- Биохимия соединительной ткани

Содержание
- 2. К соединительной ткани относят ткани костей, зубов, хрящей, сухожилий, подкожную клетчатку. Соединительная ткань может образовывать твердые
- 3. Особенностью строения соединительной ткани является наличие хорошо развитого внеклеточного матрикса. Межклеточный матрикс занимает больше место, чем
- 4. Внеклеточный матрикс (ВКМ) соединительной ткани: 1. Имеет сложный химический состав, содержит разнообразные белки и полисахариды, в
- 5. Белки внеклеточного матрикса Среди белков внеклеточного матрикса выделяют белки двух функциональных типов - фибриллярные и адгезивные.
- 6. Полисахариды внеклеточного матрикса Полисахариды представлены гликозаминогликанами (ГАГ), которые различаются составом дисахаридов и их количеством. Могут быть
- 7. Полисахариды – гликозамингликаны и протеогликаны, а также и адгезивные белки составляют группу неколлагеновых белков межклеточного матрикса.
- 8. Коллаген – основной структурный белок соединительной ткани Коллагены составляют приблизительно 30 % общего количества белка в
- 9. Строение молекул коллагенов Молекулы коллагенов имеют трехспиральную структуру, полученную при скручивании трех полипептидных α – цепей,
- 10. Особенности аминокислотного состава коллагена Полипептидная цепь коллагена состоит из повторяющихся триплетов: [Гли-Х-Y], где Гли – глицин,
- 11. Схематически цепь коллагена может быть представлена следующим образом: Гли-Ала-ГиПро-Гли-Про-ГиЛиз-Гли-Ала-ГиПро Глицин обеспечивает плотность укладки трех полипептидных цепей
- 12. Синтез коллагена Коллаген синтезируется внутри различных клеток соединительной ткани в виде препроколлагена, содержащего на N –
- 13. Созревание коллагена (процессинг) После синтеза цепи коллагена следует сложный многоступенчатый процесс - созревания коллагена. Включает 2
- 14. Внутриклеточный этап созревание коллагена включает в себя ряд последовательных изменений цепи препроколлагена: 1) Отщепление сигнальной пептидной
- 15. 2) Гидроксилирование пролина и лизина проколлагена, катализируют ферментативные реакции - гидроксилазы Синтезированный гидроксипролин (ГиПро) участвует в
- 16. Ферментам гидроксилазам для осуществления реакции гидроксилирования необходим атом двухвалентного железа и его восстановитель - витамин С
- 17. 3) Гликозилирование гидроксилизина под действием гликозилтрансфераз (присоединение углеводного компонента) Углеводные компоненты связываются с гидроксилином О -
- 18. 4) Формирование тройной спирали проколлагена Каждая α – цепь проколлагена соединяется водородными связями с двумя другими
- 19. Внеклеточный этап созревания включает в себя ряд последовательных изменений 1) Образование молекулы тропоколлагена. Отщепление от проколлагена
- 20. 2) Формирование фибрилл коллагена Из сформировавшихся молекул тропоколлагена происходит сборка коллагеновых фибрилл, в которых одна нить
- 21. 3) Стабилизация и укрепление фибрилл коллагена Модифицированные аминокислоты - гидроксипролин и гидроксилизин играют важную роль в
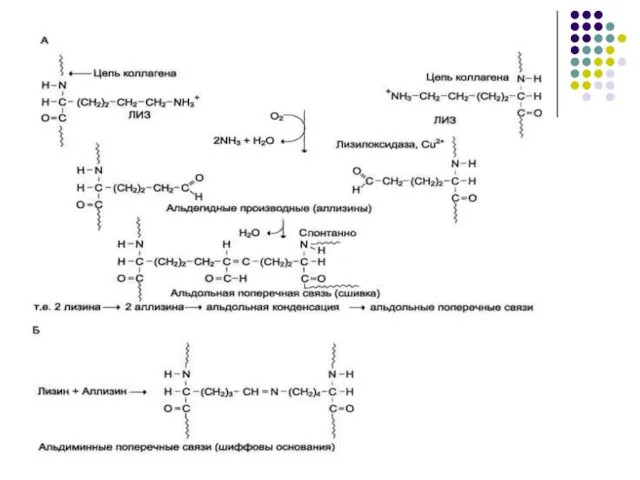
- 22. Образование аллизина Для создания укрепляющих связей фермент лизилоксидаза катализирует реакцию превращения в тропоколлагене аминогрупп отдельных лизильных
- 23. Возникшие альдегидные группы участвуют в образовании ковалентных связей, которые стабилизируют фибриллы коллагена. Альдольная сшивка – аллизин
- 25. На рисунке схематически представлены трехспиральные молекулы тропоколлагена после наложения внутри и межмолекулярных сшивок, которые изображены в
- 27. Коллагеновые фибриллы формируют коллагеновые волокны разной толщины. Фибриллы укладываются параллельно друг другу и далее объединяются в
- 28. Общая схема внеклеточного этапа созревания
- 29. Уникальные свойства коллагенов Коллагеновые волокна обладают огромной прочностью и практически нерастяжимы. Они могут выдерживать нагрузку, в
- 30. Катаболизм коллагена Распад коллагена происходит медленно под действием коллагеназ. Основной фермент - Са 2+, Zn 2+
- 31. Основной маркер распада коллагена Важнейший метаболитом характеризующим скорость распада коллагена является гидроксипролин. Повышение содержания гидроксипролина в
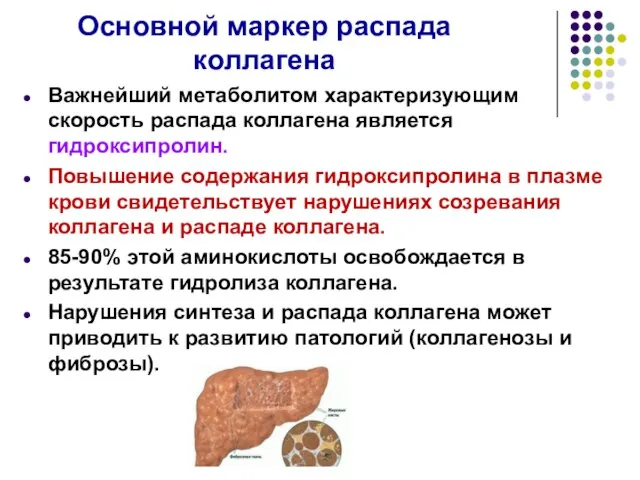
- 32. Типы коллагена В настоящее время известно около 20 различных типов коллагена, различающихся по первичной и пространственной
- 33. Наиболее распространенные типы коллагенов.
- 34. Эластин – это основной компонент эластических волокон Содержатся в тканях, обладающих значительной эластичностью - кровеносные сосуды,
- 35. Особенности аминокислотного состава эластина Эластин – гликопротеин с молекулярной массой 70кДа, содержит много гидрофобных аминокислот -
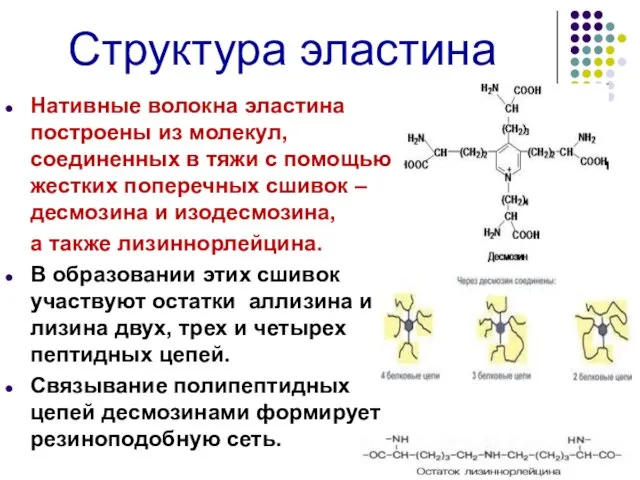
- 36. Структура эластина Нативные волокна эластина построены из молекул, соединенных в тяжи с помощью жестких поперечных сшивок
- 37. Неколлагеновые белки межклеточного матрикса К этой группе белков относятся относятся адгезивные белки, такие как фибронектин, ламинин,
- 38. Фибронектин состоят из 2-х идентичных цепей, содержащих по 7-8 доменов со специфичными центрами связывания: 1) коллагена
- 39. Адгезивные белки часто содержат последовательность Арг-Гли-Асп (RGD), участвующую в присоединении к клеточным белковым рецепторам – интегринам,
- 40. Ламинины – гликопротеины базальных мембран (3), отделяющую соединительную ткань от эпителия Содержит несколько центров связывания с
- 41. Ламинин – гликопротеин, состоит из трех полипептидных цепей (a и 2-х β), которые укладываются в пространстве
- 42. К неколлагеновым белкам относятся Gla-белки. (это белки, содержащие аминокислотные остатки 7-карбокси глутаминовой кислоты) Матриксный gla-белок содержит
- 43. К неколлагеновым белкам межклеточного матрикса относятся протеогликаны Молекулы протеогликанов участвуют в сборке межклеточного матрикса, облегчают фиксацию
- 44. Строение протеогликанов В структуре протеогликанов выделяют коровый (COR) белок (от анг. сore – основа, ядро), который
- 45. Основную часть протеогликанов составляют гликозамингликаны (ГАГ) Гликозаминогликаны – гетерополисахариды, состоящие из повторяющихся дисахаридов, в состав которых
- 46. Хондроитинсульфат построен из глюкуроновой кислоты и сульфатированного N–ацетилгалактозамина.
- 47. Дисахаридный фрагмент дерматансульфата.
- 48. Дисахариды кератинсульфата состоят из галактозы и сульфатированного N–ацетилглюкозамина.
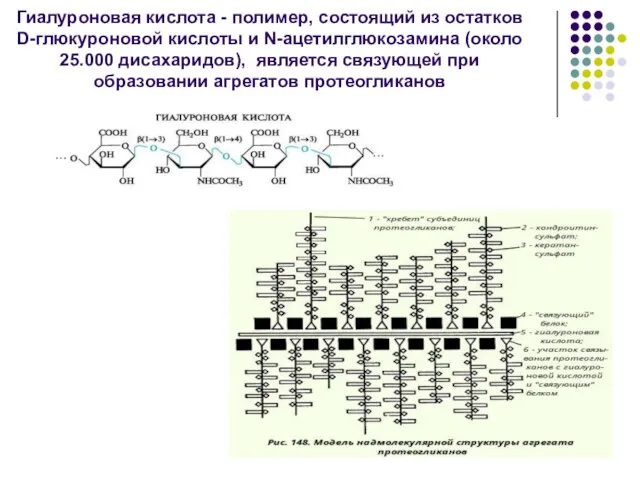
- 49. Гиалуроновая кислота - полимер, состоящий из остатков D-глюкуроновой кислоты и N-ацетилглюкозамина (около 25.000 дисахаридов), является связующей
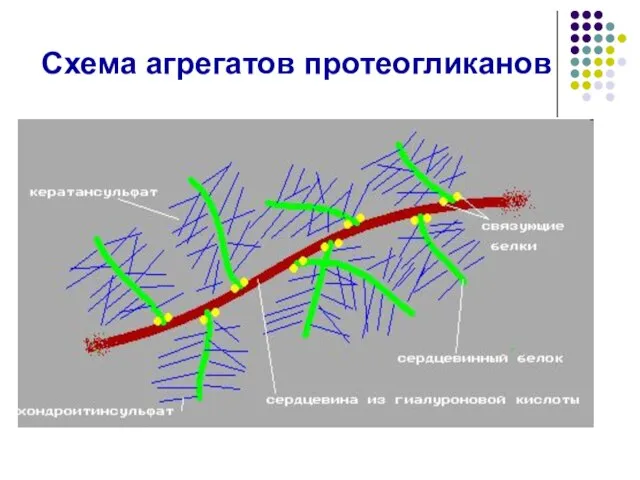
- 50. Схема агрегатов протеогликанов
- 51. Функции гликозаминогликанов - участвуют в организации межклеточного матрикса, являются основным скрепляющим веществом. - взаимодействуют с клеточными
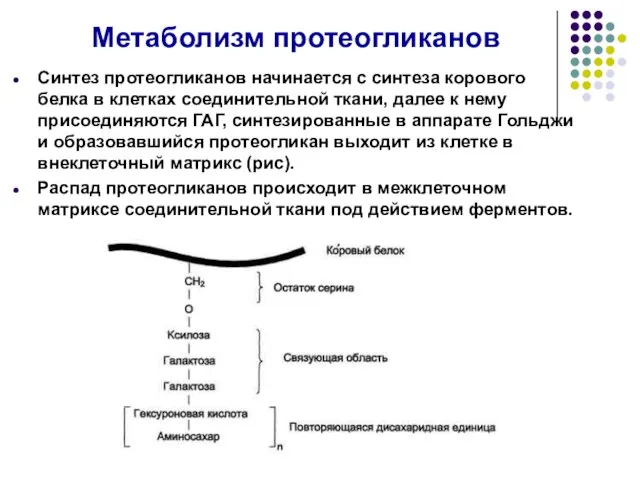
- 52. Метаболизм протеогликанов Синтез протеогликанов начинается с синтеза корового белка в клетках соединительной ткани, далее к нему
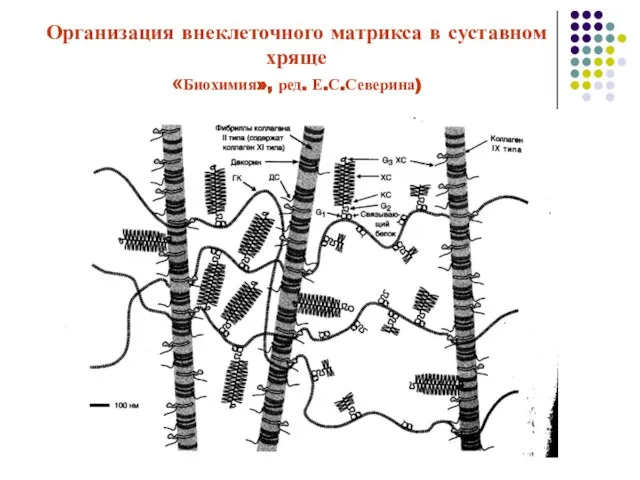
- 53. Организация внеклеточного матрикса в суставном хряще «Биохимия», ред. Е.С.Северина)
- 55. Скачать презентацию
Слайд 2
К соединительной ткани относят ткани костей, зубов, хрящей, сухожилий, подкожную клетчатку.
Соединительная ткань
К соединительной ткани относят ткани костей, зубов, хрящей, сухожилий, подкожную клетчатку.
Соединительная ткань
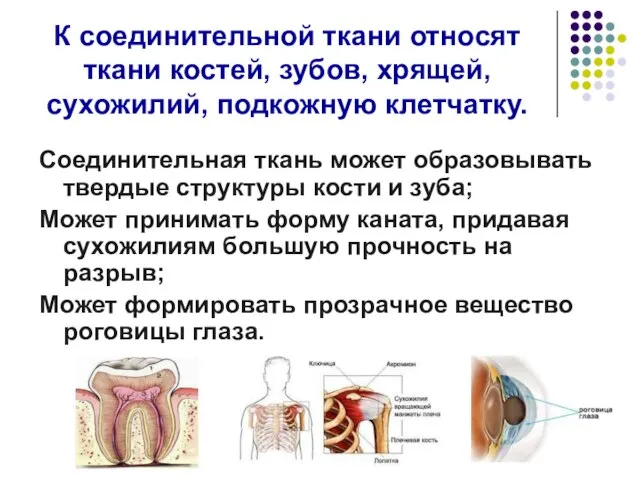
Слайд 3Особенностью строения соединительной ткани является наличие хорошо развитого внеклеточного матрикса. Межклеточный матрикс
Особенностью строения соединительной ткани является наличие хорошо развитого внеклеточного матрикса. Межклеточный матрикс
Слайд 4Внеклеточный матрикс (ВКМ) соединительной ткани:
1. Имеет сложный химический состав, содержит разнообразные белки
Внеклеточный матрикс (ВКМ) соединительной ткани:
1. Имеет сложный химический состав, содержит разнообразные белки

Слайд 5Белки внеклеточного матрикса
Среди белков внеклеточного матрикса выделяют белки двух функциональных типов
Белки внеклеточного матрикса
Среди белков внеклеточного матрикса выделяют белки двух функциональных типов

Слайд 6Полисахариды
внеклеточного матрикса
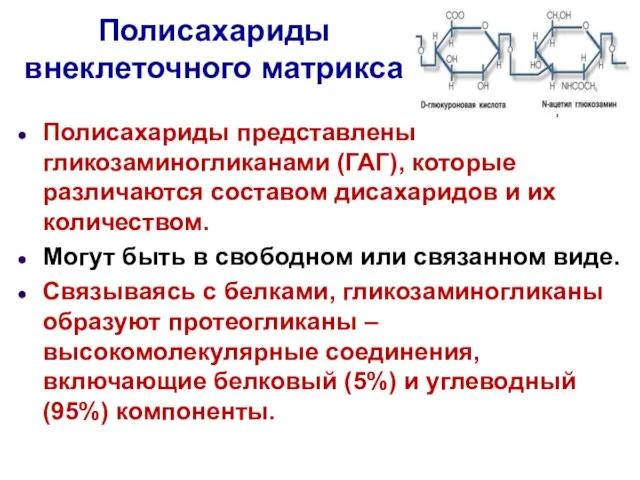
Полисахариды представлены гликозаминогликанами (ГАГ), которые различаются составом дисахаридов и их
Полисахариды
внеклеточного матрикса
Полисахариды представлены гликозаминогликанами (ГАГ), которые различаются составом дисахаридов и их
Слайд 7Полисахариды – гликозамингликаны и протеогликаны, а также и адгезивные белки составляют группу
Полисахариды – гликозамингликаны и протеогликаны, а также и адгезивные белки составляют группу

Слайд 8Коллаген – основной структурный белок соединительной ткани
Коллагены составляют приблизительно 30 % общего
Коллаген – основной структурный белок соединительной ткани
Коллагены составляют приблизительно 30 % общего

Слайд 9
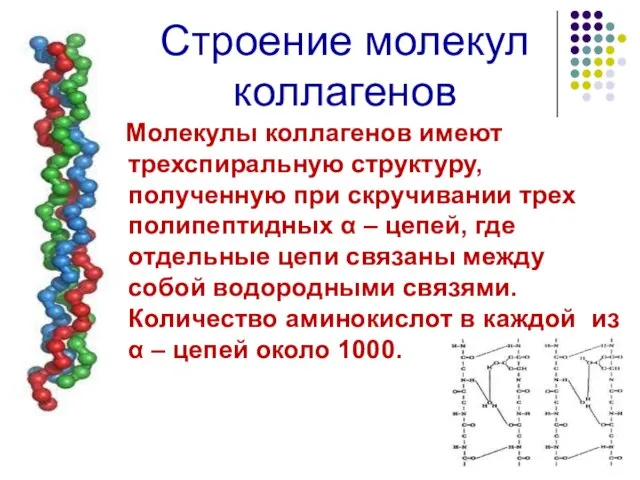
Строение молекул коллагенов
Молекулы коллагенов имеют трехспиральную структуру, полученную при скручивании трех
Строение молекул коллагенов
Молекулы коллагенов имеют трехспиральную структуру, полученную при скручивании трех
Слайд 10Особенности аминокислотного состава коллагена
Полипептидная цепь коллагена состоит из повторяющихся триплетов:
[Гли-Х-Y],
Особенности аминокислотного состава коллагена
Полипептидная цепь коллагена состоит из повторяющихся триплетов:
[Гли-Х-Y],
![Особенности аминокислотного состава коллагена Полипептидная цепь коллагена состоит из повторяющихся триплетов: [Гли-Х-Y],](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/367341/slide-9.jpg)
Слайд 11
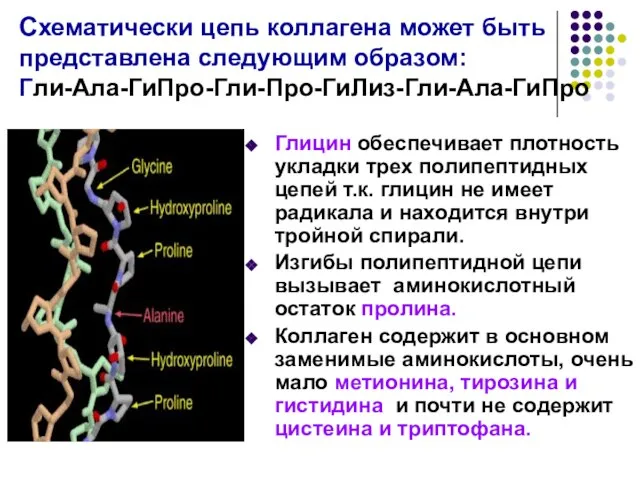
Схематически цепь коллагена может быть представлена следующим образом:
Гли-Ала-ГиПро-Гли-Про-ГиЛиз-Гли-Ала-ГиПро
Глицин обеспечивает плотность укладки
Схематически цепь коллагена может быть представлена следующим образом:
Гли-Ала-ГиПро-Гли-Про-ГиЛиз-Гли-Ала-ГиПро
Глицин обеспечивает плотность укладки
Слайд 12Синтез коллагена
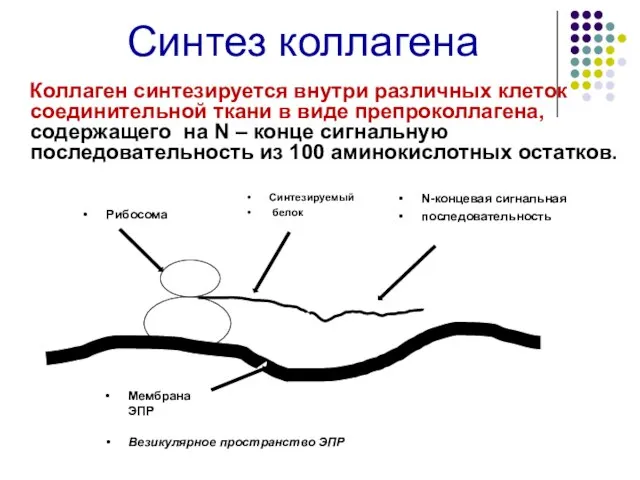
Коллаген синтезируется внутри различных клеток соединительной ткани в виде препроколлагена,
Синтез коллагена
Коллаген синтезируется внутри различных клеток соединительной ткани в виде препроколлагена,
Слайд 13Созревание коллагена (процессинг)
После синтеза цепи коллагена следует сложный многоступенчатый процесс - созревания
Созревание коллагена (процессинг)
После синтеза цепи коллагена следует сложный многоступенчатый процесс - созревания

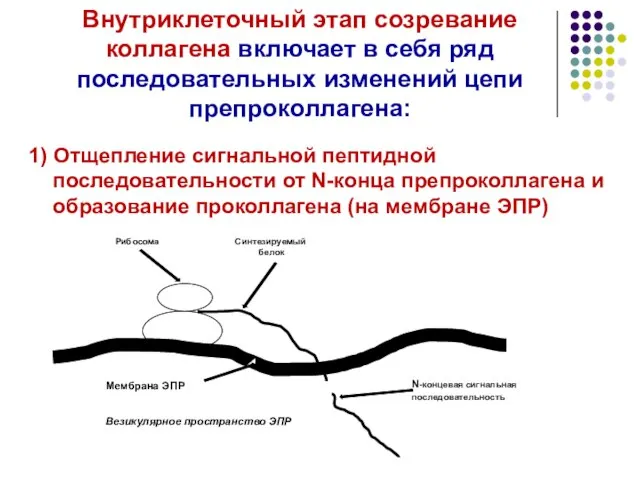
Слайд 14Внутриклеточный этап созревание коллагена включает в себя ряд последовательных изменений цепи препроколлагена:
1)
Внутриклеточный этап созревание коллагена включает в себя ряд последовательных изменений цепи препроколлагена:
1)
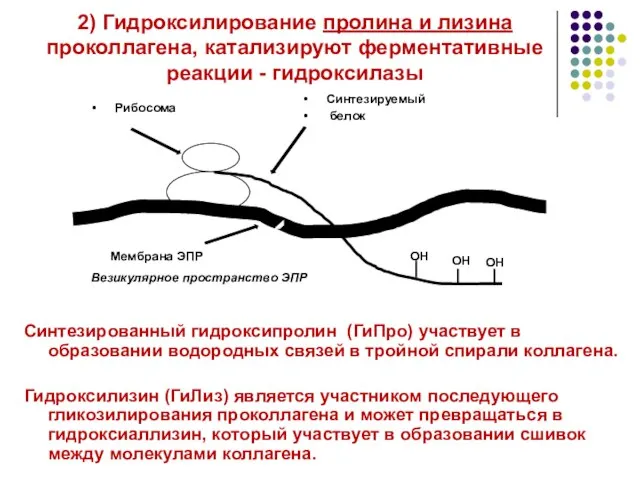
Слайд 152) Гидроксилирование пролина и лизина проколлагена, катализируют ферментативные реакции - гидроксилазы
Синтезированный гидроксипролин
2) Гидроксилирование пролина и лизина проколлагена, катализируют ферментативные реакции - гидроксилазы
Синтезированный гидроксипролин
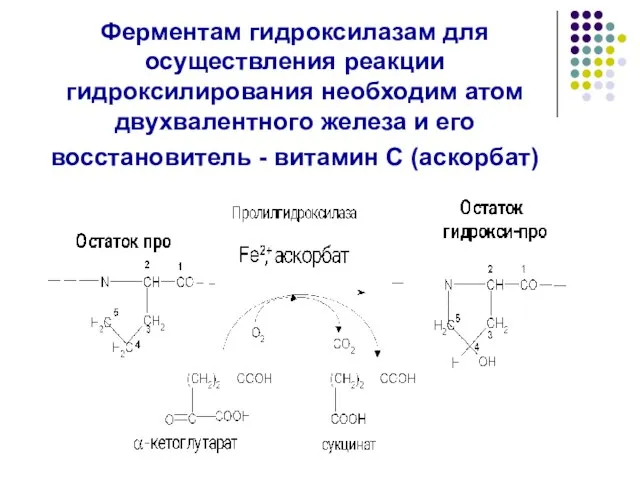
Слайд 16Ферментам гидроксилазам для осуществления реакции гидроксилирования необходим атом двухвалентного железа и его
Ферментам гидроксилазам для осуществления реакции гидроксилирования необходим атом двухвалентного железа и его
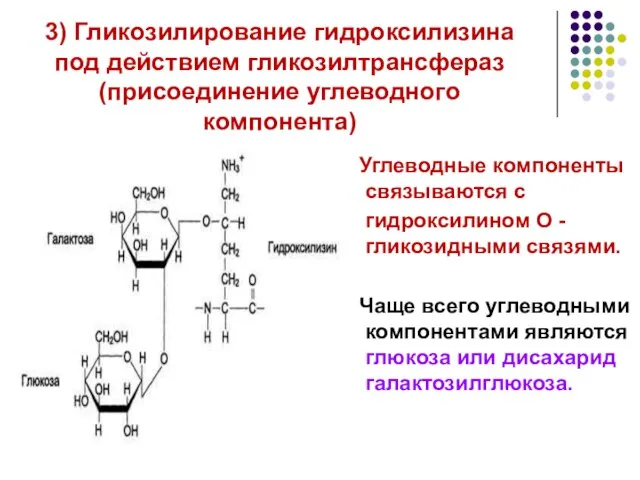
Слайд 173) Гликозилирование гидроксилизина под действием гликозилтрансфераз (присоединение углеводного компонента)
Углеводные компоненты
3) Гликозилирование гидроксилизина под действием гликозилтрансфераз (присоединение углеводного компонента)
Углеводные компоненты
Слайд 184) Формирование тройной спирали проколлагена
Каждая α – цепь проколлагена соединяется водородными связями
4) Формирование тройной спирали проколлагена
Каждая α – цепь проколлагена соединяется водородными связями

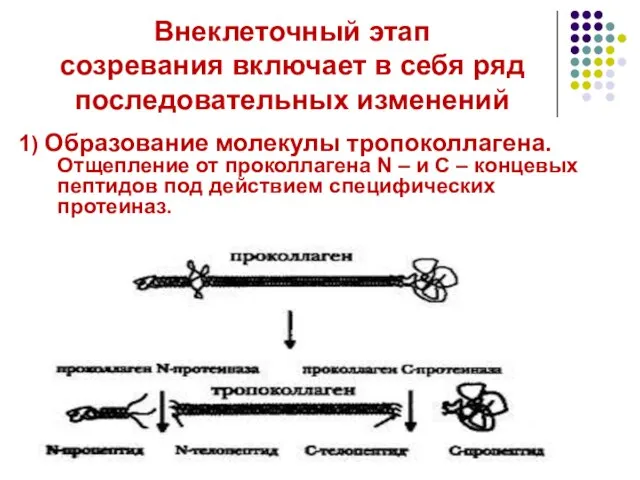
Слайд 19Внеклеточный этап
созревания включает в себя ряд последовательных изменений
1) Образование молекулы тропоколлагена. Отщепление
Внеклеточный этап
созревания включает в себя ряд последовательных изменений
1) Образование молекулы тропоколлагена. Отщепление
Слайд 202) Формирование фибрилл коллагена
Из сформировавшихся молекул тропоколлагена происходит сборка коллагеновых фибрилл,
2) Формирование фибрилл коллагена
Из сформировавшихся молекул тропоколлагена происходит сборка коллагеновых фибрилл,

Слайд 213) Стабилизация и укрепление фибрилл коллагена
Модифицированные аминокислоты - гидроксипролин и гидроксилизин играют
3) Стабилизация и укрепление фибрилл коллагена
Модифицированные аминокислоты - гидроксипролин и гидроксилизин играют

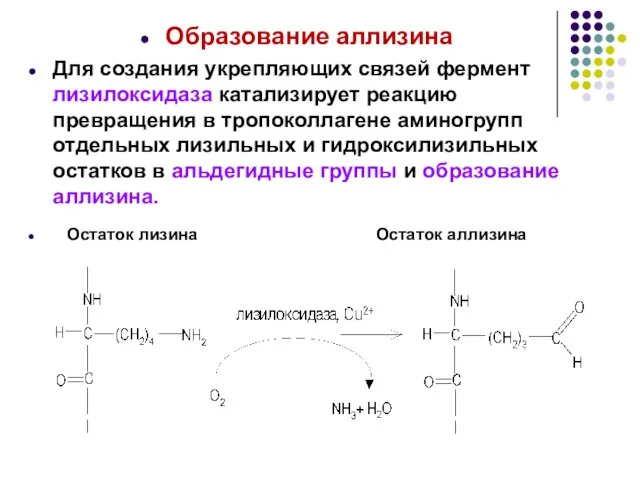
Слайд 22Образование аллизина
Для создания укрепляющих связей фермент лизилоксидаза катализирует реакцию превращения в
Образование аллизина
Для создания укрепляющих связей фермент лизилоксидаза катализирует реакцию превращения в
Слайд 23Возникшие альдегидные группы участвуют в образовании ковалентных связей, которые стабилизируют фибриллы коллагена.
Возникшие альдегидные группы участвуют в образовании ковалентных связей, которые стабилизируют фибриллы коллагена.

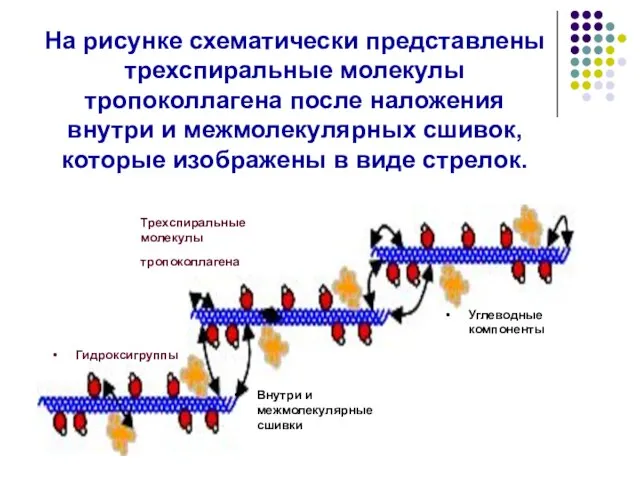
Слайд 25На рисунке схематически представлены трехспиральные молекулы тропоколлагена после наложения внутри и межмолекулярных
На рисунке схематически представлены трехспиральные молекулы тропоколлагена после наложения внутри и межмолекулярных

Слайд 27
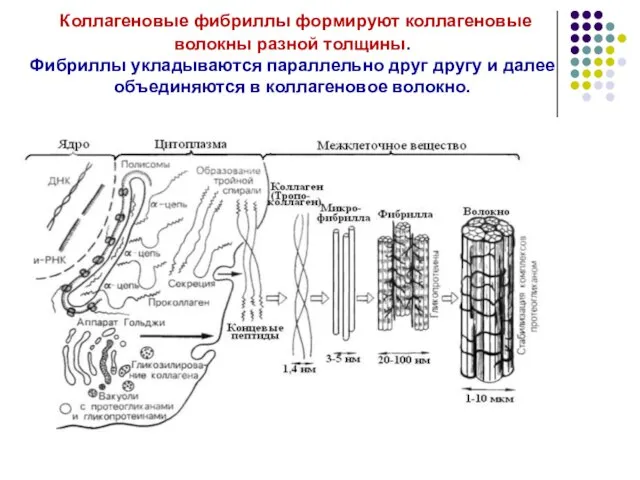
Коллагеновые фибриллы формируют коллагеновые волокны разной толщины.
Фибриллы укладываются параллельно друг
Коллагеновые фибриллы формируют коллагеновые волокны разной толщины. Фибриллы укладываются параллельно друг
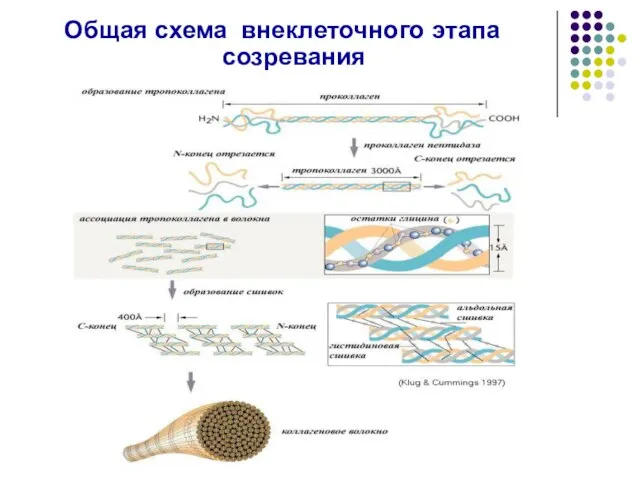
Слайд 28Общая схема внеклеточного этапа созревания
Общая схема внеклеточного этапа созревания
Слайд 29Уникальные свойства коллагенов
Коллагеновые волокна обладают огромной прочностью и практически нерастяжимы. Они могут
Уникальные свойства коллагенов
Коллагеновые волокна обладают огромной прочностью и практически нерастяжимы. Они могут

Слайд 30Катаболизм коллагена
Распад коллагена происходит медленно под действием коллагеназ.
Основной фермент - Са
Катаболизм коллагена
Распад коллагена происходит медленно под действием коллагеназ.
Основной фермент - Са

Слайд 31Основной маркер распада коллагена
Важнейший метаболитом характеризующим скорость распада коллагена является гидроксипролин.
Повышение
Основной маркер распада коллагена
Важнейший метаболитом характеризующим скорость распада коллагена является гидроксипролин.
Повышение
Слайд 32Типы коллагена
В настоящее время известно около 20 различных типов коллагена, различающихся по
Типы коллагена
В настоящее время известно около 20 различных типов коллагена, различающихся по

Слайд 33Наиболее распространенные типы коллагенов.
Наиболее распространенные типы коллагенов.
Слайд 34Эластин – это основной компонент эластических волокон
Содержатся в тканях, обладающих значительной
Эластин – это основной компонент эластических волокон
Содержатся в тканях, обладающих значительной
Слайд 35Особенности аминокислотного состава эластина
Эластин – гликопротеин с молекулярной массой 70кДа, содержит много
Особенности аминокислотного состава эластина
Эластин – гликопротеин с молекулярной массой 70кДа, содержит много

Слайд 36Структура эластина
Нативные волокна эластина построены из молекул, соединенных в тяжи с помощью
Структура эластина
Нативные волокна эластина построены из молекул, соединенных в тяжи с помощью
Слайд 37Неколлагеновые белки
межклеточного матрикса
К этой группе белков относятся относятся адгезивные белки, такие
Неколлагеновые белки
межклеточного матрикса
К этой группе белков относятся относятся адгезивные белки, такие

Слайд 38Фибронектин состоят из 2-х идентичных цепей, содержащих по 7-8 доменов со специфичными
Фибронектин состоят из 2-х идентичных цепей, содержащих по 7-8 доменов со специфичными
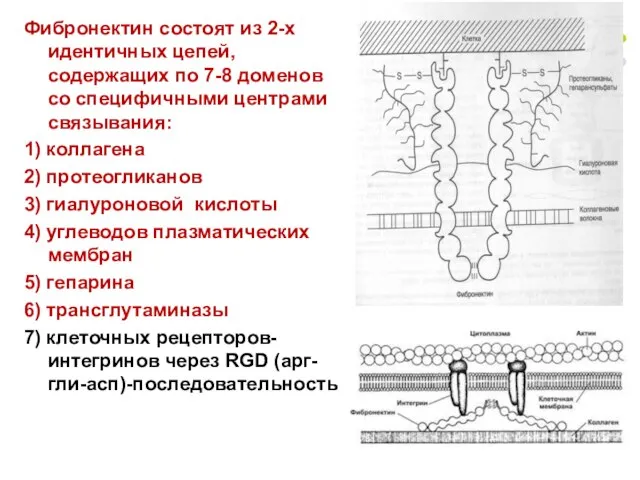
Слайд 39 Адгезивные белки часто содержат последовательность Арг-Гли-Асп (RGD), участвующую в присоединении к клеточным
Адгезивные белки часто содержат последовательность Арг-Гли-Асп (RGD), участвующую в присоединении к клеточным

Слайд 40Ламинины – гликопротеины базальных мембран (3), отделяющую соединительную
ткань от эпителия
Содержит несколько
Ламинины – гликопротеины базальных мембран (3), отделяющую соединительную
ткань от эпителия
Содержит несколько

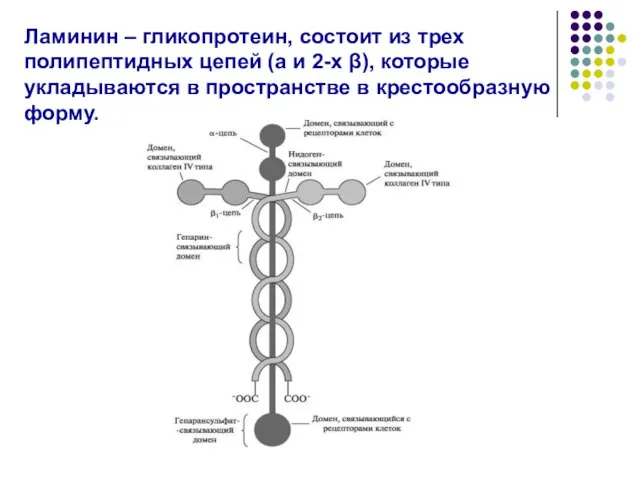
Слайд 41Ламинин – гликопротеин, состоит из трех полипептидных цепей (a и 2-х β),
Ламинин – гликопротеин, состоит из трех полипептидных цепей (a и 2-х β),
Слайд 42К неколлагеновым белкам относятся Gla-белки. (это белки, содержащие аминокислотные остатки 7-карбокси глутаминовой
К неколлагеновым белкам относятся Gla-белки. (это белки, содержащие аминокислотные остатки 7-карбокси глутаминовой

Слайд 43К неколлагеновым белкам
межклеточного матрикса относятся протеогликаны
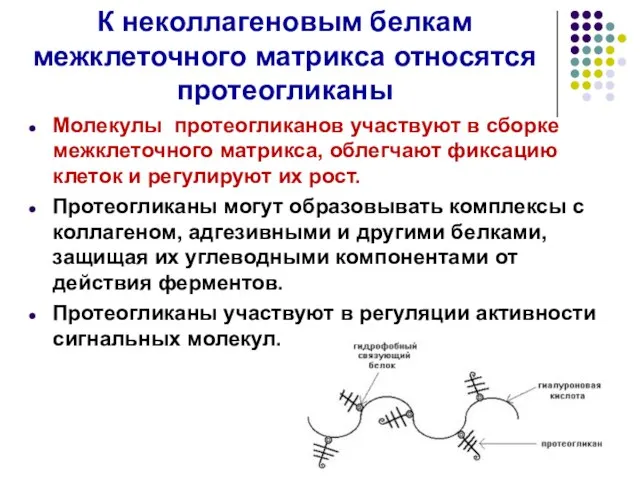
Молекулы протеогликанов участвуют в сборке межклеточного
К неколлагеновым белкам
межклеточного матрикса относятся протеогликаны
Молекулы протеогликанов участвуют в сборке межклеточного
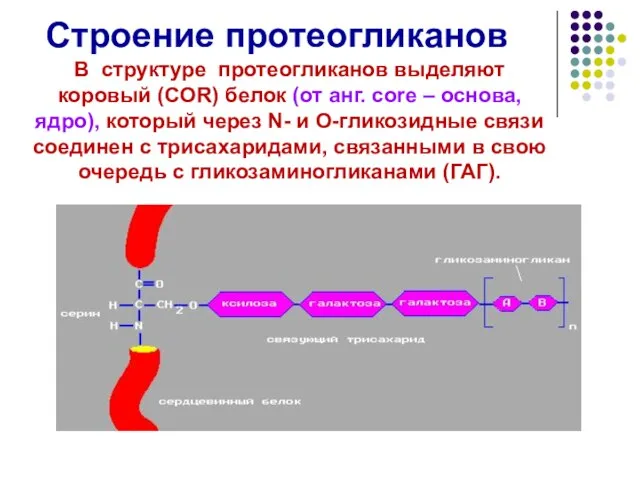
Слайд 44Строение протеогликанов
В структуре протеогликанов выделяют коровый (COR) белок (от анг. сore
Строение протеогликанов
В структуре протеогликанов выделяют коровый (COR) белок (от анг. сore
Слайд 45Основную часть протеогликанов составляют гликозамингликаны (ГАГ)
Гликозаминогликаны – гетерополисахариды, состоящие из повторяющихся дисахаридов,
Основную часть протеогликанов составляют гликозамингликаны (ГАГ)
Гликозаминогликаны – гетерополисахариды, состоящие из повторяющихся дисахаридов,
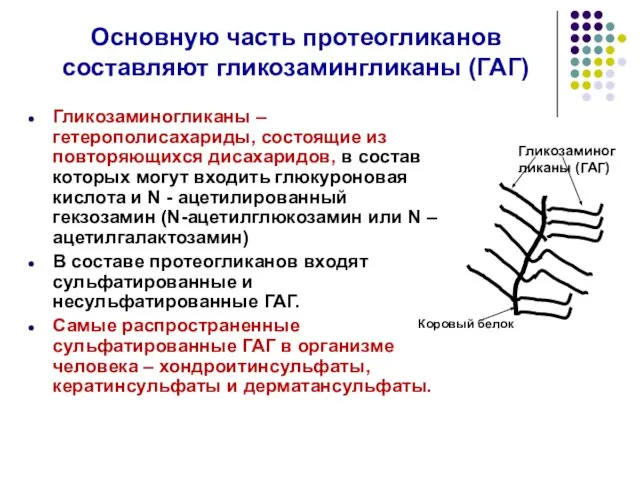
Слайд 46Хондроитинсульфат построен из глюкуроновой кислоты и сульфатированного N–ацетилгалактозамина.
Хондроитинсульфат построен из глюкуроновой кислоты и сульфатированного N–ацетилгалактозамина.
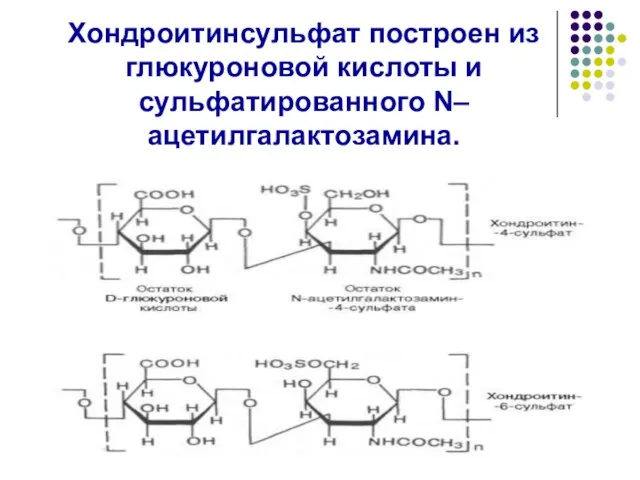
Слайд 47Дисахаридный фрагмент дерматансульфата.
Дисахаридный фрагмент дерматансульфата.
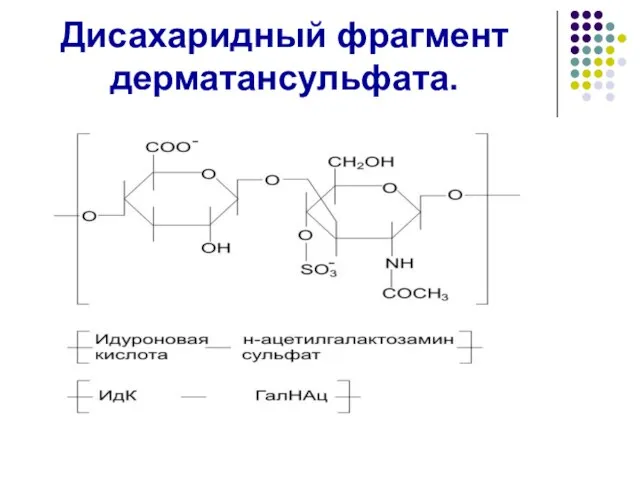
Слайд 48
Дисахариды кератинсульфата состоят из галактозы и сульфатированного N–ацетилглюкозамина.
Дисахариды кератинсульфата состоят из галактозы и сульфатированного N–ацетилглюкозамина.
Слайд 49Гиалуроновая кислота - полимер, состоящий из остатков D-глюкуроновой кислоты и N-ацетилглюкозамина (около
Гиалуроновая кислота - полимер, состоящий из остатков D-глюкуроновой кислоты и N-ацетилглюкозамина (около
Слайд 50Схема агрегатов протеогликанов
Схема агрегатов протеогликанов
Слайд 51Функции гликозаминогликанов
- участвуют в организации межклеточного матрикса, являются основным скрепляющим веществом.
Функции гликозаминогликанов
- участвуют в организации межклеточного матрикса, являются основным скрепляющим веществом.

Слайд 52Метаболизм протеогликанов
Синтез протеогликанов начинается с синтеза корового белка в клетках соединительной ткани,
Метаболизм протеогликанов
Синтез протеогликанов начинается с синтеза корового белка в клетках соединительной ткани,
Слайд 53Организация внеклеточного матрикса в суставном хряще
«Биохимия», ред. Е.С.Северина)
Организация внеклеточного матрикса в суставном хряще
«Биохимия», ред. Е.С.Северина)
 Мой четвероногий друг – собака
Мой четвероногий друг – собака Великая Отечественная Война
Великая Отечественная Война Искусство Москвы. Памятники Соборной площади
Искусство Москвы. Памятники Соборной площади Презентация на тему Давление газов. Закон Паскаля
Презентация на тему Давление газов. Закон Паскаля Барьеры общения
Барьеры общения Классификация парламентов в современный период Мосина
Классификация парламентов в современный период Мосина Героизм советского человека в годы Великой Отечественной войны
Героизм советского человека в годы Великой Отечественной войны Business excellence
Business excellence Традиции семьи для квеста
Традиции семьи для квеста Игра-викторина С Рождеством Христовым!
Игра-викторина С Рождеством Христовым! Санги Стиль
Санги Стиль «Порядок подготовки отчета о проведении самооценки образовательных программ»
«Порядок подготовки отчета о проведении самооценки образовательных программ» Выделение хитина и хитозана из оболочек цист Artemia salina под воздействием МВИ
Выделение хитина и хитозана из оболочек цист Artemia salina под воздействием МВИ PRESENT PERFECT
PRESENT PERFECT  Презентация на тему История развития вычислительной техники
Презентация на тему История развития вычислительной техники Presentation
Presentation Система высшего образования в Японии Подготовили студентки 2 курса ФТД Группа Тс01/1301 Анохина Татьяна и Шубина Арина
Система высшего образования в Японии Подготовили студентки 2 курса ФТД Группа Тс01/1301 Анохина Татьяна и Шубина Арина ВОЛОКОННЫЕ ЛАЗЕРЫ
ВОЛОКОННЫЕ ЛАЗЕРЫ  Формирование личного бренда графического дизайнера
Формирование личного бренда графического дизайнера Информационно- коммуникационные технологии в школе
Информационно- коммуникационные технологии в школе Трагедия в Герасимовке (о Павлике Морозове)
Трагедия в Герасимовке (о Павлике Морозове) Методы и приемы восприятия неречевых звуков детьми с нарушением слуха
Методы и приемы восприятия неречевых звуков детьми с нарушением слуха Гербы городов мира
Гербы городов мира Самовольное строительство. Практические вопросы применения земельного и градостроительного законодательства
Самовольное строительство. Практические вопросы применения земельного и градостроительного законодательства Терминология швейных работ
Терминология швейных работ Основные требование к транспортным и погрузочным средствам
Основные требование к транспортным и погрузочным средствам Шляпочные грибы 1 класс
Шляпочные грибы 1 класс Здоровая пища для всей семьи
Здоровая пища для всей семьи