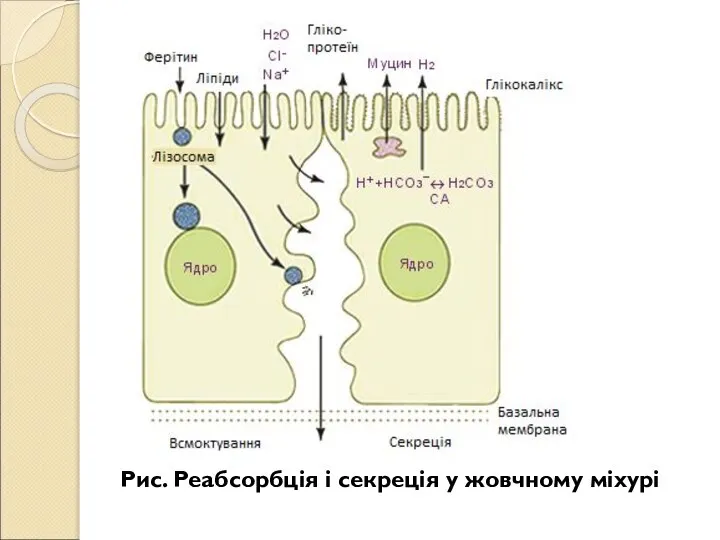
- 209. РЕГУЛЯЦІЯ СЕКРЕЦІЇ ЖОВЧІ Секреція жовчі відбувається постійно, але між прийомами їжі вона накопичується в жовчному міхурі.
- 210. Рис. Механізми наповнення (А) та випорожнення (Б) жовчного міхура
- 211. Ритмічні скорочення (2–6 у хв) спостерігаються також під час травлення і сприяють приливу жовчі в жовчний
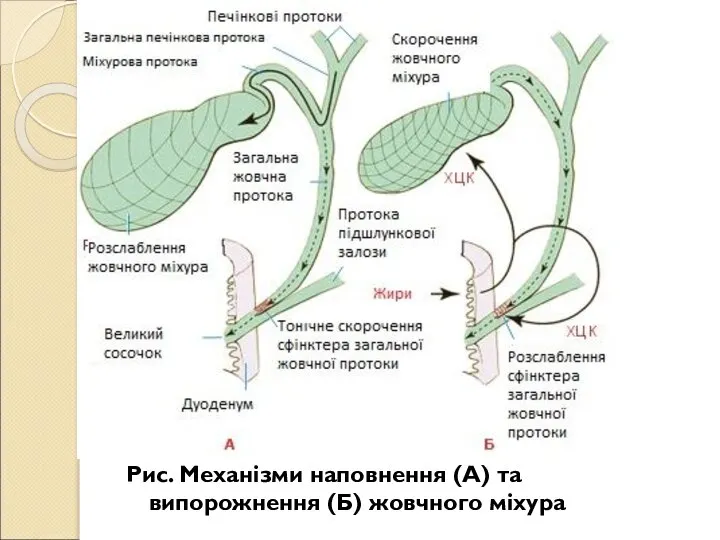
- 212. Рух жовчі до дванадцятипалої кишки відбувається відповідно до законів гідродинаміки вбік найменшого тиску. У процесі травлення
- 213. Регулятори жовчоутворювальної і жовчосекреторної функції печінки: соматостатин, вазопресин, окситоцин, енкефаліни, субстанція П, вазоінтестинальний пептид (ВІП) та
- 214. ВІП є стимулятором секреції води і бікарбонатів холангіоцитами (епітеліальними клітинами жовчних проток) печінки людини і собаки;
- 215. Гормони щитовидної залози також регулюють жовчосекреторну функцію печінки. Наприклад, тироксин і трийодтиронін (Т3 і Т4) не
- 216. Нервові шляхи, по яких до печінки надходять стимулюючі і гальмуючі імпульси, представлені холінергічними волокнами блукаючих і
- 217. Рис. Шляхи перетворення глюкози в печінці. МЕТАБОЛІЧНІ ФУНКЦІЇ ПЕЧІНКИ
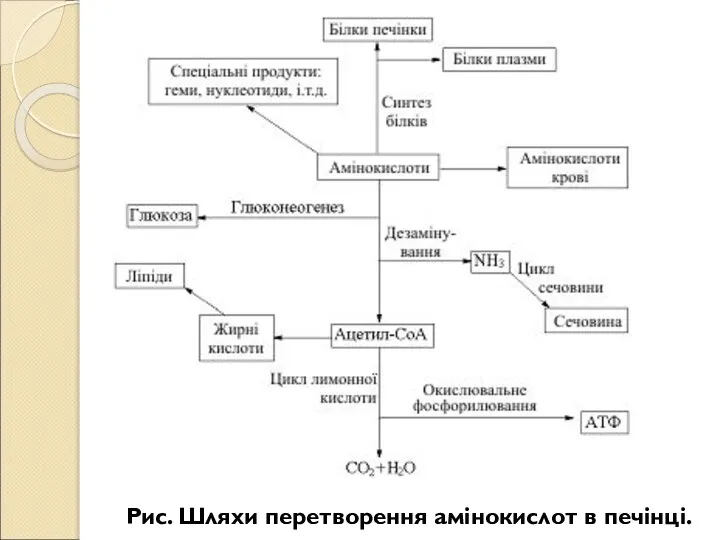
- 218. Рис. Шляхи перетворення амінокислот в печінці.
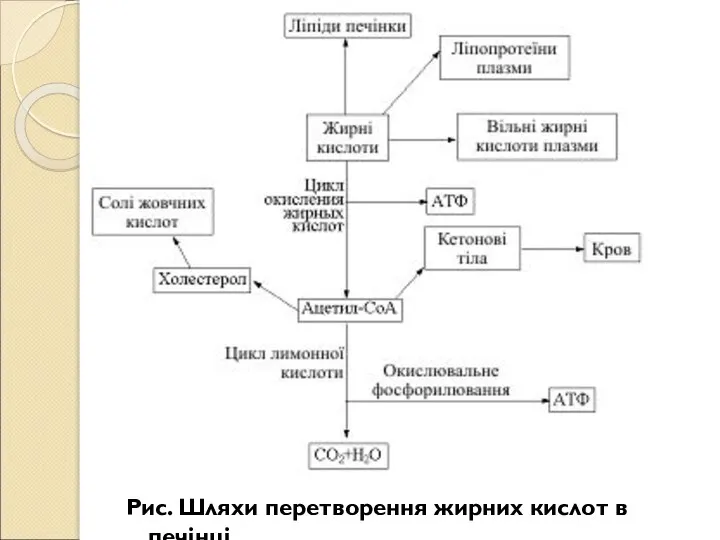
- 219. Рис. Шляхи перетворення жирних кислот в печінці
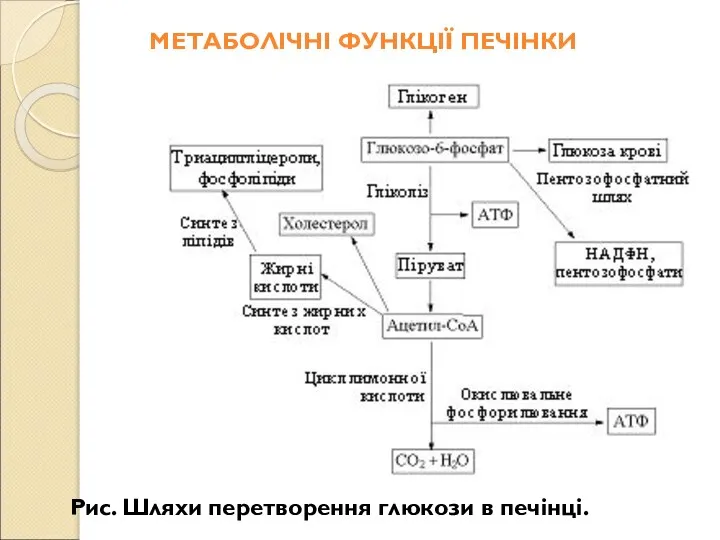
- 220. Крім перетворення і розподілу вуглеводів, жирів і амінокислот, урівноваження коливань в обміні речовин, перетворення надлишку аміногруп
- 221. КИШКОВИЙ СІК: СКЛАД, ВЛАСТИВОСТІ, РЕГУЛЯЦІЯ СОКОВИДІЛЕННЯ Найважливіші кишкові ферменти у великих кількостях зосереджені не в криптах,
- 223. Скачать презентацию
Слайд 2Функції травної системи
секреторна – утворення і виділення залозистими клітинами травних соків (слина,
Функції травної системи
секреторна – утворення і виділення залозистими клітинами травних соків (слина,

Слайд 3захисна (бактерицидна, бактеріостатична, дезінтоксикаційна) – здійснюється завдяки бар’єрним системам шлунково-кишкового тракту і
захисна (бактерицидна, бактеріостатична, дезінтоксикаційна) – здійснюється завдяки бар’єрним системам шлунково-кишкового тракту і

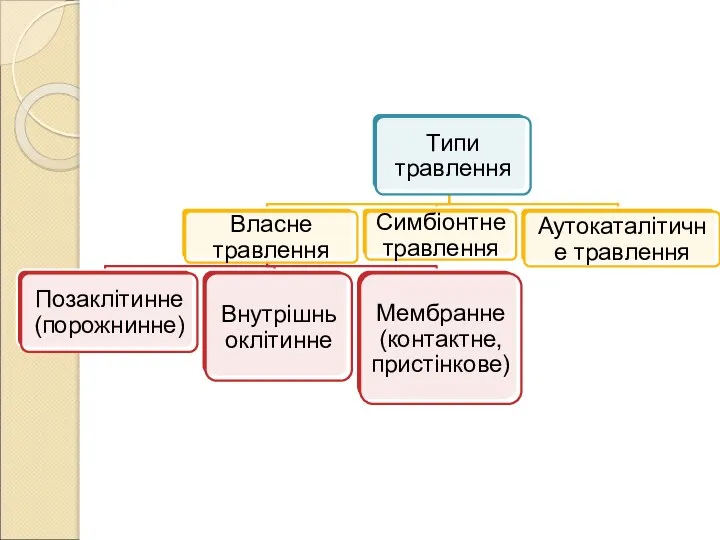
Слайд 5ХАРАКТЕРИСТИКА РІЗНИХ ТИПІВ ТРАВЛЕННЯ
Власний: організм використовує власні ферменти для асиміляції їжі. Ділиться
ХАРАКТЕРИСТИКА РІЗНИХ ТИПІВ ТРАВЛЕННЯ
Власний: організм використовує власні ферменти для асиміляції їжі. Ділиться

Слайд 6Особливості основних типів травлення
Внутрішньоклітинне. Еволюційно та філогенетично найстаріше: лізосомальні ферменти гідролізують харчові
Особливості основних типів травлення
Внутрішньоклітинне. Еволюційно та філогенетично найстаріше: лізосомальні ферменти гідролізують харчові

Слайд 72) Мембранне - здійснюється ферментами, фіксованими на апікальній мембрані ентероцитів , є
2) Мембранне - здійснюється ферментами, фіксованими на апікальній мембрані ентероцитів , є

Слайд 8Рис. Схема позаклітинного (А), внутрішньоклітинного (Б) і мембранного (В) гідролізів харчових речовин.
Рис. Схема позаклітинного (А), внутрішньоклітинного (Б) і мембранного (В) гідролізів харчових речовин.

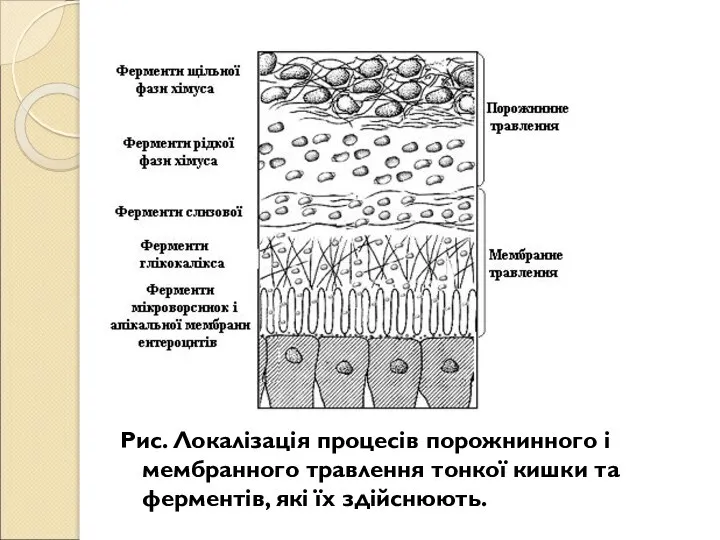
Слайд 9Рис. Локалізація процесів порожнинного і мембранного травлення тонкої кишки та ферментів, які
Рис. Локалізація процесів порожнинного і мембранного травлення тонкої кишки та ферментів, які
Слайд 10Рис. Схема деполімеризації часточок харчових речовин на поверхні мікроворсинок ентероцита, апікальному та
Рис. Схема деполімеризації часточок харчових речовин на поверхні мікроворсинок ентероцита, апікальному та

Слайд 11Травні залози і секреція
Секреція – це складний процес, під час якого секреторна
Травні залози і секреція
Секреція – це складний процес, під час якого секреторна

Слайд 12Секреторний цикл - процеси, які послідовно повторюються в секреторних клітинах. Може бути
Секреторний цикл - процеси, які послідовно повторюються в секреторних клітинах. Може бути

Слайд 131 фаза секреторного циклу– надходження первинних речовин
Основні механізми проникнення речовин через плазматичну
1 фаза секреторного циклу– надходження первинних речовин
Основні механізми проникнення речовин через плазматичну

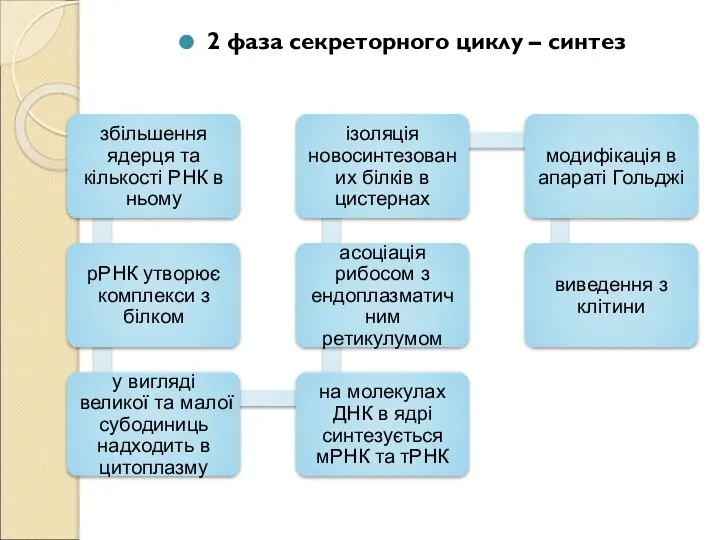
Слайд 142 фаза секреторного циклу – синтез
2 фаза секреторного циклу – синтез
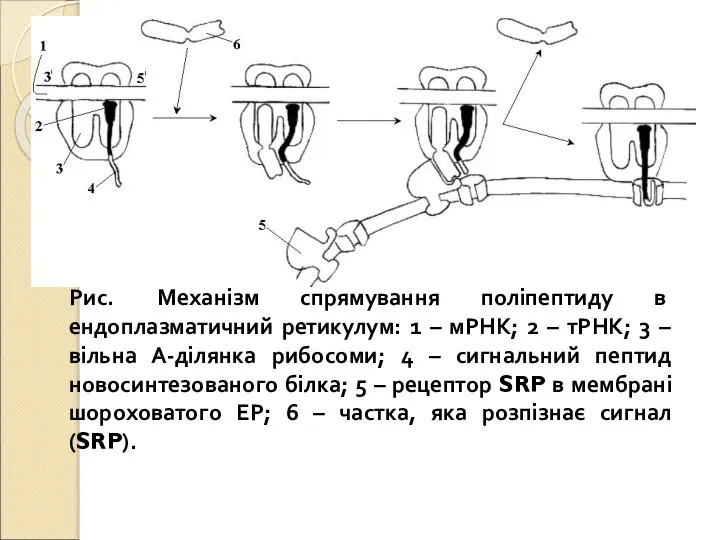
Слайд 15Рис. Механізм спрямування поліпептиду в ендоплазматичний ретикулум: 1 – мРНК; 2 –
Рис. Механізм спрямування поліпептиду в ендоплазматичний ретикулум: 1 – мРНК; 2 –
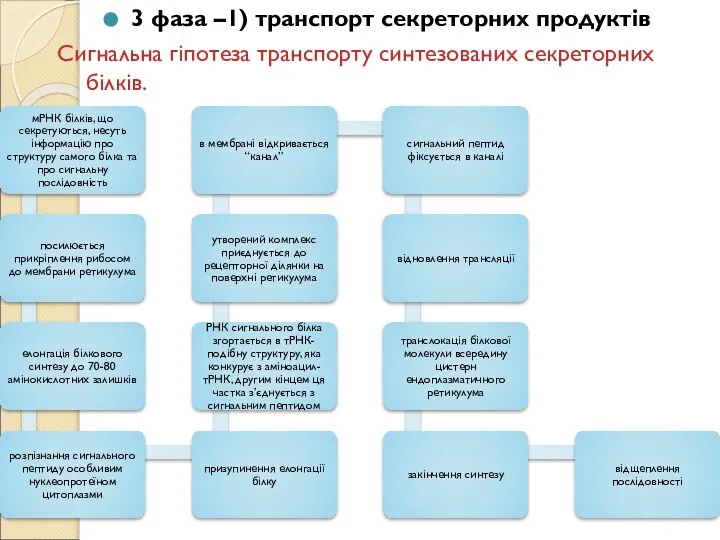
Слайд 163 фаза –1) транспорт секреторних продуктів
Сигнальна гіпотеза транспорту синтезованих секреторних білків.
3 фаза –1) транспорт секреторних продуктів
Сигнальна гіпотеза транспорту синтезованих секреторних білків.
Слайд 173 фаза секреторного циклу – 2) дозрівання секрету
Модифікація поліпептидів секрету в ендоплазматичному
3 фаза секреторного циклу – 2) дозрівання секрету
Модифікація поліпептидів секрету в ендоплазматичному

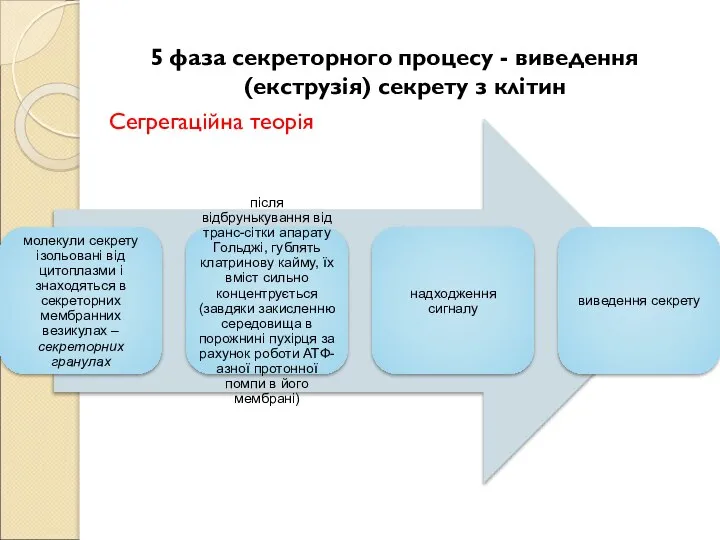
Слайд 185 фаза секреторного процесу - виведення (екструзія) секрету з клітин
Сегрегаційна теорія
5 фаза секреторного процесу - виведення (екструзія) секрету з клітин
Сегрегаційна теорія
Слайд 19Cекреторні клітини за типом виділення секрету:
голокринові (клітини епітелію шлунка), які характеризуються тим,
Cекреторні клітини за типом виділення секрету:
голокринові (клітини епітелію шлунка), які характеризуються тим,

Слайд 20Рис. Типи виділення секрету травними клітинами: 1 – голокриновий; 2 – мікро-

Слайд 21Речовини, які поступають в клітину, використовуються як вихідний матеріал для секреторного продукту
Речовини, які поступають в клітину, використовуються як вихідний матеріал для секреторного продукту

Слайд 22Секретовані речовини виводяться з клітини за конститутивним та регульованим механізмами (молекули зберігаються
Секретовані речовини виводяться з клітини за конститутивним та регульованим механізмами (молекули зберігаються

Слайд 235 етапів регульованого екзоцитозу:
1) спряження стимул-секреція – це послідовність процесів,
5 етапів регульованого екзоцитозу:
1) спряження стимул-секреція – це послідовність процесів,

Слайд 24Захисні механізми травного тракту
Органи травного тракту виконують захисну функцію завдяки центральним рефлекторним
Захисні механізми травного тракту
Органи травного тракту виконують захисну функцію завдяки центральним рефлекторним

Слайд 25Нудота – неприємне, суб’єктивне відчуття, формується в ЦНС при значному підвищенні збудження
Нудота – неприємне, суб’єктивне відчуття, формується в ЦНС при значному підвищенні збудження

Слайд 26Форми нудоти:
1) мозкова;
2) рефлекторна виникає під час механічного або токсичного подразнення кореня
Форми нудоти:
1) мозкова;
2) рефлекторна виникає під час механічного або токсичного подразнення кореня

Слайд 27Блювота – це захисна складнокоординована системна рефлекторна реакція. Вона виникає при збудженні
Блювота – це захисна складнокоординована системна рефлекторна реакція. Вона виникає при збудженні

Слайд 28Аферентні впливи. На блювотний рефлекс впливають вищі відділи стовбуру мозку і кора.
Аферентні впливи. На блювотний рефлекс впливають вищі відділи стовбуру мозку і кора.

Слайд 29Ефекторні впливи по доцентровим волокнам в складі блукаючого і черевного нервів передаються
Ефекторні впливи по доцентровим волокнам в складі блукаючого і черевного нервів передаються

Слайд 30Акту блювоти передує виникнення нудоти і антиперистальтики. Антиперистальтика в кишечнику фізіологічно обумовлена
Акту блювоти передує виникнення нудоти і антиперистальтики. Антиперистальтика в кишечнику фізіологічно обумовлена

Слайд 31НЕСПЕЦИФІЧНІ МЕХАНІЗМИ ЗАХИСТУ
З харчовими речовинами в травну систему попадають різноманітні шкідливі речовини
НЕСПЕЦИФІЧНІ МЕХАНІЗМИ ЗАХИСТУ
З харчовими речовинами в травну систему попадають різноманітні шкідливі речовини

Слайд 32нормальна рухова і секреторна активність тонкого кишечника. Постійна секреція рідини в криптах
нормальна рухова і секреторна активність тонкого кишечника. Постійна секреція рідини в криптах

Слайд 33Захисні механізми ентероцита: ендоплазматична сітка, комплекс Гольджі, в яких частина харчових речовин
Захисні механізми ентероцита: ендоплазматична сітка, комплекс Гольджі, в яких частина харчових речовин

Слайд 34Механічна або пасивна система захисної системи ШКТ:
обмеження проникності слизової оболонки для водорозчинних
Механічна або пасивна система захисної системи ШКТ:
обмеження проникності слизової оболонки для водорозчинних

Слайд 35СПЕЦИФІЧНІ ІМУННІ МЕХАНІЗМИ ЗАХИСТУ
Специфічні імунні механізми забезпечуються імунокомпетентною “лімфоїдною тканиною”:
інтраепітеліальні Т і
СПЕЦИФІЧНІ ІМУННІ МЕХАНІЗМИ ЗАХИСТУ
Специфічні імунні механізми забезпечуються імунокомпетентною “лімфоїдною тканиною”:
інтраепітеліальні Т і

Слайд 36Вплив імуноглобулінів:
антивірусний;
антитоксичний;
протимікробний;
місцевий імунітет;
антипаразитарний.
Первинна імунна нейтралізація антигенів відбувається в пристінковому шарі в процесі
Вплив імуноглобулінів:
антивірусний;
антитоксичний;
протимікробний;
місцевий імунітет;
антипаразитарний.
Первинна імунна нейтралізація антигенів відбувається в пристінковому шарі в процесі

Слайд 37SIgA складається з підкласів SIgA1 (25-30%), SIgA2 (60-70%), SIgA – мономер (5%).
SIgA складається з підкласів SIgA1 (25-30%), SIgA2 (60-70%), SIgA – мономер (5%).

Слайд 38Недиференційовані лімфоцити, розташовані у власному шарі або пейєрових бляшках продукують секреторні антитіла
Недиференційовані лімфоцити, розташовані у власному шарі або пейєрових бляшках продукують секреторні антитіла

Слайд 39Імунна система організму, кишечні гідролітичні ферменти і система тканинної детоксикації (апарат окислювального
Імунна система організму, кишечні гідролітичні ферменти і система тканинної детоксикації (апарат окислювального

Слайд 40ТРАВЛЕННЯ В РОТОВІЙ ПОРОЖНИНІ
ТРАВЛЕННЯ В РОТОВІЙ ПОРОЖНИНІ
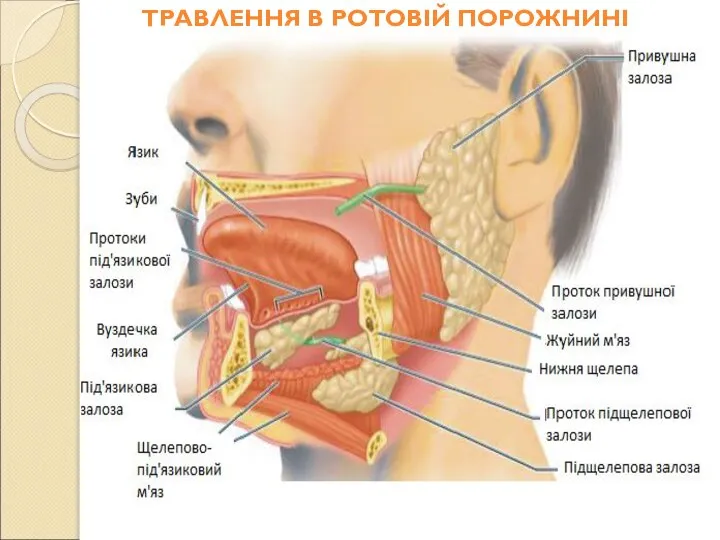
Слайд 41Ротова порожнина, глотка і стравохід - функціональний комплекс, який забезпечує первинну обробку
Ротова порожнина, глотка і стравохід - функціональний комплекс, який забезпечує первинну обробку

Слайд 42СКЛАД І ВЛАСТИВОСТІ СЛИНИ
Слина – сукупний зовнішньо-секреторний продукт привушних, підщелепних, під’язикових великих
СКЛАД І ВЛАСТИВОСТІ СЛИНИ
Слина – сукупний зовнішньо-секреторний продукт привушних, підщелепних, під’язикових великих

Слайд 43Сухий залишок містить: неорганічні та органічні речовини (›50%).Неорганічні компоненти: Na+, K+, Ca2+,
Сухий залишок містить: неорганічні та органічні речовини (›50%).Неорганічні компоненти: Na+, K+, Ca2+,

Слайд 44РЕГУЛЯЦІЯ СЛИНОВИДІЛЕННЯ
Секреторна функція слинних залоз і слиновиділення регулюються рефлекторно. Розрізняють безумовно- і
РЕГУЛЯЦІЯ СЛИНОВИДІЛЕННЯ
Секреторна функція слинних залоз і слиновиділення регулюються рефлекторно. Розрізняють безумовно- і

Слайд 45До слинних залоз імпульси передаються парасимпатичними і симпатичними нервовими волокнами.
Парасимпатична іннервація привушних
До слинних залоз імпульси передаються парасимпатичними і симпатичними нервовими волокнами.
Парасимпатична іннервація привушних

Слайд 46Симпатична іннервація слинних залоз здійснюється волокнами, що відходять від верхніх шийних симпатичних
Симпатична іннервація слинних залоз здійснюється волокнами, що відходять від верхніх шийних симпатичних

Слайд 47Умовно-рефлекторне слиновиділення викликають вид і запах їжі, а також звуки, пов'язані з
Умовно-рефлекторне слиновиділення викликають вид і запах їжі, а також звуки, пов'язані з

Слайд 48МЕХАНІЧНІ ПРОЦЕСИ: ЖУВАННЯ І КОВТАННЯ
Жування — це процес механічної обробки їжі, що
МЕХАНІЧНІ ПРОЦЕСИ: ЖУВАННЯ І КОВТАННЯ
Жування — це процес механічної обробки їжі, що

Слайд 49У людини і більшості тварин верхня щелепа нерухома, тому жування здійснюється рухами
У людини і більшості тварин верхня щелепа нерухома, тому жування здійснюється рухами

Слайд 50Жування відбувається мимоволі, однак включає і довільні компоненти. Регуляція здійснюється на різних
Жування відбувається мимоволі, однак включає і довільні компоненти. Регуляція здійснюється на різних

Слайд 51Ковтання — складний рефлекторний акт, у процесі якого їжа переходить з ротової
Ковтання — складний рефлекторний акт, у процесі якого їжа переходить з ротової
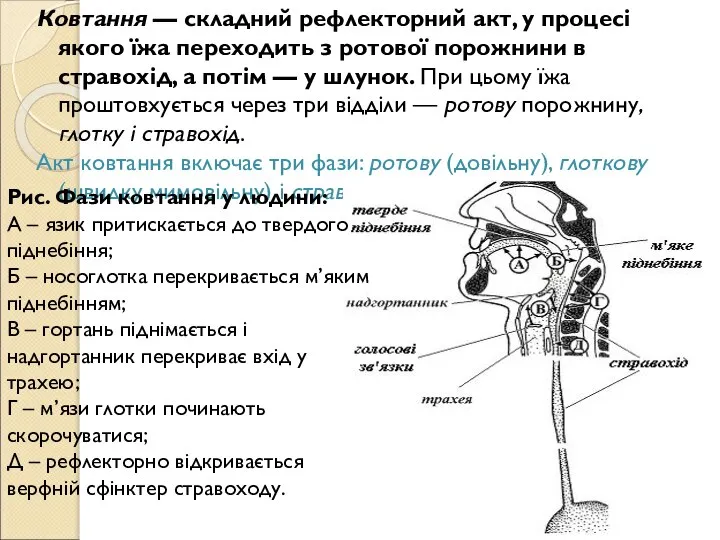
Слайд 52У глоткову фазу: харчова грудка на корені язика подразнює чутливі нервові закінчення
У глоткову фазу: харчова грудка на корені язика подразнює чутливі нервові закінчення

Слайд 53Стравохідна фаза: потрапивши в стравохід, харчову грудку проходить в шлунок, що складає
Стравохідна фаза: потрапивши в стравохід, харчову грудку проходить в шлунок, що складає

Слайд 54Перистальтика стравоходу
Рухи стравоходу взаємопов’язані з рухами ковтального апарату і викликаються рефлекторно при
Перистальтика стравоходу
Рухи стравоходу взаємопов’язані з рухами ковтального апарату і викликаються рефлекторно при

Слайд 55Швидкість поширення перистальтичної хвилі залежить від консистенції харчової грудки і положення тіла.
Швидкість поширення перистальтичної хвилі залежить від консистенції харчової грудки і положення тіла.

Слайд 56ПОРУШЕННЯ НОРМАЛЬНОЇ ФУНКЦІЇ СТРАВОХОДУ
При ахалазії (спазм кардіального відділу стравоходу і розширення його
ПОРУШЕННЯ НОРМАЛЬНОЇ ФУНКЦІЇ СТРАВОХОДУ
При ахалазії (спазм кардіального відділу стравоходу і розширення його

Слайд 57Скорочення м'язів стравоходу пов'язані зі збудженням холінергічних нейронів. Їх розслаблення не обумовлене
Скорочення м'язів стравоходу пов'язані зі збудженням холінергічних нейронів. Їх розслаблення не обумовлене

Слайд 58 ТРАВЛЕННЯ В ШЛУНКУ
ТРАВЛЕННЯ В ШЛУНКУ
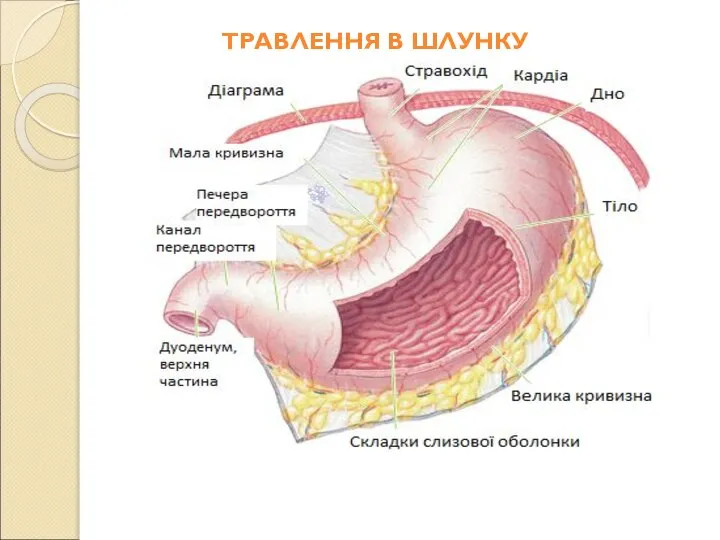
Слайд 59Шлунок – це відділ травної системи, в якому накопичується проковтнута їжа. Вона

Слайд 60ФУНКЦІЇ ШЛУНКУ:
Депонування їжі: в проксимальному відділі шлунка відсутній ритм збудження і перистальтики.
ФУНКЦІЇ ШЛУНКУ:
Депонування їжі: в проксимальному відділі шлунка відсутній ритм збудження і перистальтики.

Слайд 61Секреторна функція: виділення соку шлунковими залозами. Сік містить іони і макромолекули, забезпечує
Секреторна функція: виділення соку шлунковими залозами. Сік містить іони і макромолекули, забезпечує

Слайд 62Рухова функція: перемішування їжі з шлунковим соком і її просування в дванадцятипалу
Рухова функція: перемішування їжі з шлунковим соком і її просування в дванадцятипалу

Слайд 63Інкреторна: утворення біологічно активних речовин (гормонів): гастрину, гістаміну, соматостатину, мотиліну й інших
Інкреторна: утворення біологічно активних речовин (гормонів): гастрину, гістаміну, соматостатину, мотиліну й інших

Слайд 64СКЛАД І ВЛАСТИВОСТІ ШЛУНКОВОГО СОКУ
Шлунковий сік — продукт зовнішньо секреторної і екскреторної
СКЛАД І ВЛАСТИВОСТІ ШЛУНКОВОГО СОКУ
Шлунковий сік — продукт зовнішньо секреторної і екскреторної

Слайд 65Неорганічні речовини: хлориди, сульфати, фосфати, бікарбонати натрію, калію, кальцію, магнію. Соляна кислота
Неорганічні речовини: хлориди, сульфати, фосфати, бікарбонати натрію, калію, кальцію, магнію. Соляна кислота

Слайд 66Табл. Амінокислотний склад (в мг на 1 г азоту) шлункового соку здорової
Табл. Амінокислотний склад (в мг на 1 г азоту) шлункового соку здорової
Слайд 67Ферменти шлункового соку. Виявлено щонайменше 8 протеолітичних ферментів. 5 швидко мігруючих пепсиногенів
Ферменти шлункового соку. Виявлено щонайменше 8 протеолітичних ферментів. 5 швидко мігруючих пепсиногенів

Слайд 68Важливий компонент шлункового соку — слиз, що захищає слизову від механічного і
Важливий компонент шлункового соку — слиз, що захищає слизову від механічного і

Слайд 69Рис. Схематичне зображення шлункових залоз
Рис. Схематичне зображення шлункових залоз
Слайд 70РЕГУЛЯЦІЯ ШЛУНКОВОЇ СЕКРЕЦІЇ
Переривчастість або безперервність шлункової секреції залежить від умов харчування, що
РЕГУЛЯЦІЯ ШЛУНКОВОЇ СЕКРЕЦІЇ
Переривчастість або безперервність шлункової секреції залежить від умов харчування, що

Слайд 71Шлункова секреція регулюється комплексом ендокринних, паракринних, нейрональних і місцевих механізмів (парасимпатических, симпатичних,
Шлункова секреція регулюється комплексом ендокринних, паракринних, нейрональних і місцевих механізмів (парасимпатических, симпатичних,

Слайд 72ПЕРША ЦЕФАЛІЧНА (СКЛАДНОРЕФЛЕКТОРНА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ
ПЕРША ЦЕФАЛІЧНА (СКЛАДНОРЕФЛЕКТОРНА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ

Слайд 73Важливу роль у нейрогенній стимуляції секреторних процесів відіграє гастрин. Видалення пілоричного відділу
Важливу роль у нейрогенній стимуляції секреторних процесів відіграє гастрин. Видалення пілоричного відділу

Слайд 74Швидкість секреції в цефалічну фазу складає 40-50% від максимальної. Це було докладно
Швидкість секреції в цефалічну фазу складає 40-50% від максимальної. Це було докладно
Слайд 75Удаване годування, що викликає збудження блукаючих нервів, здійснює як гальмівний, так і
Удаване годування, що викликає збудження блукаючих нервів, здійснює як гальмівний, так і

Слайд 76 Усі гальмівні нейропептиди, окрім бомбезина, справляють свій вплив на шлункову секрецію
Усі гальмівні нейропептиди, окрім бомбезина, справляють свій вплив на шлункову секрецію

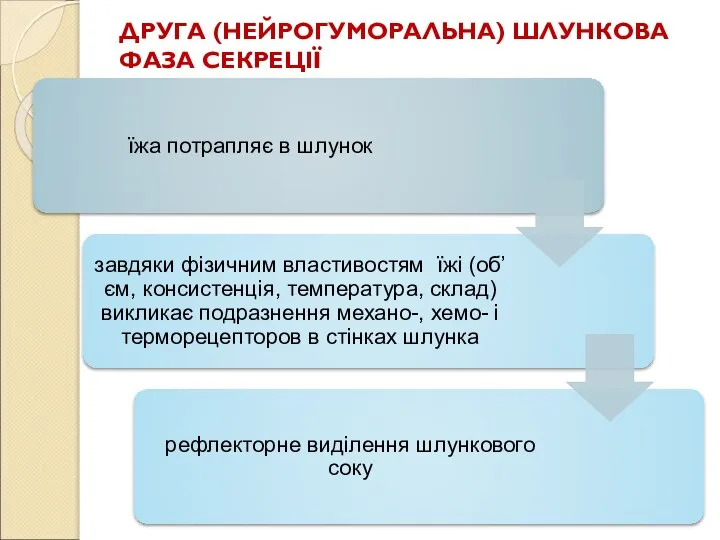
Слайд 77ДРУГА (НЕЙРОГУМОРАЛЬНА) ШЛУНКОВА ФАЗА СЕКРЕЦІЇ
ДРУГА (НЕЙРОГУМОРАЛЬНА) ШЛУНКОВА ФАЗА СЕКРЕЦІЇ
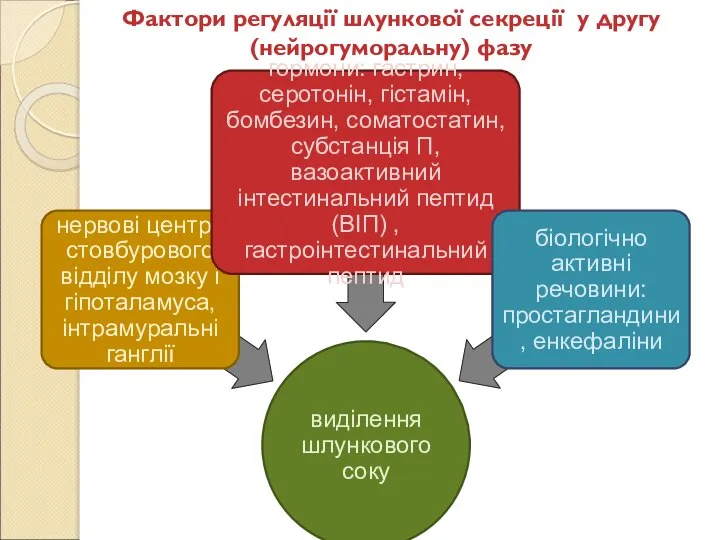
Слайд 78Фактори регуляції шлункової секреції у другу (нейрогуморальну) фазу
Фактори регуляції шлункової секреції у другу (нейрогуморальну) фазу
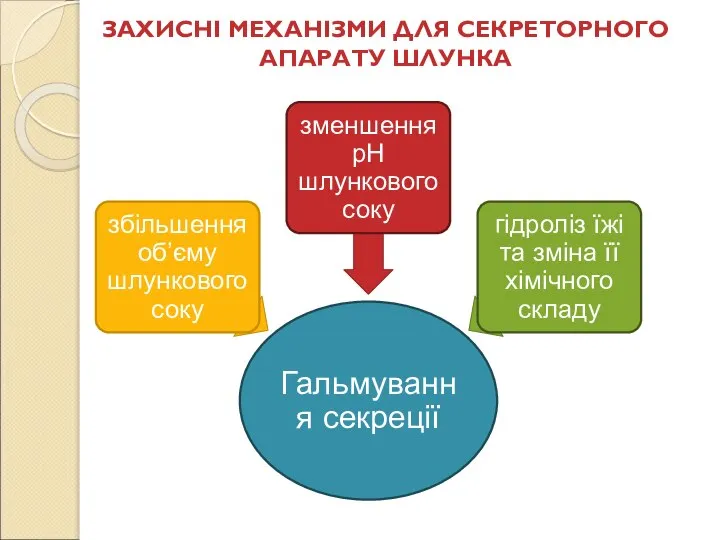
Слайд 79ЗАХИСНІ МЕХАНІЗМИ ДЛЯ СЕКРЕТОРНОГО АПАРАТУ ШЛУНКА
ЗАХИСНІ МЕХАНІЗМИ ДЛЯ СЕКРЕТОРНОГО АПАРАТУ ШЛУНКА
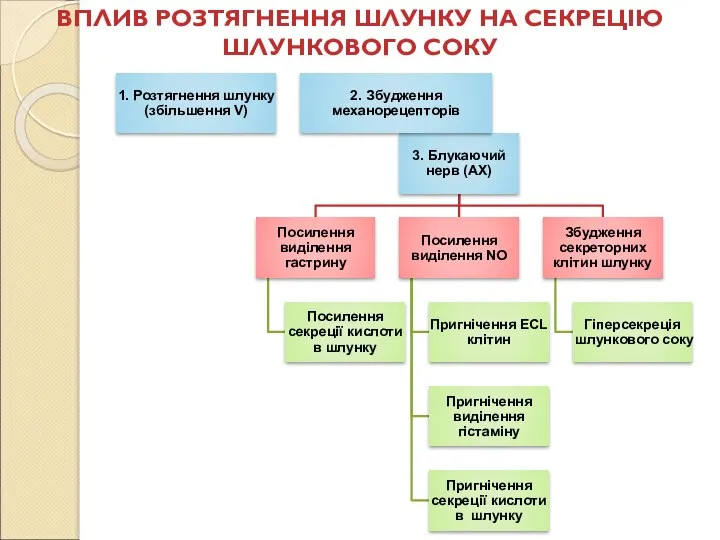
Слайд 80ВПЛИВ РОЗТЯГНЕННЯ ШЛУНКУ НА СЕКРЕЦІЮ ШЛУНКОВОГО СОКУ
ВПЛИВ РОЗТЯГНЕННЯ ШЛУНКУ НА СЕКРЕЦІЮ ШЛУНКОВОГО СОКУ
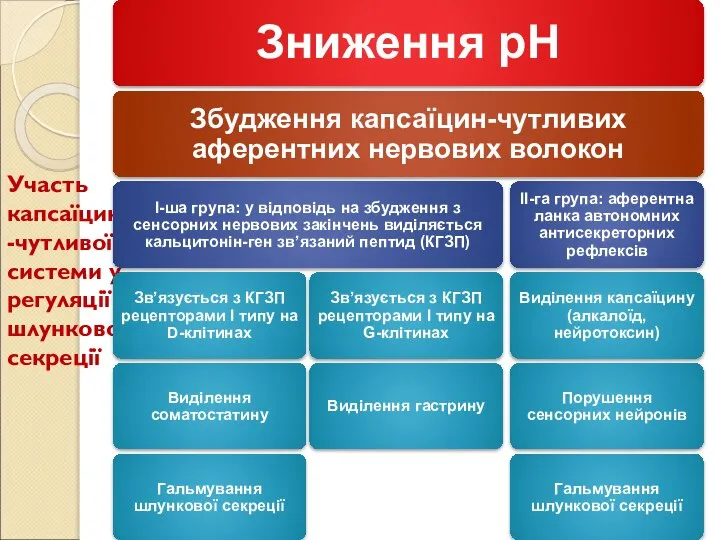
Слайд 81Участь капсаїцин-чутливої системи у регуляції шлункової секреції
Участь капсаїцин-чутливої системи у регуляції шлункової секреції
Слайд 82ШЛУНКОВА ФАЗА ГАЛЬМУВАННЯ
Це гальмування звільнення гастрина при омиванні антральної слизової кислим розчином.
ШЛУНКОВА ФАЗА ГАЛЬМУВАННЯ
Це гальмування звільнення гастрина при омиванні антральної слизової кислим розчином.

Слайд 83Дія соматостатину:
гальмує виділення гормонів гіпофіза;
гальмує виділення інсуліну і глюкагона, стимульоване
Дія соматостатину:
гальмує виділення гормонів гіпофіза;
гальмує виділення інсуліну і глюкагона, стимульоване

Слайд 84Шлунковий соматостатин – паракринний гальмівний регулятор секреції кислоти в шлунку. Виділяється в
Шлунковий соматостатин – паракринний гальмівний регулятор секреції кислоти в шлунку. Виділяється в

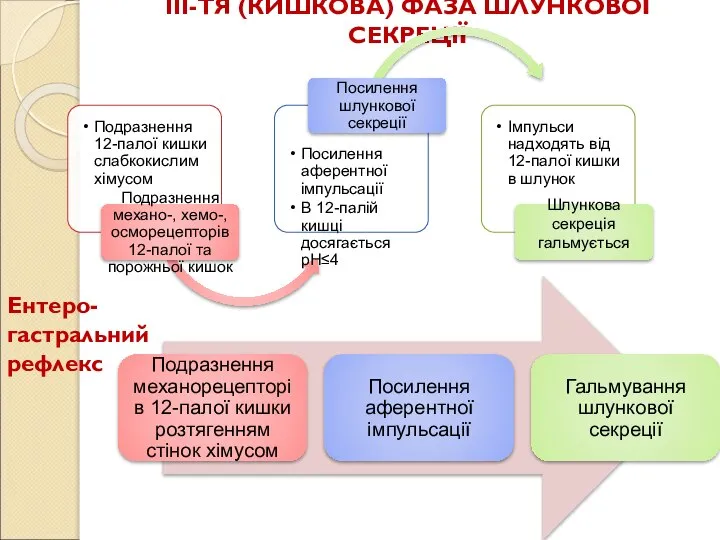
Слайд 85ІІІ-ТЯ (КИШКОВА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ
Ентеро-гастральний рефлекс
ІІІ-ТЯ (КИШКОВА) ФАЗА ШЛУНКОВОЇ СЕКРЕЦІЇ
Ентеро-гастральний рефлекс
Слайд 86Гальмування шлункової секреції продовжується під дією секретину, що звільняється S-клітинами слизової оболонки
Гальмування шлункової секреції продовжується під дією секретину, що звільняється S-клітинами слизової оболонки

Слайд 87У собак видалення проксимального відділу тонкої кишки, зменшує рівень секретину в плазмі
У собак видалення проксимального відділу тонкої кишки, зменшує рівень секретину в плазмі
Слайд 88Гальмівний вплив секретину не залежить від вагальної іннервації фундального відділу шлунка. Однак,
Гальмівний вплив секретину не залежить від вагальної іннервації фундального відділу шлунка. Однак,

Слайд 89Жир, введений у тонкий кишечник, гальмує секрецію кислоти в шлунку. Механізм гальмування
Жир, введений у тонкий кишечник, гальмує секрецію кислоти в шлунку. Механізм гальмування

Слайд 90До ентерогастронів відносять:
гастроінгібуючий поліпептид,
секретин,
глюкагоно-подібні пептиди 1 і 2,
пептид YY,
До ентерогастронів відносять:
гастроінгібуючий поліпептид,
секретин,
глюкагоно-подібні пептиди 1 і 2,
пептид YY,

Слайд 91ШЛУНКОВИЙ СЛИЗ: УТВОРЕННЯ І ВИДІЛЕННЯ
Слиз – складна динамічна система колоїдних розчинів високомолекулярних
ШЛУНКОВИЙ СЛИЗ: УТВОРЕННЯ І ВИДІЛЕННЯ
Слиз – складна динамічна система колоїдних розчинів високомолекулярних

Слайд 92До складу слизу входять нейтральні мукополісахариди, сіаломуцини, глікопротеїди і протеоглікани (кислі мукополісахариди).
Слиз
До складу слизу входять нейтральні мукополісахариди, сіаломуцини, глікопротеїди і протеоглікани (кислі мукополісахариди).
Слиз

Слайд 93При значеннях рН між 7,0 і 7,9 шлунковий слиз має мінімальну в'язкість;
При значеннях рН між 7,0 і 7,9 шлунковий слиз має мінімальну в'язкість;

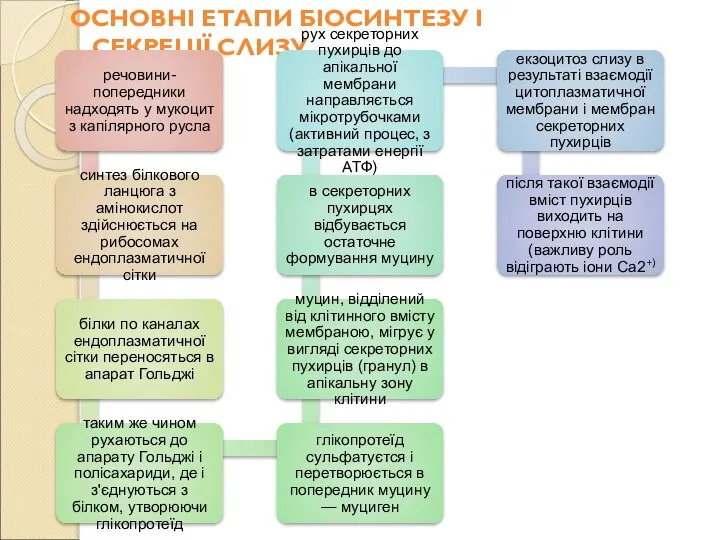
Слайд 94ОСНОВНІ ЕТАПИ БІОСИНТЕЗУ І СЕКРЕЦІЇ СЛИЗУ
ОСНОВНІ ЕТАПИ БІОСИНТЕЗУ І СЕКРЕЦІЇ СЛИЗУ
Слайд 95Шлунковий слиз утворюється в результаті взаємодії мукоїдних речовин, що надходять у просвіт
Шлунковий слиз утворюється в результаті взаємодії мукоїдних речовин, що надходять у просвіт

Слайд 96Мукоцити одночасно з муцином продукують бікарбонат. Муцин взаємодіє з бікарбонатом, в результаті
Мукоцити одночасно з муцином продукують бікарбонат. Муцин взаємодіє з бікарбонатом, в результаті

Слайд 97На підставі гістохімічного аналізу виділено чотири основних класи біополімерів, що утворюють нерозчинний
На підставі гістохімічного аналізу виділено чотири основних класи біополімерів, що утворюють нерозчинний

Слайд 983. Сіаломуцини беруть участь у синтезі соляної кислоти. Сиалові сульфоглікопротеїни синтезуються поверхневим
3. Сіаломуцини беруть участь у синтезі соляної кислоти. Сиалові сульфоглікопротеїни синтезуються поверхневим

Слайд 99Шлунковий слиз, бікарбонати, фосфоліпіди і вода утворюють гель, що створює фізико-хімічний бар'єр
Шлунковий слиз, бікарбонати, фосфоліпіди і вода утворюють гель, що створює фізико-хімічний бар'єр

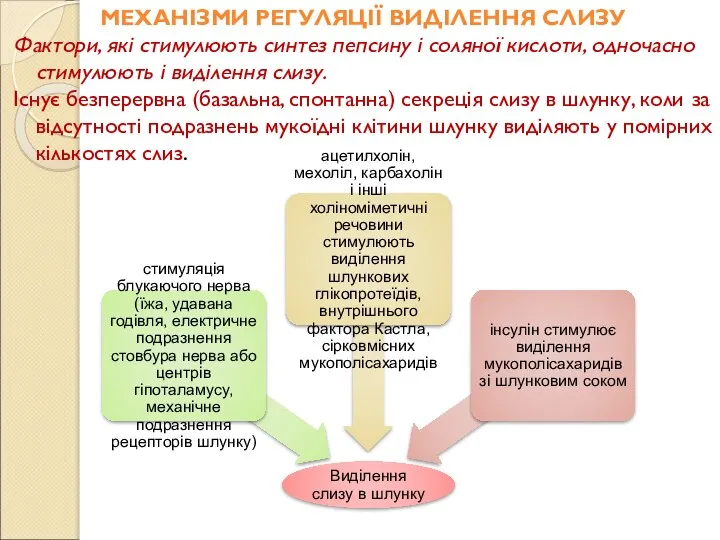
Слайд 100МЕХАНІЗМИ РЕГУЛЯЦІЇ ВИДІЛЕННЯ СЛИЗУ
Фактори, які стимулюють синтез пепсину і соляної кислоти, одночасно
МЕХАНІЗМИ РЕГУЛЯЦІЇ ВИДІЛЕННЯ СЛИЗУ
Фактори, які стимулюють синтез пепсину і соляної кислоти, одночасно
Слайд 101МЕХАНІЗМ ВПЛИВУ ІНСУЛІНУ НА СЕКРЕЦІЮ ТА ВИДІЛЕННЯ ШЛУНКОВОГО СЛИЗУ
МЕХАНІЗМ ВПЛИВУ ІНСУЛІНУ НА СЕКРЕЦІЮ ТА ВИДІЛЕННЯ ШЛУНКОВОГО СЛИЗУ
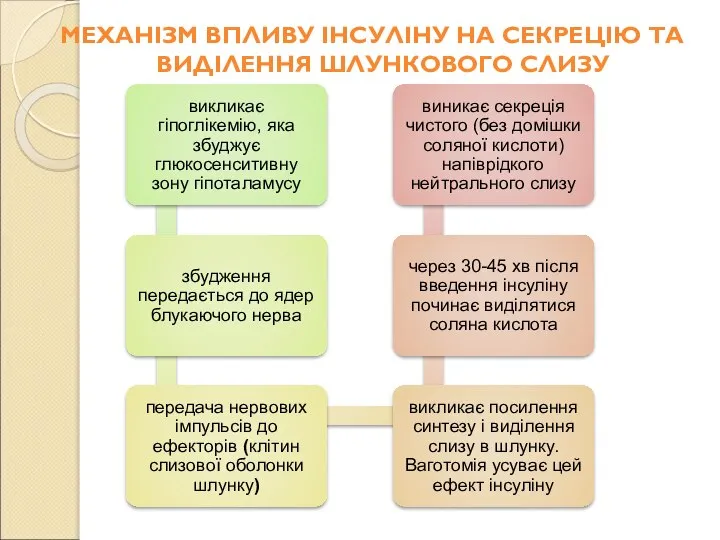
Слайд 102Інсулін стимулює також секрецію внутрішнього фактора Кастла за механізмом, аналогічним стимуляції секреції
Інсулін стимулює також секрецію внутрішнього фактора Кастла за механізмом, аналогічним стимуляції секреції

Слайд 103Місцеве механічне подразнення значно стимулює секрецію слизу мукоїдними клітинами антрального і фундального
Місцеве механічне подразнення значно стимулює секрецію слизу мукоїдними клітинами антрального і фундального

Слайд 104ГОРМОНАЛЬНІ ФАКТОРИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СЛИЗУ
Істотну роль відіграють гормони гіпофізарно-адреналової системи. АКТГ стимулює
ГОРМОНАЛЬНІ ФАКТОРИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СЛИЗУ
Істотну роль відіграють гормони гіпофізарно-адреналової системи. АКТГ стимулює

Слайд 105Серотонін (5-гідрокситриптамін) є однією з ланок гормональної регуляції секреції мукоїдних речовин. Він
Серотонін (5-гідрокситриптамін) є однією з ланок гормональної регуляції секреції мукоїдних речовин. Він

Слайд 106Участь місцевого подразнення слизової оболонки шлунку
Механічне і хімічне (гормони, нейротропні речовини,
Участь місцевого подразнення слизової оболонки шлунку
Механічне і хімічне (гормони, нейротропні речовини,

Слайд 107МЕХАНІЗМИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СОЛЯНОЇ КИСЛОТИ І ЇЇ РОЛЬ В ТРАВЛЕННІ
Обкладові клітини мають
МЕХАНІЗМИ РЕГУЛЯЦІЇ СЕКРЕЦІЇ СОЛЯНОЇ КИСЛОТИ І ЇЇ РОЛЬ В ТРАВЛЕННІ
Обкладові клітини мають

Слайд 108БУДОВА ОБКЛАДОВИХ КЛІТИН
Обкладові клітини містять численні великі мітохондрії, характерні внутрішньоклітинні канальці, вистелені
БУДОВА ОБКЛАДОВИХ КЛІТИН
Обкладові клітини містять численні великі мітохондрії, характерні внутрішньоклітинні канальці, вистелені

Слайд 109Рис. Обкладова клітина в спокої (А) і при стимуляції (Б): Я –
Рис. Обкладова клітина в спокої (А) і при стимуляції (Б): Я –

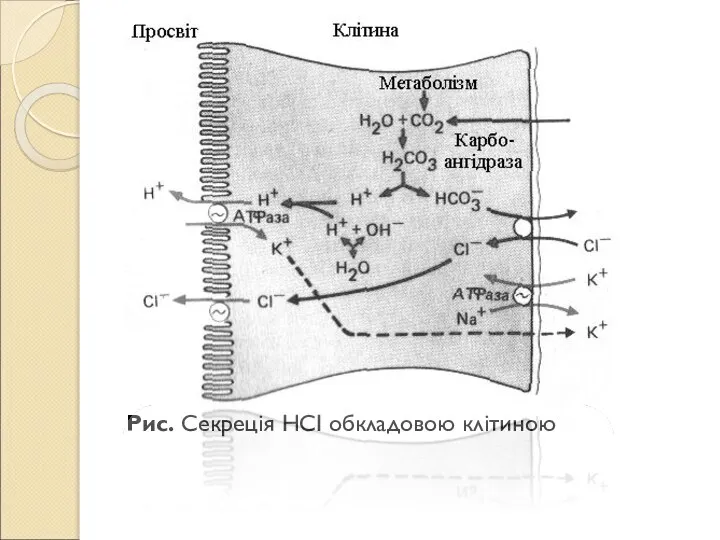
Слайд 110В мікроворсинках канальців локалізована H+,K+-АТФаза, що транспортує іони Н+. Обкладові клітини утворюють
В мікроворсинках канальців локалізована H+,K+-АТФаза, що транспортує іони Н+. Обкладові клітини утворюють

Слайд 111Рис. Секреція НСІ обкладовою клітиною
Рис. Секреція НСІ обкладовою клітиною
Слайд 112В секреторній фазі секретуєтся більше іонів Cl-, ніж іонів Н+.
У стані
В секреторній фазі секретуєтся більше іонів Cl-, ніж іонів Н+.
У стані

Слайд 113Активація секреторного процесу в обкладових клітинах є результатом інтеграції нейрогуморальних механізмів.
Медіатори-первинні посередники
Активація секреторного процесу в обкладових клітинах є результатом інтеграції нейрогуморальних механізмів.
Медіатори-первинні посередники

Слайд 114Гуморальні чинники регуляції шлункової секреції
На обкладовій клітині знаходиться три види рецепторів, збудження
Гуморальні чинники регуляції шлункової секреції
На обкладовій клітині знаходиться три види рецепторів, збудження

Слайд 115Слизова оболонка містить піридоксальзалежний фермент гістидиндекарбоксилазу, необхідний для синтезу гістамину.
Гістидиндекарбоксилаза активується
Слизова оболонка містить піридоксальзалежний фермент гістидиндекарбоксилазу, необхідний для синтезу гістамину.
Гістидиндекарбоксилаза активується

Слайд 116У шлунку ссавців виділяють два депо гістаміну: гладкі і ентерохромафіно-подібні (ECL) клітини.
У шлунку ссавців виділяють два депо гістаміну: гладкі і ентерохромафіно-подібні (ECL) клітини.

Слайд 117В нормальних регуляторних процесах бере участь гістамін ЕСL-клітин.
ECL-клітини — це маленькі
В нормальних регуляторних процесах бере участь гістамін ЕСL-клітин.
ECL-клітини — це маленькі

Слайд 118Рецептори, що гальмують активність на ECL-клітинах:
SST2 чутливі до соматостатину
CGRP, чутливі до кальцитонін
Рецептори, що гальмують активність на ECL-клітинах:
SST2 чутливі до соматостатину
CGRP, чутливі до кальцитонін

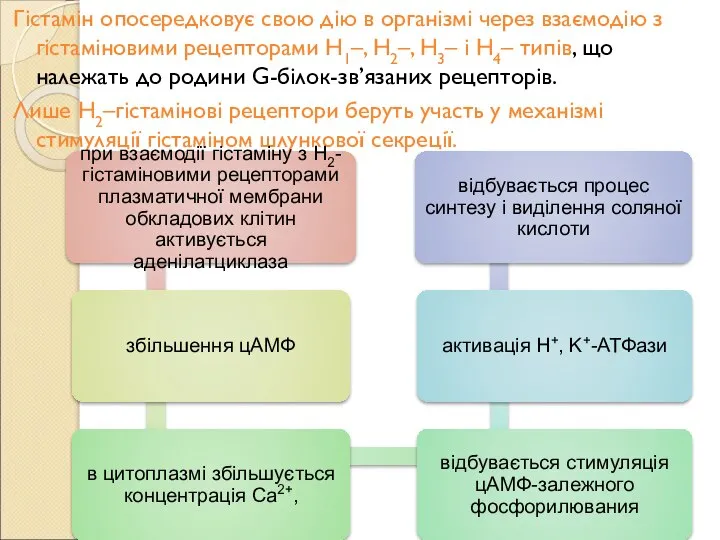
Слайд 119Гістамін опосередковує свою дію в організмі через взаємодію з гістаміновими рецепторами Н1–,
Гістамін опосередковує свою дію в організмі через взаємодію з гістаміновими рецепторами Н1–,
Слайд 120Н2-блокатори гальмують стимульовану гістаміном секрецію кислоти в шлунку людини, собак, кішок, щурів,
Н2-блокатори гальмують стимульовану гістаміном секрецію кислоти в шлунку людини, собак, кішок, щурів,

Слайд 121В міжхарчову фазу секреторного процесу гістамін зберігається, головним чином, в секреторних гранулах
В міжхарчову фазу секреторного процесу гістамін зберігається, головним чином, в секреторних гранулах

Слайд 122Ацетилхолін виділяється з терміналей постгангліонарних холінергічних нейронів і впливає на обкладові клітини
Ацетилхолін виділяється з терміналей постгангліонарних холінергічних нейронів і впливає на обкладові клітини
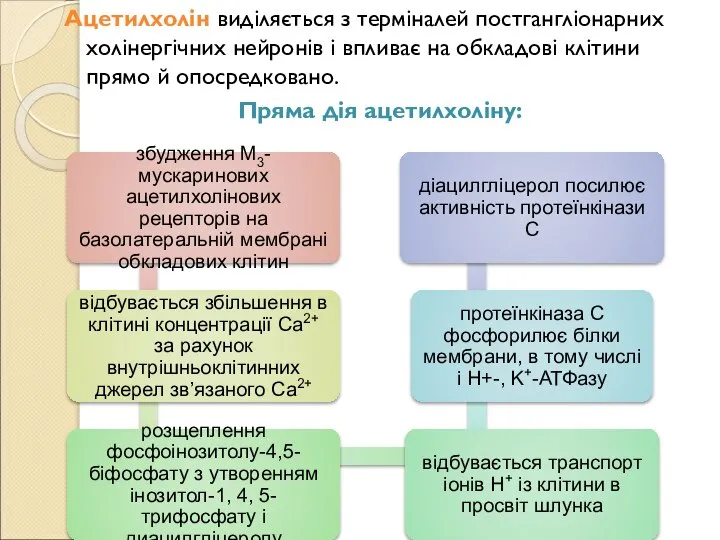
Слайд 123Непрямий вплив ацетилхоліну реалізується через його дію на ECL- клітини і G-
Непрямий вплив ацетилхоліну реалізується через його дію на ECL- клітини і G-

Слайд 124Феномен потенціації
Ефект дії між гістаміном і гастрином, між гістаміном і ацетилхоліном,
Феномен потенціації
Ефект дії між гістаміном і гастрином, між гістаміном і ацетилхоліном,

Слайд 125Роль соляної кислоти у травленні:
– сприяє перетворенню пепсиногену в пепсин шляхом дисоціації
Роль соляної кислоти у травленні:
– сприяє перетворенню пепсиногену в пепсин шляхом дисоціації

Слайд 126Патології, викликані порушеннями секреції соляної кислоти
Гіперсекреція соляної кислоти є однією з важливих
Патології, викликані порушеннями секреції соляної кислоти
Гіперсекреція соляної кислоти є однією з важливих

Слайд 127РУХОВА АКТИВНІСТЬ ШЛУНКА І ЕВАКУАЦІЯ ЙОГО ВМІСТУ
Шлунок виконує важливу транспортну функцію: накопичує,
РУХОВА АКТИВНІСТЬ ШЛУНКА І ЕВАКУАЦІЯ ЙОГО ВМІСТУ
Шлунок виконує важливу транспортну функцію: накопичує,

Слайд 128Стінка дна шлунка складається з трьох шарів:
зовнішнього шару поздовжніх м'язів,
середнього
Стінка дна шлунка складається з трьох шарів:
зовнішнього шару поздовжніх м'язів,
середнього
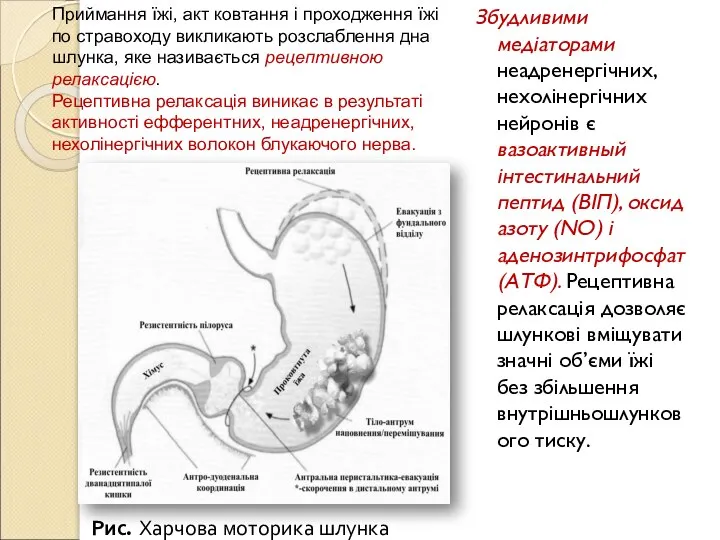
Слайд 129Збудливими медіаторами неадренергічних, нехолінергічних нейронів є вазоактивный інтестинальний пептид (ВІП), оксид азоту
Збудливими медіаторами неадренергічних, нехолінергічних нейронів є вазоактивный інтестинальний пептид (ВІП), оксид азоту
Слайд 130Властивість шлунка зберігати постійний рівень внутрішньопорожнинного тиску, незважаючи на прогресивне збільшення або
Властивість шлунка зберігати постійний рівень внутрішньопорожнинного тиску, незважаючи на прогресивне збільшення або

Слайд 131На відміну від пластичного тонусу, тонічні хвилі є фазними скороченнями, сила і
На відміну від пластичного тонусу, тонічні хвилі є фазними скороченнями, сила і

Слайд 132Рис. Пейсмекерні потенціали шлунка (повільні хвилі):
1 – 3 скорочення за хвилину,
Рис. Пейсмекерні потенціали шлунка (повільні хвилі):
1 – 3 скорочення за хвилину,
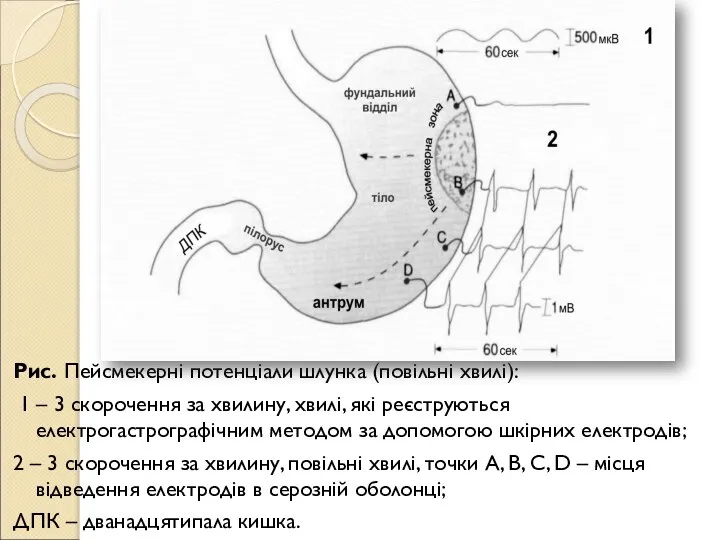
Слайд 133Пейсмекерні потенціали:
контролюють в шлунку кількість скорочень за хвилину;
впливають на поріг скорочень циркулярних
Пейсмекерні потенціали:
контролюють в шлунку кількість скорочень за хвилину;
впливають на поріг скорочень циркулярних

Слайд 134Рис. Перистальтичні скорочення шлунка:
1 – 3 скорочення в хвилину, хвилі, які реєструються
Рис. Перистальтичні скорочення шлунка:
1 – 3 скорочення в хвилину, хвилі, які реєструються
Слайд 135Скорочення виникає в результаті надходження імпульсів в антрум, але не кожна хвиля
Скорочення виникає в результаті надходження імпульсів в антрум, але не кожна хвиля

Слайд 136Частота появи перистальтичної хвилі у шлунку відносно стала серед представників одного виду.
Частота появи перистальтичної хвилі у шлунку відносно стала серед представників одного виду.

Слайд 137Внутрішньопорожнинний тиск у пілоричному відділі шлунка в стані спокою в голодних собак
Внутрішньопорожнинний тиск у пілоричному відділі шлунка в стані спокою в голодних собак

Слайд 138Перистальтичні хвилі виникають і передаються убік пілоруса. Пілорус є воротами шлунка і
Перистальтичні хвилі виникають і передаються убік пілоруса. Пілорус є воротами шлунка і

Слайд 139Коли хімус готовий до евакуації, кожна перистальтична хвиля евакуює 3-4 мол химуса
Коли хімус готовий до евакуації, кожна перистальтична хвиля евакуює 3-4 мол химуса

Слайд 140Концепція, що пояснює, чому ефективному стисканню піддаються лише великі частки їжі: вони
Концепція, що пояснює, чому ефективному стисканню піддаються лише великі частки їжі: вони

Слайд 141Рідини евакуюються зі шлунка значно швидше, ніж тверда їжа, яка звичайно залишається
Рідини евакуюються зі шлунка значно швидше, ніж тверда їжа, яка звичайно залишається

Слайд 142У багатьох роботах, присвячених фізіології шлунка, показано, що скорочення дна шлунка обумовлює
У багатьох роботах, присвячених фізіології шлунка, показано, що скорочення дна шлунка обумовлює

Слайд 143Довгий час наявність градієнта тиску між шлунком і дванадцятипалою кишкою, що є
Довгий час наявність градієнта тиску між шлунком і дванадцятипалою кишкою, що є

Слайд 144На швидкість евакуації рідин впливає їхня кількість і хімічний склад. Збільшення усередині
На швидкість евакуації рідин впливає їхня кількість і хімічний склад. Збільшення усередині

Слайд 145Уповільнення евакуації зі шлунка не завжди здатне вберегти від потрапляння в дванадцятипалу
Уповільнення евакуації зі шлунка не завжди здатне вберегти від потрапляння в дванадцятипалу

Слайд 146Структура і функції рецепторів кишки, зв'язаних з евакуаторною функцією шлунка складні.
Існують
Структура і функції рецепторів кишки, зв'язаних з евакуаторною функцією шлунка складні.
Існують

Слайд 147Під час відсутності імпульсів від блукаючого нерва (наприклад, при його перерізанні) перистальтика
Під час відсутності імпульсів від блукаючого нерва (наприклад, при його перерізанні) перистальтика

Слайд 148Уповільнення евакуації їжі зі шлунка називається гастропарезом. Він може виникнути внаслідок порушення
Уповільнення евакуації їжі зі шлунка називається гастропарезом. Він може виникнути внаслідок порушення

Слайд 149ТРАВЛЕННЯ В ТОНКОМУ КИШЕЧНИКУ
Ферментативне перетравлення їжі (корму) завершується в тонкому кишечнику. Всі
ТРАВЛЕННЯ В ТОНКОМУ КИШЕЧНИКУ
Ферментативне перетравлення їжі (корму) завершується в тонкому кишечнику. Всі

Слайд 150СЕКРЕЦІЯ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Підшлункова залоза синтезує і секретує у дванадцятипалу кишку більш 25
СЕКРЕЦІЯ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Підшлункова залоза синтезує і секретує у дванадцятипалу кишку більш 25

Слайд 151Характеристика панкреатичного соку: не має запаху, ізоосмотичний плазмі крові, його питома вага
Характеристика панкреатичного соку: не має запаху, ізоосмотичний плазмі крові, його питома вага

Слайд 152Основні катіони підшлункового соку: Na+, концентрація якого в соці відрізняється від концентрації
Основні катіони підшлункового соку: Na+, концентрація якого в соці відрізняється від концентрації

Слайд 153В основі секреції електролітів лежить два активних транспортних механізми:
пов'язаний з транспортом натрію
В основі секреції електролітів лежить два активних транспортних механізми:
пов'язаний з транспортом натрію

Слайд 154Підшлункова залоза синтезує і виділяє багато ферментів, що беруть участь у розкладанні
Підшлункова залоза синтезує і виділяє багато ферментів, що беруть участь у розкладанні

Слайд 155ФЕРМЕНТИ ПІДШЛУНКОВОГО СОКУ
Протеолітичні ферменти синтезуються в ацинарних клітинах у неактивній формі —
ФЕРМЕНТИ ПІДШЛУНКОВОГО СОКУ
Протеолітичні ферменти синтезуються в ацинарних клітинах у неактивній формі —
Слайд 156Процеси активації є високоспецифічними й в основному складаються з розщеплення пептидного зв'язку
Процеси активації є високоспецифічними й в основному складаються з розщеплення пептидного зв'язку

Слайд 157Еластаза (панкреатопептидаза) має меншу специфічність, ніж два попередні ферменти. Kарбоксипептидази гідролізують С-кінцеві
Еластаза (панкреатопептидаза) має меншу специфічність, ніж два попередні ферменти. Kарбоксипептидази гідролізують С-кінцеві

Слайд 158Один з цих білків— інгібітор Kазаля — активний тільки стосовно трипсину. Він
Один з цих білків— інгібітор Kазаля — активний тільки стосовно трипсину. Він

Слайд 159РЕГУЛЯЦІЯ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Регуляція поживними речовинами
Підшлункова залоза людини і тварин бере участь
РЕГУЛЯЦІЯ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Регуляція поживними речовинами
Підшлункова залоза людини і тварин бере участь

Слайд 160Рис. Секреція панкреатичного соку пішлункової залози собаки в залежності від характеру харчування:
а
Рис. Секреція панкреатичного соку пішлункової залози собаки в залежності від характеру харчування:
а
Слайд 161Склад ферментів соку також залежить від складу їжі. У соці, виділеному на
Склад ферментів соку також залежить від складу їжі. У соці, виділеному на

Слайд 162Виробляється така реакція (зміни ферментативного складу соку) на харчові речовини протягом індивідуального
Виробляється така реакція (зміни ферментативного складу соку) на харчові речовини протягом індивідуального

Слайд 163Ферментативна адаптація підшлункової залози починає розвиватися з перших днів змін в харчуванні
Ферментативна адаптація підшлункової залози починає розвиватися з перших днів змін в харчуванні

Слайд 164У собак здатність підшлункової залози адаптуватися до характеру їжі різко знижується при
У собак здатність підшлункової залози адаптуватися до характеру їжі різко знижується при

Слайд 165Таким чином, зміна секреції ферментів відповідно до вмісту харчових речовин у кормі
Таким чином, зміна секреції ферментів відповідно до вмісту харчових речовин у кормі

Слайд 166НЕРВОВИЙ КОНТРОЛЬ ПАНКРЕАТИЧНОЇ СЕКРЕЦІЇ
Більш 100 років тому І.П.Павлов вперше встановив, що блукаючі
НЕРВОВИЙ КОНТРОЛЬ ПАНКРЕАТИЧНОЇ СЕКРЕЦІЇ
Більш 100 років тому І.П.Павлов вперше встановив, що блукаючі

Слайд 167Антихолінергичні речовини (атропін, бантін, пробантін, ізопропамід) справляють гальмівний ефект на секрецію ферментів
Антихолінергичні речовини (атропін, бантін, пробантін, ізопропамід) справляють гальмівний ефект на секрецію ферментів

Слайд 168Наявність першої фази секреції підшлункової залози свідчить про участь вищих відділів ЦНС
Наявність першої фази секреції підшлункової залози свідчить про участь вищих відділів ЦНС


Слайд 170
Шлункова фаза секреції підшлункової залози починається при надходженні їжі в шлунок, має
Шлункова фаза секреції підшлункової залози починається при надходженні їжі в шлунок, має

Слайд 171У кишковій фазі секреції нейрогуморальні впливи на підшлункову залозу сприяють більш інтенсивній
У кишковій фазі секреції нейрогуморальні впливи на підшлункову залозу сприяють більш інтенсивній

Слайд 172Ваготомія й атропін знижують секрецію ферментів, стимульовану невеликим навантаженням амінокислотами і жирними
Ваготомія й атропін знижують секрецію ферментів, стимульовану невеликим навантаженням амінокислотами і жирними

Слайд 173В усіх фазах травлення закиснення дванадцятипалої кишки стимулює виділення секретину. Присутність жовчі,
В усіх фазах травлення закиснення дванадцятипалої кишки стимулює виділення секретину. Присутність жовчі,
Слайд 174ГУМОРАЛЬНІ МЕХАНІЗМИ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Гуморальні механізми забезпечують функціональну єдність усіх компонентів гастропанкреодуоденальної
ГУМОРАЛЬНІ МЕХАНІЗМИ СЕКРЕЦІЇ ПІДШЛУНКОВОЇ ЗАЛОЗИ Гуморальні механізми забезпечують функціональну єдність усіх компонентів гастропанкреодуоденальної

Слайд 175Основна маса ендокринних клітин підшлункової залози зосереджена в острівцях Лангерганса. Частина таких
Основна маса ендокринних клітин підшлункової залози зосереджена в острівцях Лангерганса. Частина таких

Слайд 176Однак на підшлункову залозу впливають гормони й інших ендокринних клітин й органів.
Однак на підшлункову залозу впливають гормони й інших ендокринних клітин й органів.

Слайд 177Секретин виділяється S-клітинами слизової оболонки тонкого кишечнику і стимулює клітини, що вистилають
Секретин виділяється S-клітинами слизової оболонки тонкого кишечнику і стимулює клітини, що вистилають

Слайд 178Тироліберин-рилізинг-фактор гіпоталамуса гальмує секрецію підшлункової залози. У ранньому неонатальному періоді його в
Тироліберин-рилізинг-фактор гіпоталамуса гальмує секрецію підшлункової залози. У ранньому неонатальному періоді його в

Слайд 179Соматостатин також проявляє різнонаправлені ефекти на підшлункову залозу, можливо, тому, що соматостатинсекретуючі
Соматостатин також проявляє різнонаправлені ефекти на підшлункову залозу, можливо, тому, що соматостатинсекретуючі

Слайд 180Соматостатин діє на ЦНС (гальмує нейронну активність і виділення ацетилхоліну, є агоністом
Соматостатин діє на ЦНС (гальмує нейронну активність і виділення ацетилхоліну, є агоністом

Слайд 181Енкефаліни мають широкий спектр дії і, крім участі в больовій чутливості і
Енкефаліни мають широкий спектр дії і, крім участі в больовій чутливості і

Слайд 182Введення лей-енкефаліна в периферичний кровотік знижує кількість панкреатичного соку в два рази.
Введення лей-енкефаліна в периферичний кровотік знижує кількість панкреатичного соку в два рази.

Слайд 183Бомбезин, що знаходиться в нервовій тканині, в епітеліальних клітинах кишки і шлунка,
Бомбезин, що знаходиться в нервовій тканині, в епітеліальних клітинах кишки і шлунка,

Слайд 184Гастрин за структурою близький до холецистокініну, але має більш слабку дію на
Гастрин за структурою близький до холецистокініну, але має більш слабку дію на

Слайд 185Панкреатичний поліпептид (ПП) виявлений у ендо- і екзокринних клітинах підшлункової залози і,
Панкреатичний поліпептид (ПП) виявлений у ендо- і екзокринних клітинах підшлункової залози і,

Слайд 186Ацетилхолін. Механізм цієї активації — звільнення іонів кальцію з ендоплазматичного ретикулума з
Ацетилхолін. Механізм цієї активації — звільнення іонів кальцію з ендоплазматичного ретикулума з

Слайд 187У механізмах регуляції секреції підшлункової залози важливе місце належить неорганічним іонам, у
У механізмах регуляції секреції підшлункової залози важливе місце належить неорганічним іонам, у

Слайд 188Рис. Схема спряження „стимул-секреція” в ацинарних клітинах підшлункової залози:
АХ – ацетилхолін; ГЗП
Рис. Схема спряження „стимул-секреція” в ацинарних клітинах підшлункової залози:
АХ – ацетилхолін; ГЗП
Слайд 189У випадку ацинарних клітин підшлункової залози миші і щура, у яких не
У випадку ацинарних клітин підшлункової залози миші і щура, у яких не
Слайд 190ВЗАЄМОДІЯ ЕКЗО- І ЕНДОКРИННИХ ФУНКЦІЙ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Перше зазначення існування екзо- і ендокринних
ВЗАЄМОДІЯ ЕКЗО- І ЕНДОКРИННИХ ФУНКЦІЙ ПІДШЛУНКОВОЇ ЗАЛОЗИ
Перше зазначення існування екзо- і ендокринних

Слайд 191Також, ендокринна функція підшлункової залози залежить від її зовнішньосекреторної активності. Про це
Також, ендокринна функція підшлункової залози залежить від її зовнішньосекреторної активності. Про це

Слайд 192ЖОВЧОУТВОРЮВАЛЬНА ФУНКЦІЯ ПЕЧІНКИ
Печінка виконує в організмі важливі функції, серед яких:
участь у
ЖОВЧОУТВОРЮВАЛЬНА ФУНКЦІЯ ПЕЧІНКИ
Печінка виконує в організмі важливі функції, серед яких:
участь у

Слайд 193Бар'єрна функція печінки складається в детоксикації (знешкодженні) продуктів обміну (наприклад, продуктів розпаду
Бар'єрна функція печінки складається в детоксикації (знешкодженні) продуктів обміну (наприклад, продуктів розпаду

Слайд 194СКЛАД, ВЛАСТИВОСТІ Й УТВОРЕННЯ ЖОВЧІ
Таблиця.
Склад, pH та питома вага печінкової та
СКЛАД, ВЛАСТИВОСТІ Й УТВОРЕННЯ ЖОВЧІ
Таблиця.
Склад, pH та питома вага печінкової та

Слайд 195Через присутність білірубіна (продукт розпаду гемоглобіну) жовч має золотостий колір.
Склад електролітів
Через присутність білірубіна (продукт розпаду гемоглобіну) жовч має золотостий колір.
Склад електролітів

Слайд 196У печінці жовчні кислоти знаходяться у виді кон’югатів, в основному, з гліцином
У печінці жовчні кислоти знаходяться у виді кон’югатів, в основному, з гліцином
Слайд 197Завдяки амфіпатичним властивостям, жовчні кислоти, їхні солі і кон’югати є ефективними детергентами
Завдяки амфіпатичним властивостям, жовчні кислоти, їхні солі і кон’югати є ефективними детергентами

Слайд 198Гідрофільні (гідроксильні і карбоксильні) групи молекул жовчних кислот знаходяться на одній стороні
Гідрофільні (гідроксильні і карбоксильні) групи молекул жовчних кислот знаходяться на одній стороні

Слайд 199Міцели часто містять і холестерол, і фосфоліпіди (змішані міцели). Якщо концентрація холестерола
Міцели часто містять і холестерол, і фосфоліпіди (змішані міцели). Якщо концентрація холестерола

Слайд 200За добу печінка дорослої людини секретує 600–1200 мл (зі швидкістю близько 0,4-0,8
За добу печінка дорослої людини секретує 600–1200 мл (зі швидкістю близько 0,4-0,8

Слайд 201У жовчі людини, собаки і кішки переважає оранжево-коричневий пігмент білірубін; у жовчі
У жовчі людини, собаки і кішки переважає оранжево-коричневий пігмент білірубін; у жовчі

Слайд 202Рис. Білірубін
До складу жовчі входять також:
ферменти (амілази, фосфатази, протеази, каталази, оксидази),
Рис. Білірубін
До складу жовчі входять також:
ферменти (амілази, фосфатази, протеази, каталази, оксидази),
Слайд 203СЕКРЕЦІЯ І ФУНКЦІЇ ЖОВЧІ
Близько 70% добової жовчі надходить з канальців печінки, а
СЕКРЕЦІЯ І ФУНКЦІЇ ЖОВЧІ
Близько 70% добової жовчі надходить з канальців печінки, а

Слайд 205Близько 10% (іноді до 20%) жовчних кислот виводиться з організму. Жовчні кислоти
Близько 10% (іноді до 20%) жовчних кислот виводиться з організму. Жовчні кислоти

Слайд 206Білірубін (як і інші жовчні пігменти) реабсорбується в жовчному міхурі і кишечнику
Білірубін (як і інші жовчні пігменти) реабсорбується в жовчному міхурі і кишечнику

Слайд 207Рис. Реабсорбція і секреція у жовчному міхурі
Рис. Реабсорбція і секреція у жовчному міхурі
Слайд 208У кишечнику жовч:
сприяє розщепленню, омиленню, емульгуванню й усмоктуванню жирів,
підсилює перистальтику
У кишечнику жовч:
сприяє розщепленню, омиленню, емульгуванню й усмоктуванню жирів,
підсилює перистальтику

Слайд 209РЕГУЛЯЦІЯ СЕКРЕЦІЇ ЖОВЧІ
Секреція жовчі відбувається постійно, але між прийомами їжі вона накопичується
РЕГУЛЯЦІЯ СЕКРЕЦІЇ ЖОВЧІ
Секреція жовчі відбувається постійно, але між прийомами їжі вона накопичується

Слайд 210Рис. Механізми наповнення (А) та випорожнення (Б) жовчного міхура
Рис. Механізми наповнення (А) та випорожнення (Б) жовчного міхура
Слайд 211Ритмічні скорочення (2–6 у хв) спостерігаються також під час травлення і сприяють
Ритмічні скорочення (2–6 у хв) спостерігаються також під час травлення і сприяють

Слайд 212Рух жовчі до дванадцятипалої кишки відбувається відповідно до законів гідродинаміки вбік найменшого
Рух жовчі до дванадцятипалої кишки відбувається відповідно до законів гідродинаміки вбік найменшого

Слайд 213Регулятори жовчоутворювальної і жовчосекреторної функції печінки: соматостатин, вазопресин, окситоцин, енкефаліни, субстанція П,
Регулятори жовчоутворювальної і жовчосекреторної функції печінки: соматостатин, вазопресин, окситоцин, енкефаліни, субстанція П,

Слайд 214ВІП є стимулятором секреції води і бікарбонатів холангіоцитами (епітеліальними клітинами жовчних проток)
ВІП є стимулятором секреції води і бікарбонатів холангіоцитами (епітеліальними клітинами жовчних проток)

Слайд 215Гормони щитовидної залози також регулюють жовчосекреторну функцію печінки.
Наприклад, тироксин і трийодтиронін
Гормони щитовидної залози також регулюють жовчосекреторну функцію печінки.
Наприклад, тироксин і трийодтиронін

Слайд 216Нервові шляхи, по яких до печінки надходять стимулюючі і гальмуючі імпульси, представлені
Нервові шляхи, по яких до печінки надходять стимулюючі і гальмуючі імпульси, представлені

Слайд 217Рис. Шляхи перетворення глюкози в печінці.
МЕТАБОЛІЧНІ ФУНКЦІЇ ПЕЧІНКИ
Рис. Шляхи перетворення глюкози в печінці.
МЕТАБОЛІЧНІ ФУНКЦІЇ ПЕЧІНКИ
Слайд 218Рис. Шляхи перетворення амінокислот в печінці.
Рис. Шляхи перетворення амінокислот в печінці.
Слайд 219Рис. Шляхи перетворення жирних кислот в печінці
Рис. Шляхи перетворення жирних кислот в печінці
Слайд 220Крім перетворення і розподілу вуглеводів, жирів і амінокислот, урівноваження коливань в обміні
Крім перетворення і розподілу вуглеводів, жирів і амінокислот, урівноваження коливань в обміні

Слайд 221КИШКОВИЙ СІК: СКЛАД, ВЛАСТИВОСТІ, РЕГУЛЯЦІЯ СОКОВИДІЛЕННЯ
Найважливіші кишкові ферменти у великих кількостях зосереджені
КИШКОВИЙ СІК: СКЛАД, ВЛАСТИВОСТІ, РЕГУЛЯЦІЯ СОКОВИДІЛЕННЯ
Найважливіші кишкові ферменти у великих кількостях зосереджені

 Височно-нижнечелюстной сустав
Височно-нижнечелюстной сустав Постиндустриальный этап развития цивилизации
Постиндустриальный этап развития цивилизации Анатомия полушарий головного мозга человека
Анатомия полушарий головного мозга человека Органы чувств. Анализаторы
Органы чувств. Анализаторы Көбелек сабағы
Көбелек сабағы Таблицы по зоологии (часть 1)
Таблицы по зоологии (часть 1) Животные Севера
Животные Севера Радула молюсків та її будова
Радула молюсків та її будова Фазы митоза
Фазы митоза Индивидуальное развитие организмов
Индивидуальное развитие организмов Healthy Lifestyle
Healthy Lifestyle Экологическая викторина О птицах в смокингах
Экологическая викторина О птицах в смокингах Ботанический сад имени профессора Б.М. Козо-Полянского
Ботанический сад имени профессора Б.М. Козо-Полянского Коэволюция
Коэволюция Group of marine microbial
Group of marine microbial Почему кровь красная?
Почему кровь красная? Элемент металлического происхождения марганец
Элемент металлического происхождения марганец Лишайники. Биологический диктант
Лишайники. Биологический диктант Воздух в жизни растений 6 класс
Воздух в жизни растений 6 класс Соцветия
Соцветия Соболь – дорогой зверёк
Соболь – дорогой зверёк Транспортная функция плазмалеммы
Транспортная функция плазмалеммы Кроссворд. Типы: Плоские, Круглые, Кольчатые черви
Кроссворд. Типы: Плоские, Круглые, Кольчатые черви Любимый огород
Любимый огород Строение и работа сердца
Строение и работа сердца Реликтовые растения Забайкальского края
Реликтовые растения Забайкальского края خمسة أنواع من الماشية
خمسة أنواع من الماشية Строение и функции листа
Строение и функции листа