- Закономерности роста и размножения клеток. Скорость растущей биомассы клеток и даваемый ею прирост

Содержание
- 2. СКОРОСТЬ РОСТА КЛЕТОК При исследовании ростовых процессов нужно различать растущую биомассу клеток и даваемой ею прирост.
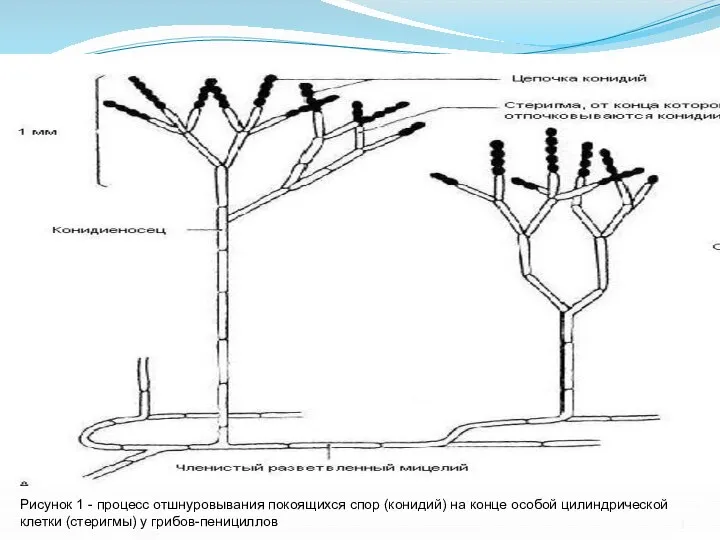
- 3. Рисунок 1 - процесс отшнуровывания покоящихся спор (конидий) на конце особой цилиндрической клетки (стеригмы) у грибов-пенициллов
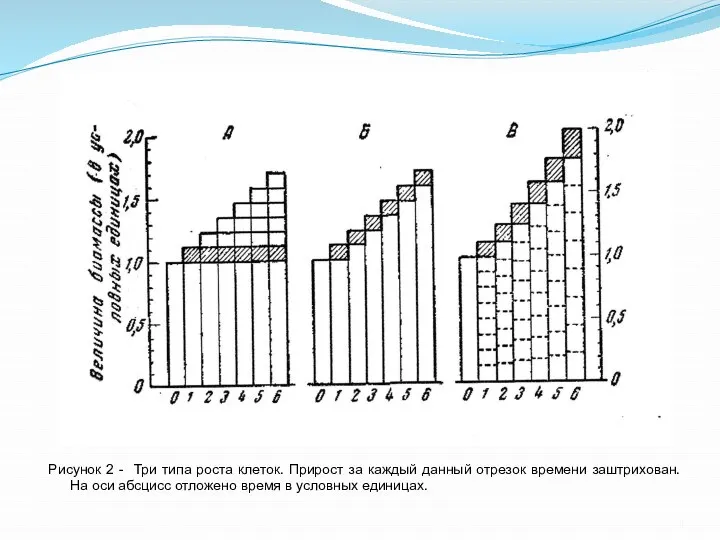
- 4. Рисунок 2 - Три типа роста клеток. Прирост за каждый данный отрезок времени заштрихован. На оси
- 5. Для количественной характеристики ростовых процессов пользуются двумя показателями: 1) абсолютной (валовой) и 2) относительной (удельной) скоростью
- 7. Средняя удельная скорость роста (μср) за какой-либо период времени t1 - t0 равна:
- 8. Относительная скорость роста характеризуется увеличением натуральных логарифмов биомассы за единицу времени. Этим она отличается от валовой
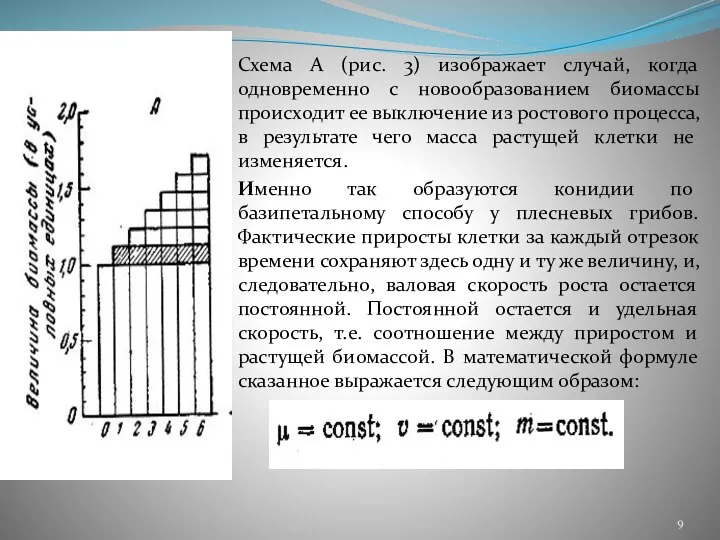
- 9. Схема А (рис. 3) изображает случай, когда одновременно с новообразованием биомассы происходит ее выключение из ростового
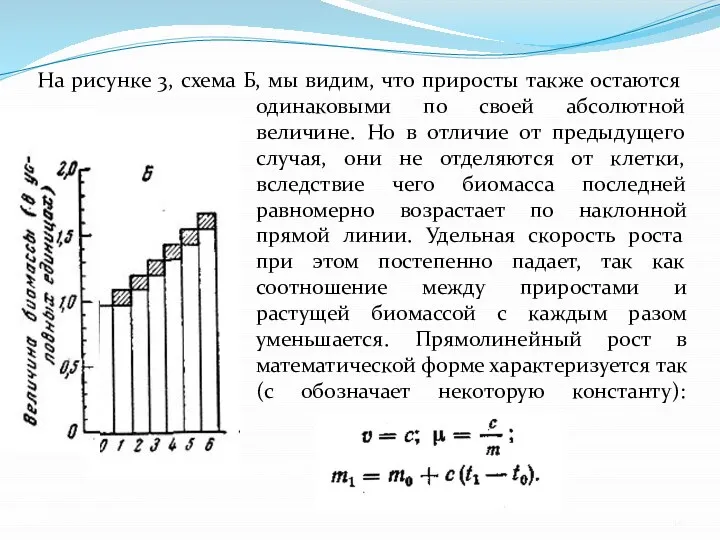
- 10. На рисунке 3, схема Б, мы видим, что приросты также остаются одинаковыми по своей абсолютной величине.
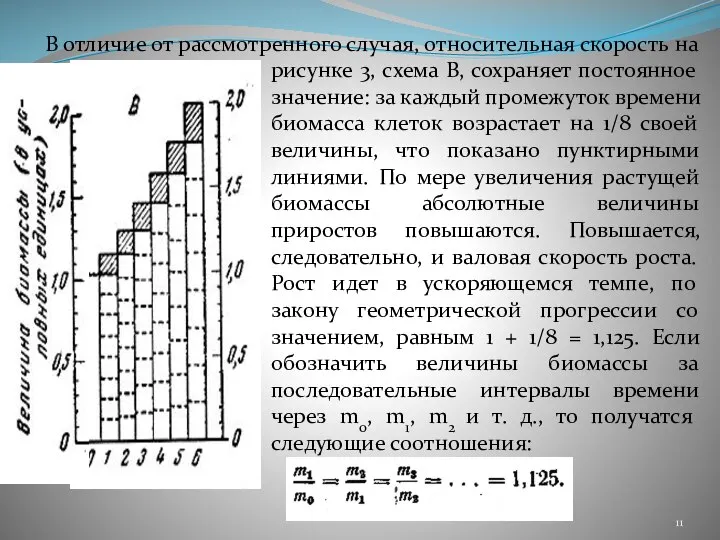
- 11. В отличие от рассмотренного случая, относительная скорость на рисунке 3, схема В, сохраняет постоянное значение: за
- 12. Рост с постоянной удельной скоростью в общей форме характеризуется так: Здесь с – некоторый постоянный коэффициент,
- 13. Необходимо подчеркнуть, что не всякий ускоряющийся рост подчиняется закону геометрической прогрессии. Это бывает только в том
- 14. Бактериальная клетка в физиологически активном состоянии растет всей массой своего тела (в этом состоит ее отличие
- 15. Иное наблюдается у шаровидных бактерий. На скорость их роста существенное влияние оказывает изменяющееся соотношение между поверхностью
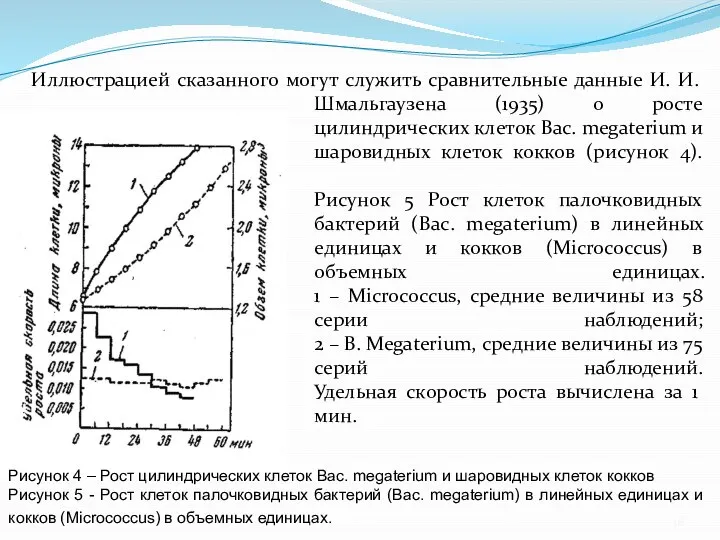
- 16. Иллюстрацией сказанного могут служить сравнительные данные И. И. Шмальгаузена (1935) о росте цилиндрических клеток Вac. megaterium
- 17. Рост клеток протозоа и водорослей обычно имеет такой же затухающий характер, как и у кокковых бактерий.
- 18. В случае постепенного падения удельной скорости роста в ростовом процессе может иметь замедляющийся, равномерный или даже
- 19. СКОРОСТЬ РАЗМНОЖЕНИЯ КЛЕТОК О скорости размножения одноклеточных организмов судят по тому, как часто они делятся или
- 20. Предположим, что вначале было N0 клеток. За каждую генерацию численность их повышается в 2 раза, а
- 21. Если, как было сказано выше, за отрезок времени t1 - t0, сменяется n клеточных поколений. То
- 22. В популяциях, где одновременно присутствуют клетки, находящиеся на разных ступенях индивидуального развития, средний вес одной клетки
- 23. На этом основании величины N1 и N0 можно заменить, соответственно, на m1 и m0 (коэффициент а
- 24. О скорости роста микроорганизмов принято судить по продолжительности генерации. Указанный прием достаточно обоснован, поскольку удельная скорость
- 25. Кроме того, нужно учитывать, что удвоение биомассы на протяжении одной генерации вовсе не означает, что ее
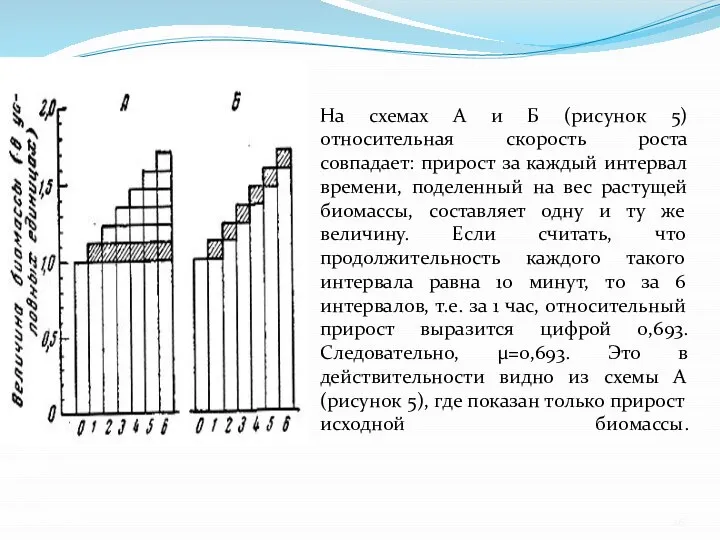
- 26. На схемах А и Б (рисунок 5) относительная скорость роста совпадает: прирост за каждый интервал времени,
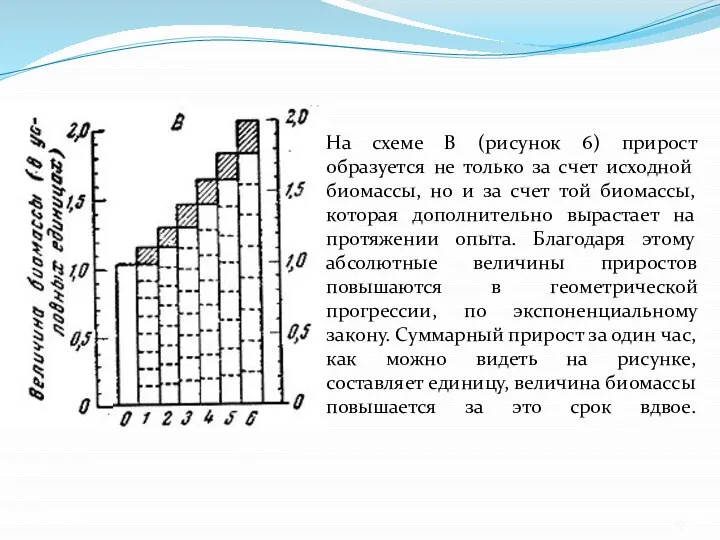
- 27. На схеме В (рисунок 6) прирост образуется не только за счет исходной биомассы, но и за
- 30. Скачать презентацию
Слайд 2СКОРОСТЬ РОСТА КЛЕТОК
При исследовании ростовых процессов нужно различать растущую биомассу клеток и
СКОРОСТЬ РОСТА КЛЕТОК
При исследовании ростовых процессов нужно различать растущую биомассу клеток и

Слайд 3Рисунок 1 - процесс отшнуровывания покоящихся спор (конидий) на конце особой цилиндрической
Рисунок 1 - процесс отшнуровывания покоящихся спор (конидий) на конце особой цилиндрической
Слайд 4Рисунок 2 - Три типа роста клеток. Прирост за каждый данный отрезок
Рисунок 2 - Три типа роста клеток. Прирост за каждый данный отрезок
Слайд 5Для количественной характеристики ростовых процессов пользуются двумя показателями:
1) абсолютной (валовой) и
2)
Для количественной характеристики ростовых процессов пользуются двумя показателями: 1) абсолютной (валовой) и 2)


Слайд 7Средняя удельная скорость роста (μср) за какой-либо период времени
t1 - t0
Средняя удельная скорость роста (μср) за какой-либо период времени t1 - t0

Слайд 8Относительная скорость роста характеризуется увеличением натуральных логарифмов биомассы за единицу времени.
Этим
Относительная скорость роста характеризуется увеличением натуральных логарифмов биомассы за единицу времени.
Этим

Слайд 9Схема А (рис. 3) изображает случай, когда одновременно с новообразованием биомассы происходит
Схема А (рис. 3) изображает случай, когда одновременно с новообразованием биомассы происходит
Слайд 10На рисунке 3, схема Б, мы видим, что приросты также остаются одинаковыми
На рисунке 3, схема Б, мы видим, что приросты также остаются одинаковыми
Слайд 11В отличие от рассмотренного случая, относительная скорость на рисунке 3, схема В,
В отличие от рассмотренного случая, относительная скорость на рисунке 3, схема В,
Слайд 12Рост с постоянной удельной скоростью в общей форме характеризуется так:
Здесь с –
Рост с постоянной удельной скоростью в общей форме характеризуется так: Здесь с –

Слайд 13 Необходимо подчеркнуть, что не всякий ускоряющийся рост подчиняется закону геометрической прогрессии.
Необходимо подчеркнуть, что не всякий ускоряющийся рост подчиняется закону геометрической прогрессии.

Слайд 14 Бактериальная клетка в физиологически активном состоянии растет всей массой своего тела
Бактериальная клетка в физиологически активном состоянии растет всей массой своего тела

Слайд 15 Иное наблюдается у шаровидных бактерий. На скорость их роста существенное влияние
Иное наблюдается у шаровидных бактерий. На скорость их роста существенное влияние

Слайд 16Иллюстрацией сказанного могут служить сравнительные данные И. И. Шмальгаузена (1935) о росте
Иллюстрацией сказанного могут служить сравнительные данные И. И. Шмальгаузена (1935) о росте
Слайд 17 Рост клеток протозоа и водорослей обычно имеет такой же затухающий характер,
Рост клеток протозоа и водорослей обычно имеет такой же затухающий характер,

Слайд 18 В случае постепенного падения удельной скорости роста в ростовом процессе может
В случае постепенного падения удельной скорости роста в ростовом процессе может

Слайд 19СКОРОСТЬ РАЗМНОЖЕНИЯ КЛЕТОК
О скорости размножения одноклеточных организмов судят по тому,
СКОРОСТЬ РАЗМНОЖЕНИЯ КЛЕТОК
О скорости размножения одноклеточных организмов судят по тому,

Слайд 20Предположим, что вначале было N0 клеток. За каждую генерацию численность их повышается
Предположим, что вначале было N0 клеток. За каждую генерацию численность их повышается

Слайд 21Если, как было сказано выше, за отрезок времени t1 - t0, сменяется
Если, как было сказано выше, за отрезок времени t1 - t0, сменяется

Слайд 22В популяциях, где одновременно присутствуют клетки, находящиеся на разных ступенях индивидуального развития,
В популяциях, где одновременно присутствуют клетки, находящиеся на разных ступенях индивидуального развития,

Слайд 23На этом основании величины N1 и N0 можно заменить, соответственно, на m1
На этом основании величины N1 и N0 можно заменить, соответственно, на m1

Слайд 24 О скорости роста микроорганизмов принято судить по продолжительности генерации.
Указанный
О скорости роста микроорганизмов принято судить по продолжительности генерации.
Указанный

Слайд 25 Кроме того, нужно учитывать, что удвоение биомассы на протяжении одной генерации
Кроме того, нужно учитывать, что удвоение биомассы на протяжении одной генерации

Слайд 26
На схемах А и Б (рисунок 5) относительная скорость роста совпадает: прирост
На схемах А и Б (рисунок 5) относительная скорость роста совпадает: прирост
Слайд 27На схеме В (рисунок 6) прирост образуется не только за счет исходной
На схеме В (рисунок 6) прирост образуется не только за счет исходной

 Самые-самые растения
Самые-самые растения Экосистемы. Повторение
Экосистемы. Повторение Класс пресмыкающиеся, особенности строения
Класс пресмыкающиеся, особенности строения Простейшие. Жгутиконосцы, инфузории
Простейшие. Жгутиконосцы, инфузории Функциональная организация генома
Функциональная организация генома Презентация на тему Паразиты человека
Презентация на тему Паразиты человека  Дыхание. Кислород. Окисление сложных органических веществ
Дыхание. Кислород. Окисление сложных органических веществ Полисахариды. Биология
Полисахариды. Биология Осенние цветы
Осенние цветы Презентация на тему Терморегуляция организма. Закаливание
Презентация на тему Терморегуляция организма. Закаливание  Эволюция пищеварительной системы
Эволюция пищеварительной системы Нервная система
Нервная система Возможности коры головного мозга человека. Работу выполнили: Новосёлов Артем Хохлов Саша Худяева Света
Возможности коры головного мозга человека. Работу выполнили: Новосёлов Артем Хохлов Саша Худяева Света Пролапс слезной железы третьего века
Пролапс слезной железы третьего века Под одной крышей с растениями. Месторасположение
Под одной крышей с растениями. Месторасположение Олигохеты, пиявки
Олигохеты, пиявки Класс Млекопитающие
Класс Млекопитающие Дыхательная система. Полость носа, гортань, трахея
Дыхательная система. Полость носа, гортань, трахея Строение и работа сердца человека
Строение и работа сердца человека Зимующие птицы
Зимующие птицы Индивидуальное развитие и рост растений
Индивидуальное развитие и рост растений Презентация на тему Отряд Таракановые
Презентация на тему Отряд Таракановые  Функции жиров
Функции жиров Приспособленность растений при пустынлевой сушливости
Приспособленность растений при пустынлевой сушливости Покрытосеменные растения. Двудольные растения. Магнолииды
Покрытосеменные растения. Двудольные растения. Магнолииды Учение И.П. Павлова о высшей нервной деятельности и ее типологических особенностях
Учение И.П. Павлова о высшей нервной деятельности и ее типологических особенностях Цитология
Цитология Некоторые важнейшие ароморфозы
Некоторые важнейшие ароморфозы