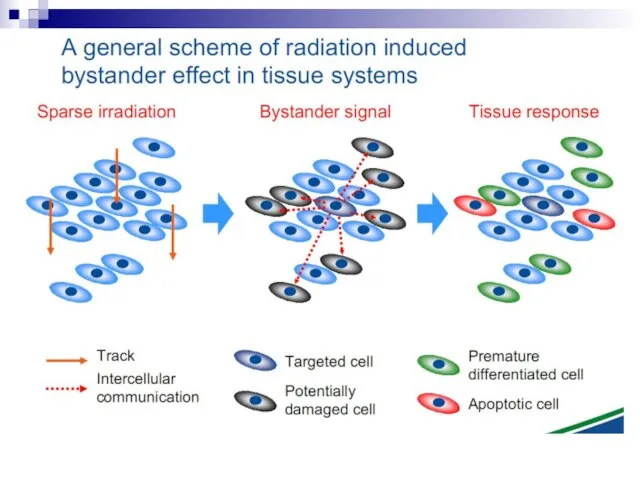
- BYSTANDER EFFECT

Содержание
- 2. The bystander effect refers to the induction of biological effects in cells that are not directly
- 3. Non-targeted biological effects of ionizing radiation
- 5. Non-targeted effects of ionizing radiation as a new paradigm of radiation biology Ward, J. (1999) New
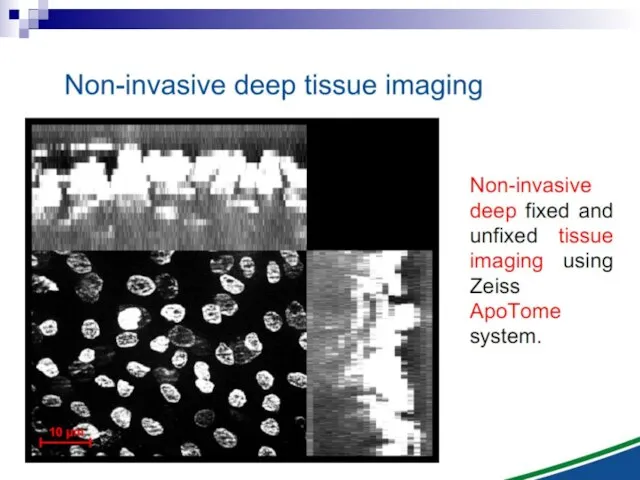
- 12. Bystander effect and genomic instability
- 73. Experiments suggest that the effect is due to a molecule secreted by irradiated cells, which is
- 74. The radiation-induced bystander effect is defined as “the induction of biological effects in cells that are
- 75. GENERATIONS OF students in radiation biology have been taught that heritable biological effects require direct damage
- 76. In the radiation field, it has come to be loosely defined as the induction of biological
- 77. The plethora of data now available concerning the bystander effect fall into two quite separate categories,
- 78. Medium transfer experiments Experiments involving the transfer of medium from irradiated to unirradiated cells have demonstrated
- 79. This bystander effect suggested that irradiated cells secreted a molecule into the culture medium that was
- 80. Some limited progress has been made in the search for the mechanisms involved in this bystander
- 81. The majority of bystander experiments involving medium transfer have utilized low-LET x or gamma rays, in
- 82. Micronuclei in normal human fibroblasts Perhaps the most direct and most dramatic demonstration of the bystander
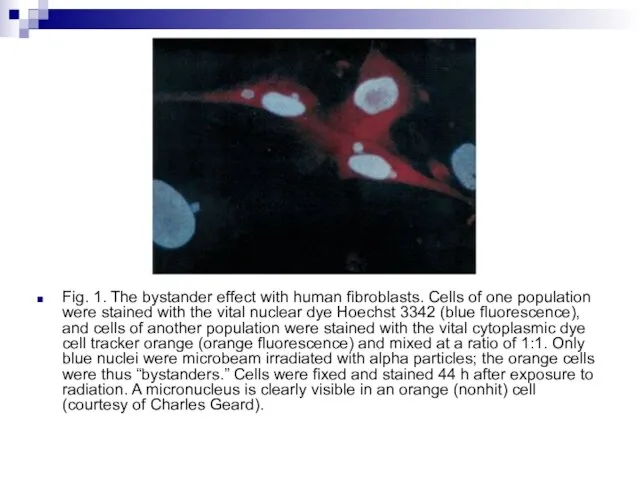
- 83. Fig. 1. The bystander effect with human fibroblasts. Cells of one population were stained with the
- 84. Cell lethality Lines of hygromycin- and neomycin-resistant V79 cells were produced. Before exposure the hygromycinresistant cells
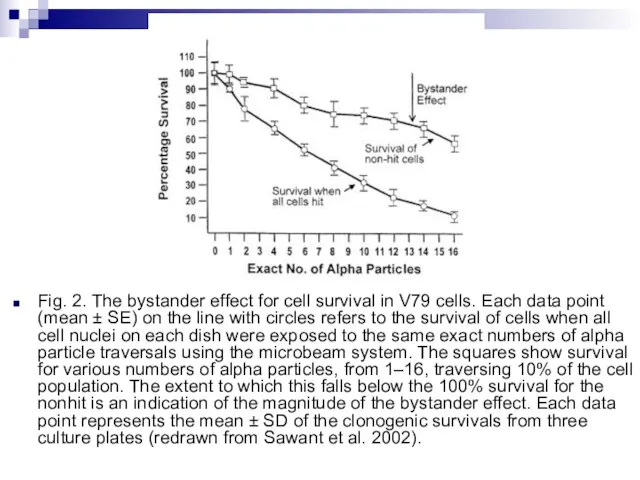
- 85. Fig. 2. The bystander effect for cell survival in V79 cells. Each data point (mean ±
- 86. There is a considerable degree of cell killing in the nonhit cells, implying a substantial bystander
- 87. Mutagenic effects in human-hamster hybrid cells Zhou et al. (2000) reported a study in which human-hamster
- 88. These mutations must clearly arise from neighbor cells, not directly exposed, but in close proximity to
- 89. Oncogenic transformation in mouse fibroblasts Mouse fibroblast (C3H10T1⁄2) cells were plated in a monolayer, and the
- 90. It is important to note that the experimental results discussed in this paper involve laboratory model
- 91. This is illustrated in Fig. 6 which combines the data of Zhou et al. (2001), in
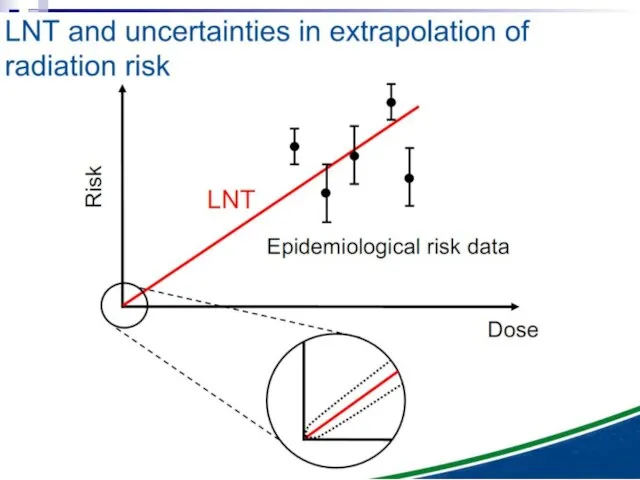
- 92. Under these experimental conditions, it is evident that a linear extrapolation of risks from high doses
- 93. Ionizing radiation Radiated cell BE factor migration (signal) Effects in non - radiated cells BYSTANDER EFFECT
- 94. Scheme of BE induced in vivo (MN test) 6000 cells seeded in 24 cm2 flasks Blood
- 96. Killing Non-transduced Tumor Cells via Bystander Effect The bystander effect was first reported by Moolten (1986)
- 97. Schematic overview of the Bystander effect
- 98. Our understanding of how radiation kills normal and tumour cells has been based on an intimate
- 99. When ionizing radiation interacts with biological material, energy is deposited and chemical bonds are broken. In
- 100. Direct DNA damage radiation model. The schematic shows the standard model of DNA damage responses to
- 101. The mechanisms underpinning DNA damage and repair processing in irradiated cells have been extensively studied since
- 102. Evidence now shows that, as well as these direct DNA damage-dependent effects, irradiated cells also send
- 103. For example, the archetypal gene therapy model is the herpes simplex virus-thymidine kinase (HSV-TK) system. In
- 104. Radiation-induced bystander responses have been observed in a range of cell types, tissue models and in
- 105. Targeted Effects of Ionizing Radiation Cell Damage Induction of death repaired clonal genetic alteration
- 106. Untargeted Effects of Exposure to Ionizing Radiation Effects in unexposed cells and their progeny i.e. in
- 107. Radiation-induced Genomic Instability micronucleus chromosome aberration cell death gene mutation mitotic failure aneuploidy
- 108. Radiation-induced Genomic Instability A genome-wide process induced at very high frequency High LET tends to be
- 109. Bystander Effects of Ionizing Radiation Signals via gap junctions Signals via medium/plasma N.B. 1950s and 60s
- 110. Bystander Effects of Ionizing Radiation Increases in damage-inducible proteins Decreases in damage-inducible proteins Increases in reactive
- 111. Bystander Effects of Ionizing Radiation Target for biological effects is larger than the cell Important implications
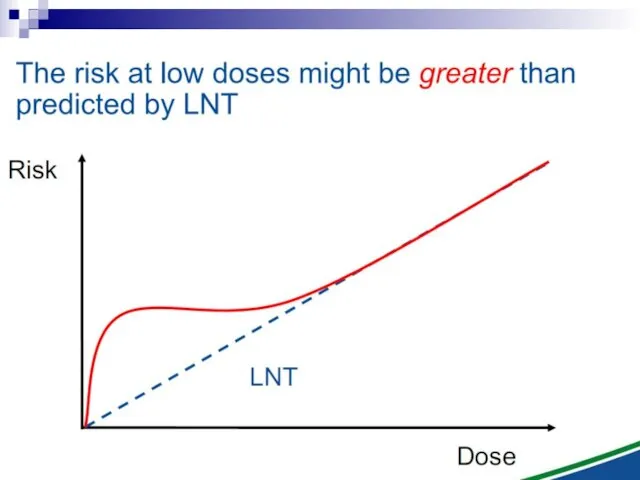
- 112. The Linear No Threshold Problem Induced Effect Dose Threshold supralinear Supralinear linear Threshold sublinear Hormesis Non-targeted
- 113. DNA Damage Genome stability
- 115. Скачать презентацию

Слайд 3Non-targeted biological effects of
ionizing radiation
Non-targeted biological effects of
ionizing radiation
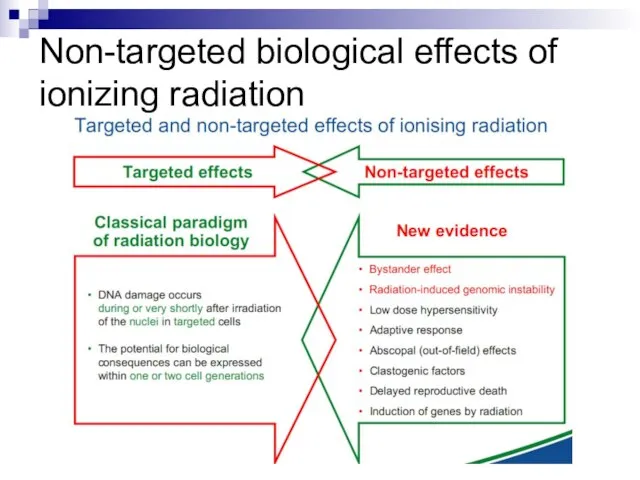

Слайд 5Non-targeted effects
of ionizing radiation
as a new paradigm
of radiation biology
Ward,
Non-targeted effects
of ionizing radiation
as a new paradigm
of radiation biology
Ward,

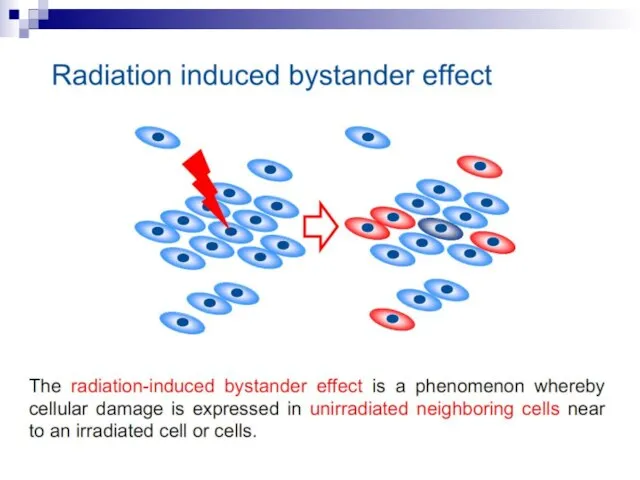
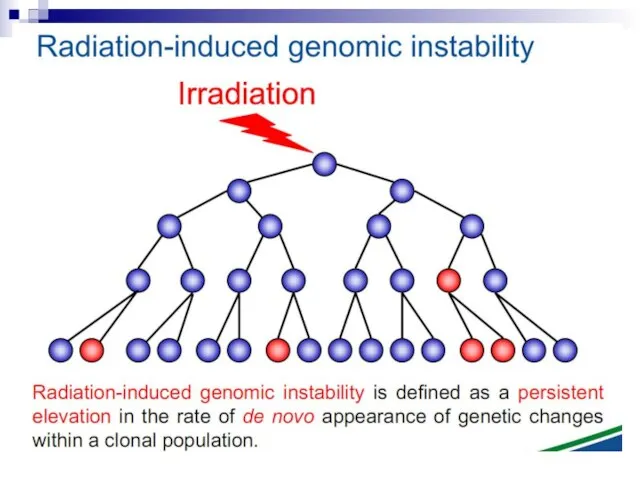


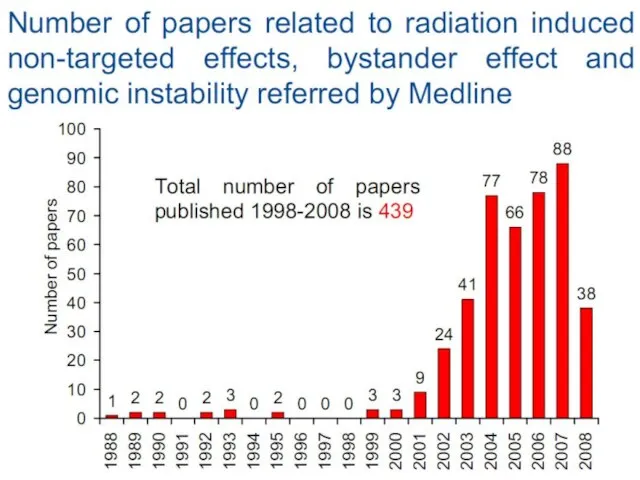

Слайд 12Bystander effect
and genomic instability
Bystander effect
and genomic instability

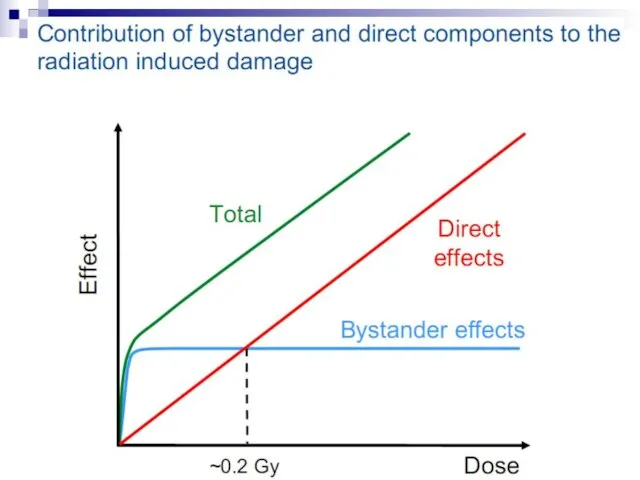
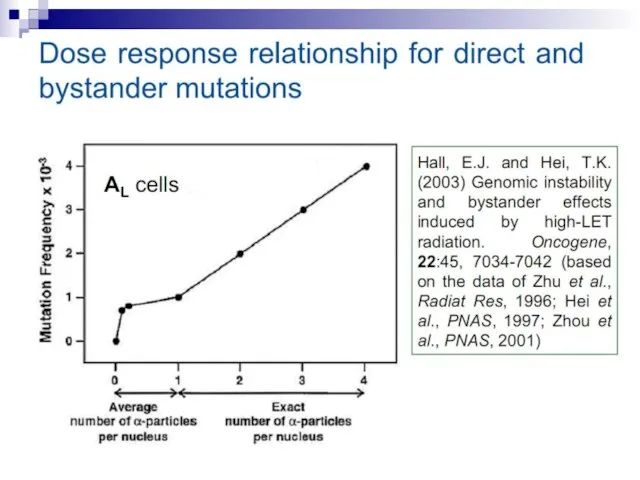

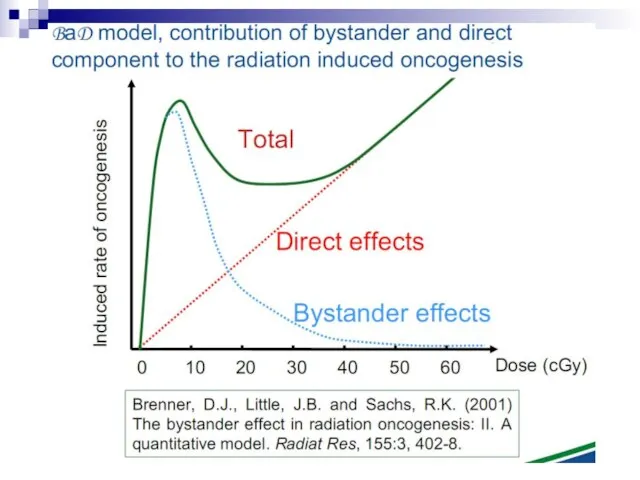
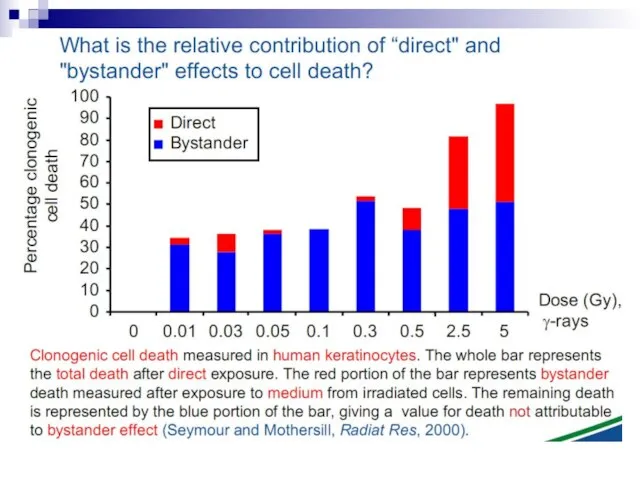

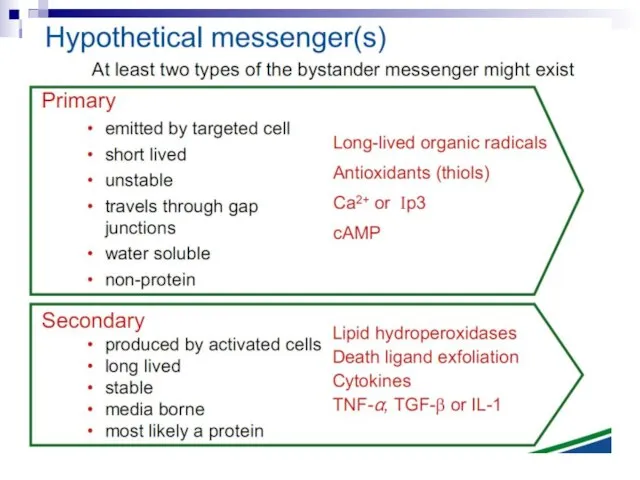



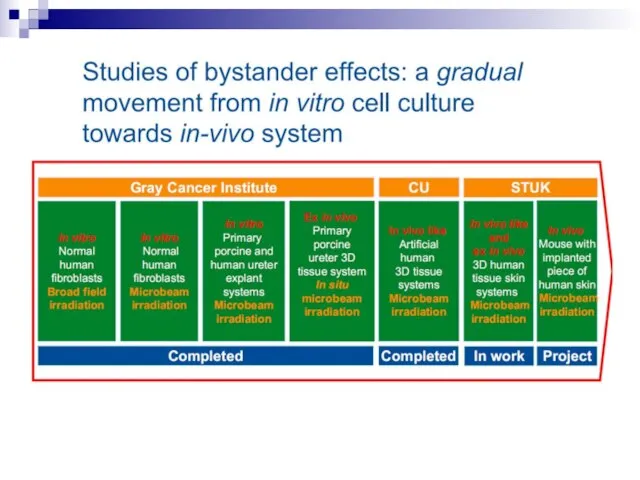



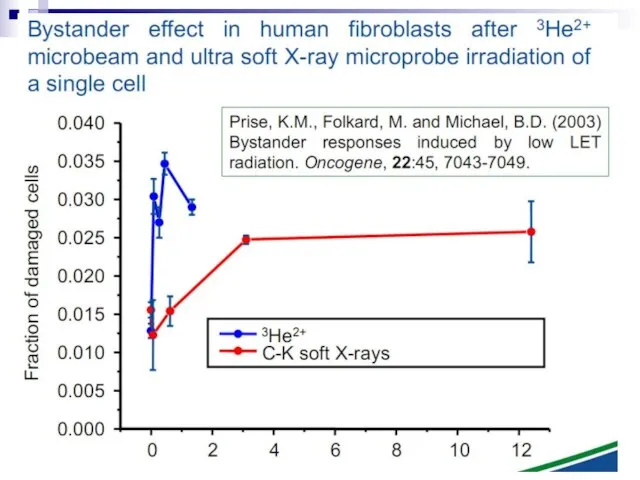
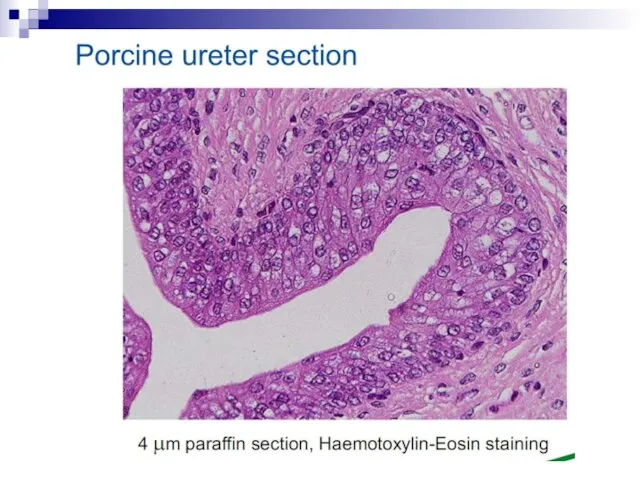
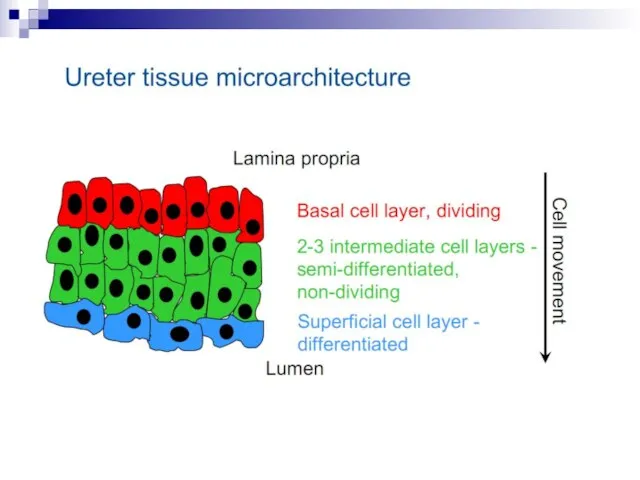
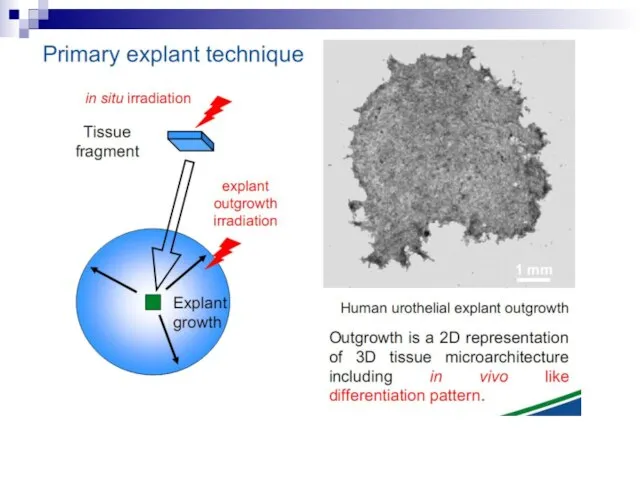

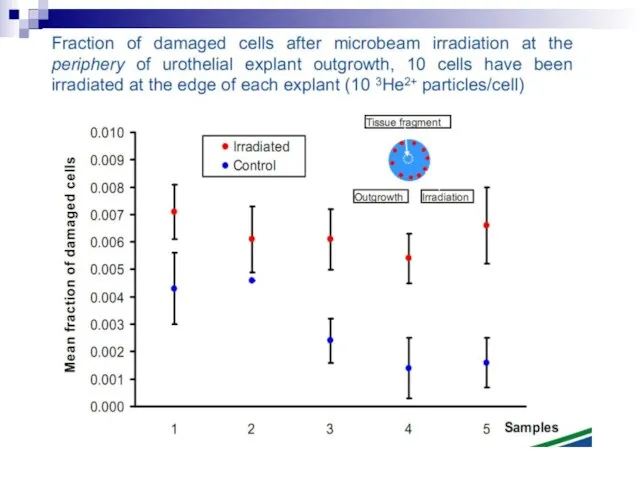

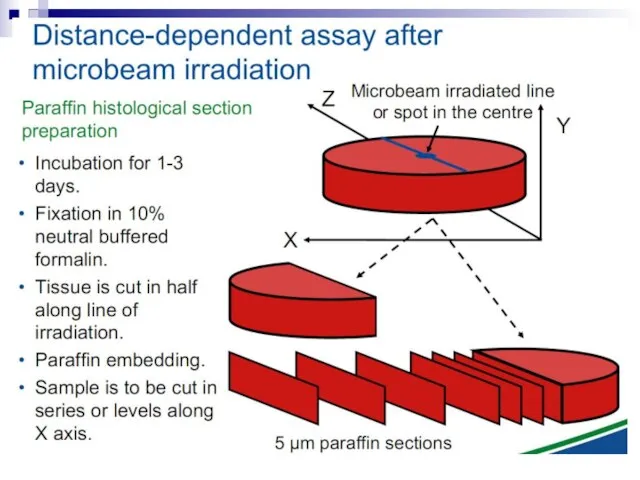
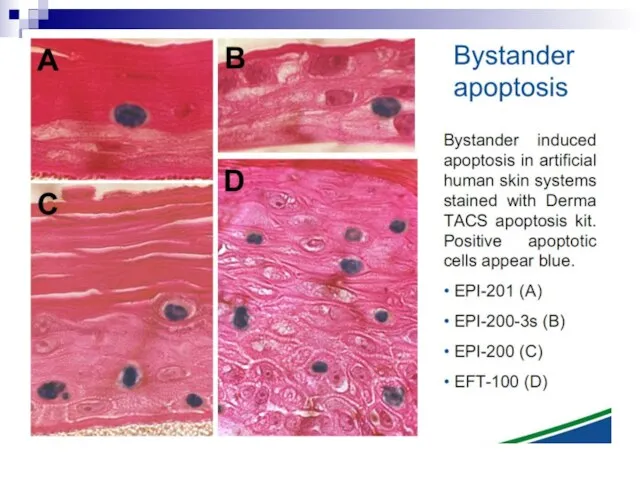

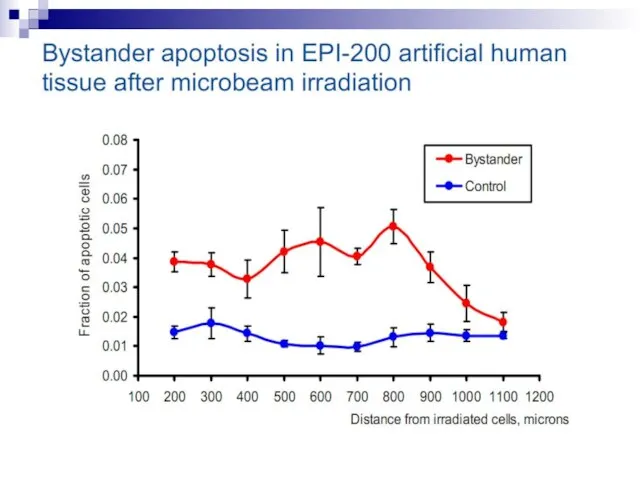
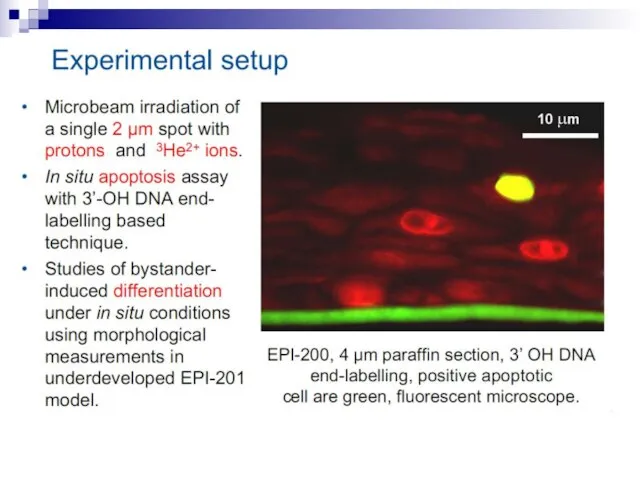
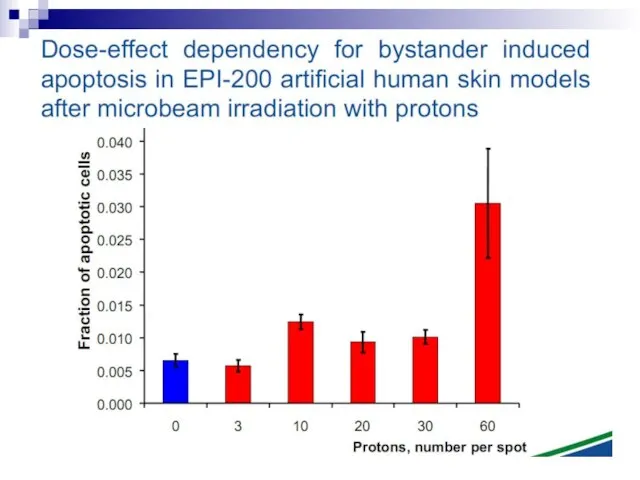
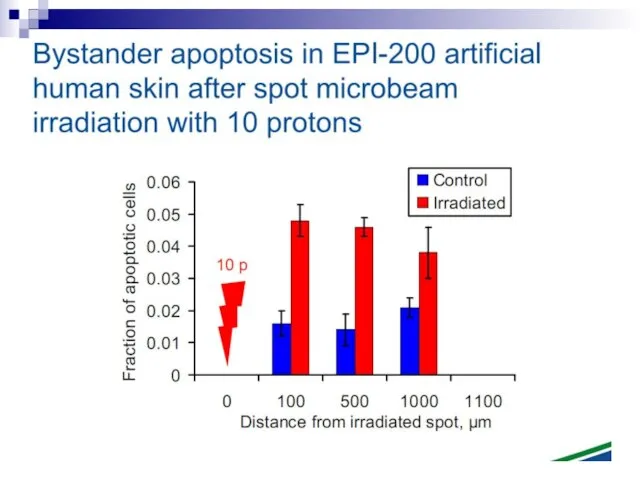
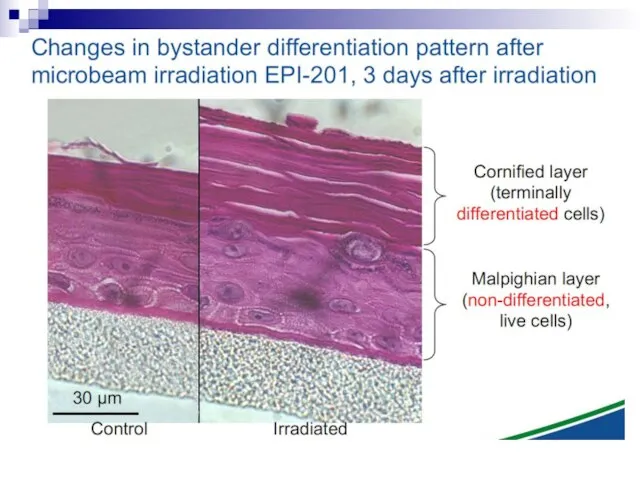
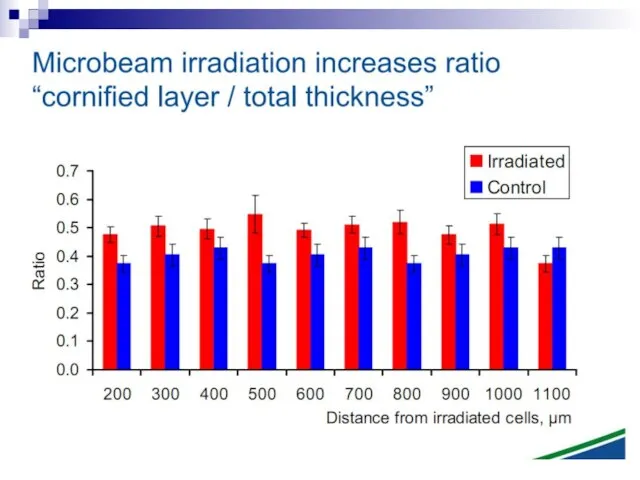
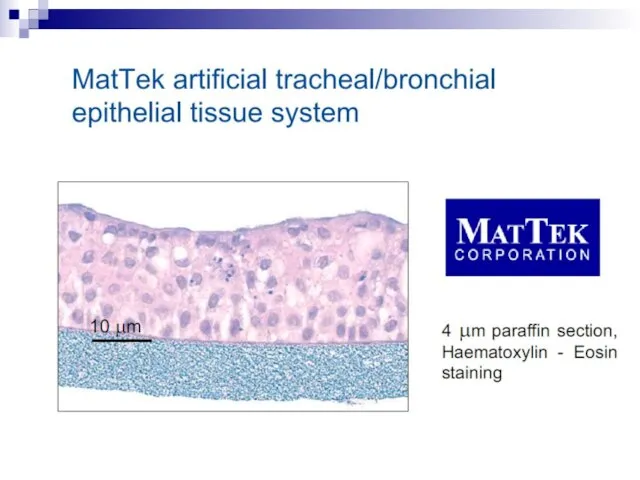
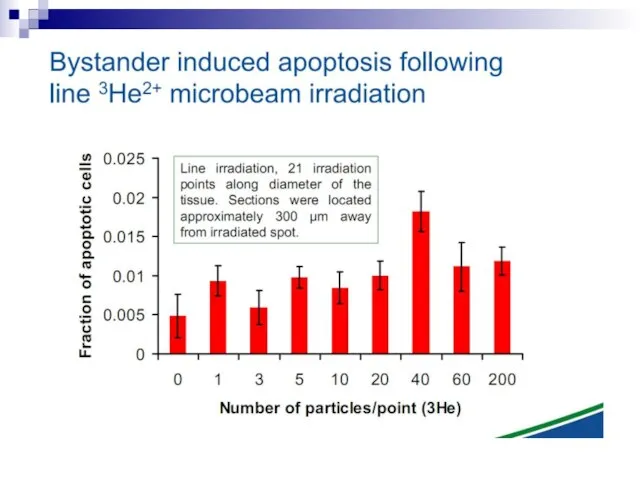
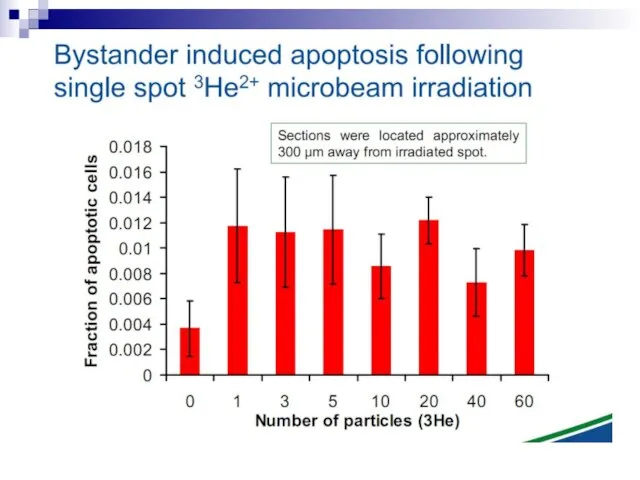
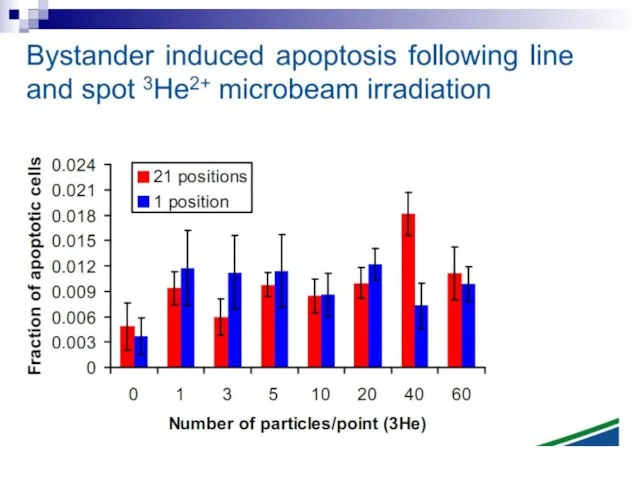















Слайд 73Experiments suggest that the effect is due to a molecule secreted by
Experiments suggest that the effect is due to a molecule secreted by

Слайд 74The radiation-induced bystander effect is defined as “the induction of biological effects
The radiation-induced bystander effect is defined as “the induction of biological effects

Слайд 75GENERATIONS OF students in radiation biology have been taught that heritable biological
GENERATIONS OF students in radiation biology have been taught that heritable biological

Слайд 76In the radiation field, it has come to be loosely defined as
In the radiation field, it has come to be loosely defined as

Слайд 77The plethora of data now available concerning the bystander effect fall into
The plethora of data now available concerning the bystander effect fall into

Слайд 78Medium transfer experiments
Experiments involving the transfer of medium from irradiated to unirradiated
Medium transfer experiments
Experiments involving the transfer of medium from irradiated to unirradiated

Слайд 79This bystander effect suggested that irradiated cells secreted a molecule into the
This bystander effect suggested that irradiated cells secreted a molecule into the

Слайд 80Some limited progress has been made in the search for the mechanisms
Some limited progress has been made in the search for the mechanisms

Слайд 81The majority of bystander experiments involving medium transfer have utilized low-LET x
The majority of bystander experiments involving medium transfer have utilized low-LET x

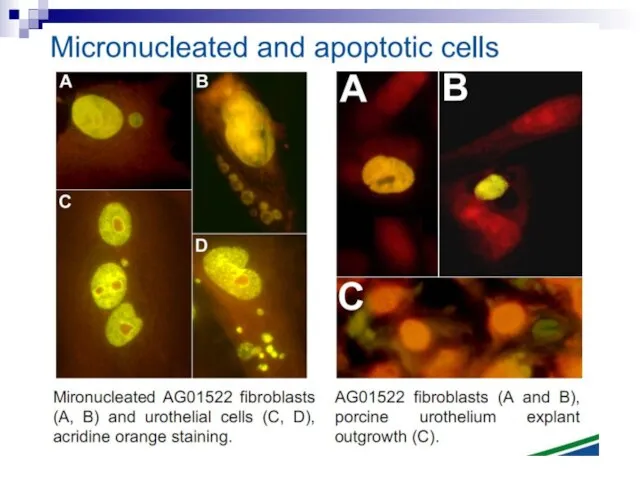
Слайд 82Micronuclei in normal human fibroblasts Perhaps the most direct and most dramatic
Micronuclei in normal human fibroblasts Perhaps the most direct and most dramatic

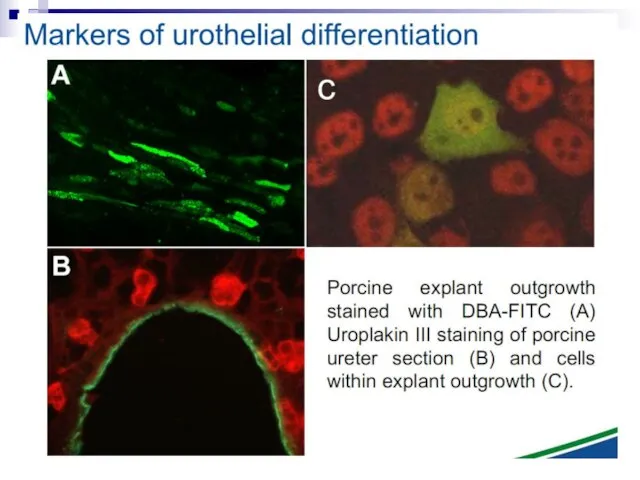
Слайд 83Fig. 1. The bystander effect with human fibroblasts. Cells of one population
Fig. 1. The bystander effect with human fibroblasts. Cells of one population
Слайд 84Cell lethality Lines of hygromycin- and neomycin-resistant V79 cells were produced. Before
Cell lethality Lines of hygromycin- and neomycin-resistant V79 cells were produced. Before

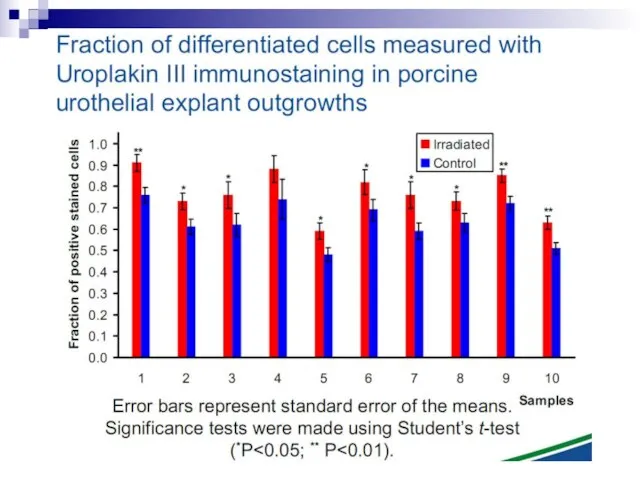
Слайд 85Fig. 2. The bystander effect for cell survival in V79 cells. Each
Fig. 2. The bystander effect for cell survival in V79 cells. Each
Слайд 86There is a considerable degree of cell killing in the nonhit cells,
There is a considerable degree of cell killing in the nonhit cells,

Слайд 87Mutagenic effects in human-hamster hybrid cells Zhou et al. (2000) reported a
Mutagenic effects in human-hamster hybrid cells Zhou et al. (2000) reported a
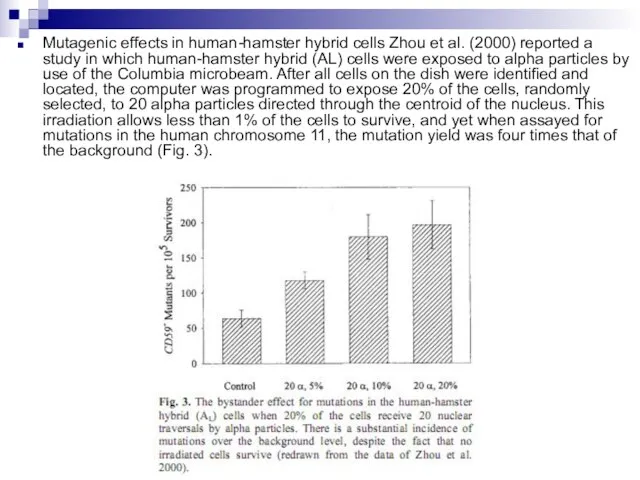
Слайд 88These mutations must clearly arise from neighbor cells, not directly exposed, but
These mutations must clearly arise from neighbor cells, not directly exposed, but
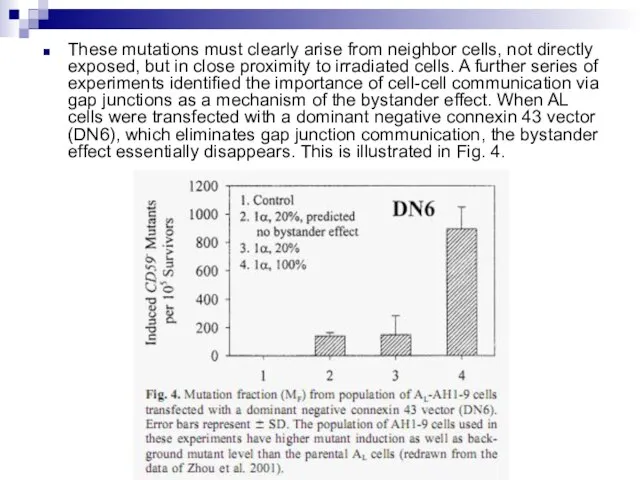
Слайд 89Oncogenic transformation in mouse fibroblasts Mouse fibroblast (C3H10T1⁄2) cells were plated in
Oncogenic transformation in mouse fibroblasts Mouse fibroblast (C3H10T1⁄2) cells were plated in
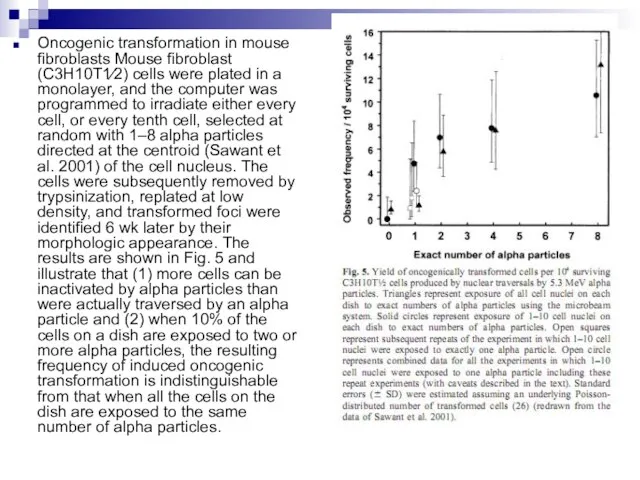
Слайд 90It is important to note that the experimental results discussed in this
It is important to note that the experimental results discussed in this

Слайд 91This is illustrated in Fig. 6 which combines the data of Zhou
This is illustrated in Fig. 6 which combines the data of Zhou
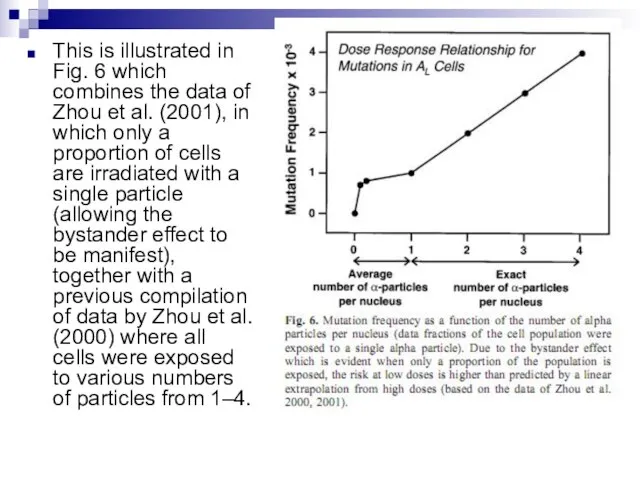
Слайд 92Under these experimental conditions, it is evident that a linear extrapolation of
Under these experimental conditions, it is evident that a linear extrapolation of

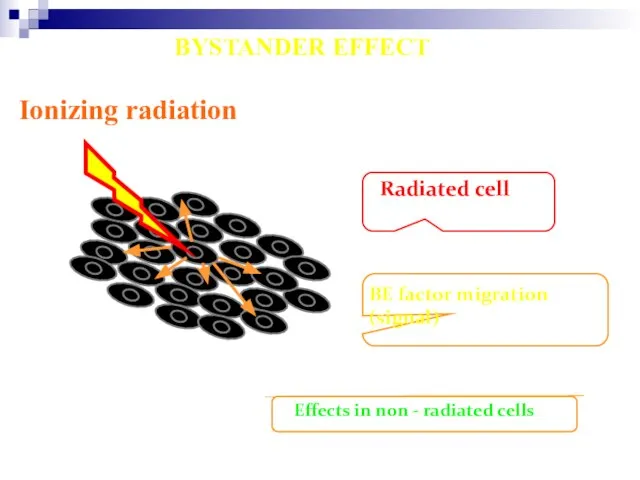
Слайд 93Ionizing radiation
Radiated cell
BE factor migration (signal)
Effects in non - radiated cells
BYSTANDER EFFECT
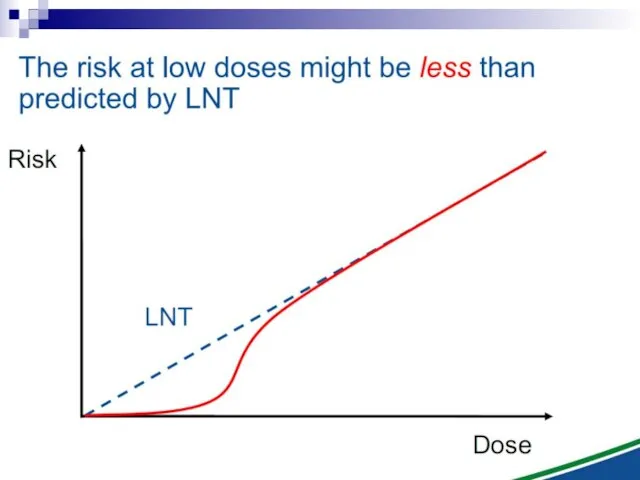
Ionizing radiation
Radiated cell
BE factor migration (signal)
Effects in non - radiated cells
BYSTANDER EFFECT
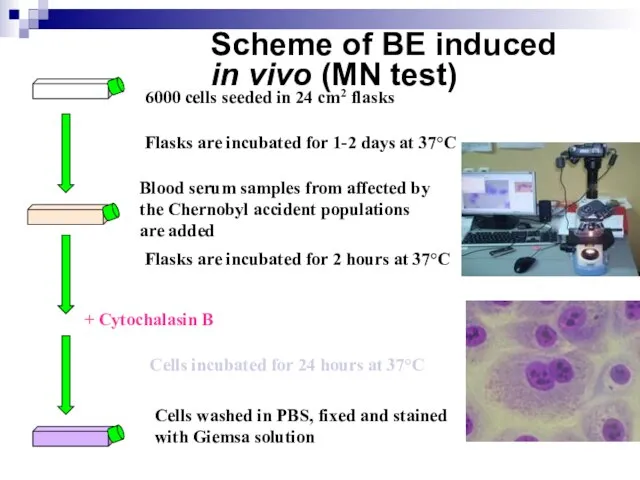
Слайд 94Scheme of BE induced
in vivo (MN test)
6000 cells seeded in 24
Scheme of BE induced
in vivo (MN test)
6000 cells seeded in 24
Слайд 96Killing Non-transduced Tumor Cells via Bystander Effect
The bystander effect was first reported
Killing Non-transduced Tumor Cells via Bystander Effect
The bystander effect was first reported

Слайд 97Schematic overview of the Bystander effect
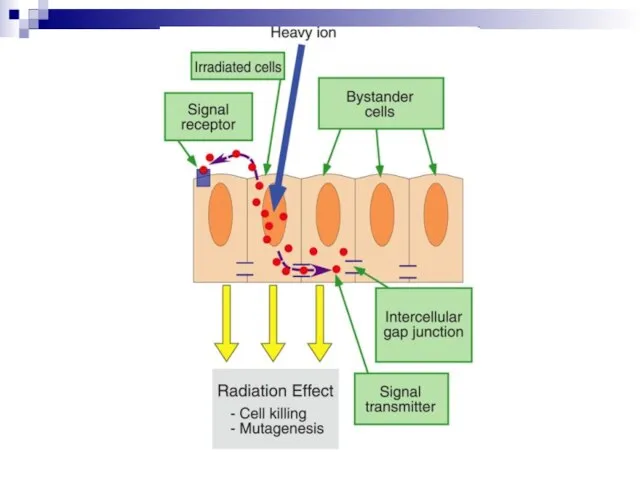
Schematic overview of the Bystander effect

Слайд 98Our understanding of how radiation kills normal and tumour cells has been
Our understanding of how radiation kills normal and tumour cells has been

Слайд 99When ionizing radiation interacts with biological material, energy is deposited and chemical
When ionizing radiation interacts with biological material, energy is deposited and chemical

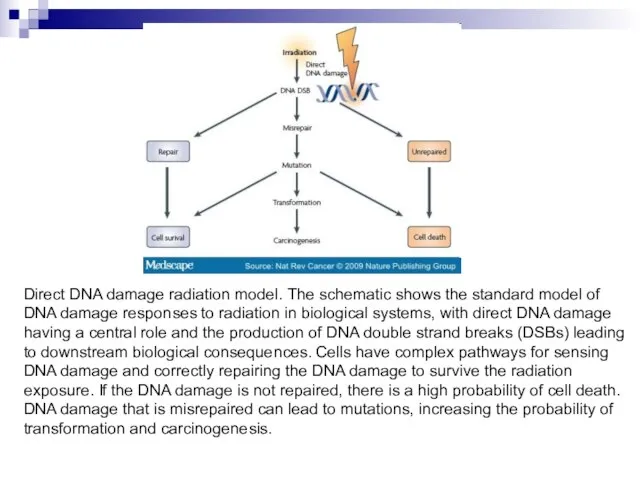
Слайд 100Direct DNA damage radiation model. The schematic shows the standard model of
Direct DNA damage radiation model. The schematic shows the standard model of
Слайд 101The mechanisms underpinning DNA damage and repair processing in irradiated cells have
The mechanisms underpinning DNA damage and repair processing in irradiated cells have

Слайд 102Evidence now shows that, as well as these direct DNA damage-dependent effects,
Evidence now shows that, as well as these direct DNA damage-dependent effects,

Слайд 103For example, the archetypal gene therapy model is the herpes simplex virus-thymidine
For example, the archetypal gene therapy model is the herpes simplex virus-thymidine

Слайд 104Radiation-induced bystander responses have been observed in a range of cell types,
Radiation-induced bystander responses have been observed in a range of cell types,

Слайд 105Targeted Effects of Ionizing Radiation
Cell Damage Induction of death repaired clonal
Targeted Effects of Ionizing Radiation
Cell Damage Induction of death repaired clonal

Слайд 106Untargeted Effects of Exposure
to Ionizing Radiation
Effects in unexposed cells and their
Untargeted Effects of Exposure
to Ionizing Radiation
Effects in unexposed cells and their

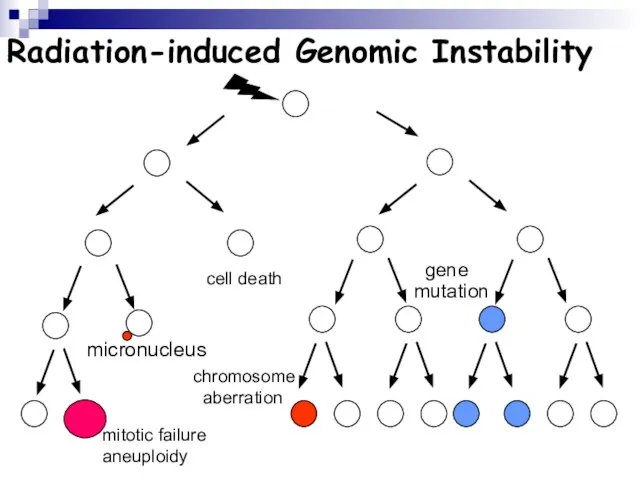
Слайд 107Radiation-induced Genomic Instability
micronucleus
chromosome
aberration
cell death
gene
mutation
mitotic failure
aneuploidy
Radiation-induced Genomic Instability
micronucleus
chromosome
aberration
cell death
gene
mutation
mitotic failure
aneuploidy
Слайд 108Radiation-induced Genomic Instability
A genome-wide process induced at very high frequency
High
Radiation-induced Genomic Instability
A genome-wide process induced at very high frequency
High

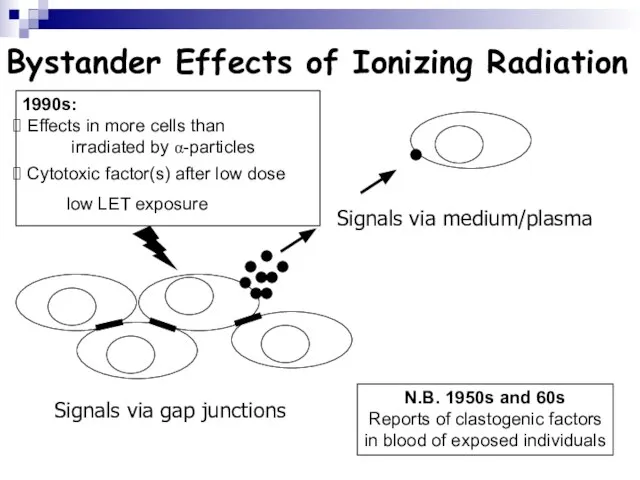
Слайд 109Bystander Effects of Ionizing Radiation
Signals via gap junctions
Signals via medium/plasma
N.B. 1950s and
Bystander Effects of Ionizing Radiation
Signals via gap junctions
Signals via medium/plasma
N.B. 1950s and
Слайд 110Bystander Effects of Ionizing Radiation
Increases in damage-inducible proteins
Decreases in damage-inducible
Bystander Effects of Ionizing Radiation
Increases in damage-inducible proteins
Decreases in damage-inducible

Слайд 111Bystander Effects of Ionizing Radiation
Target for biological effects is larger than
Bystander Effects of Ionizing Radiation
Target for biological effects is larger than

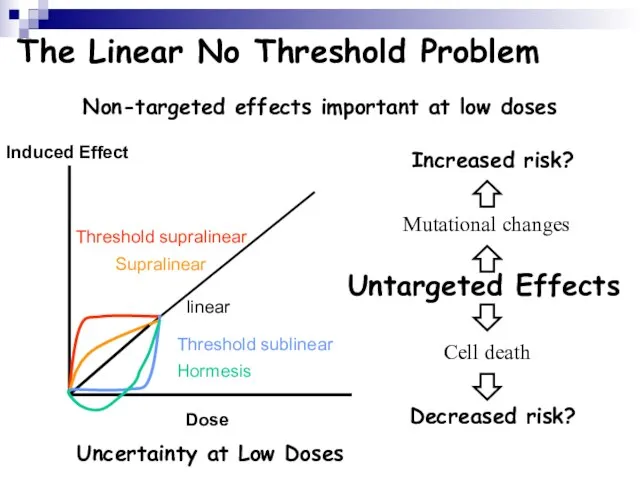
Слайд 112The Linear No Threshold Problem
Induced Effect
Dose
Threshold supralinear
Supralinear
linear
Threshold sublinear
Hormesis
Non-targeted effects important at low
The Linear No Threshold Problem
Induced Effect
Dose
Threshold supralinear
Supralinear
linear
Threshold sublinear
Hormesis
Non-targeted effects important at low
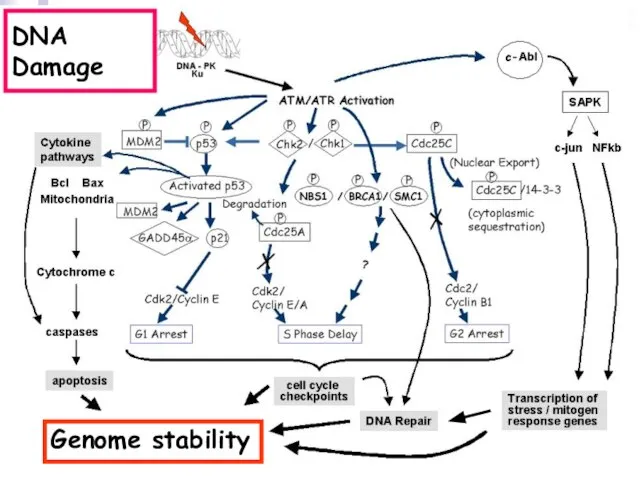
Слайд 113DNA Damage
Genome stability
DNA Damage
Genome stability
 Типология современного урока
Типология современного урока Растительный и животный мир
Растительный и животный мир Начни здоровья марафон - приходи в полиатлон
Начни здоровья марафон - приходи в полиатлон Общество евангелизации детей
Общество евангелизации детей По ту сторону света
По ту сторону света Биологические методы борьбы с вредителями
Биологические методы борьбы с вредителями Презентация на тему Культура: понятие, многообразие, формы
Презентация на тему Культура: понятие, многообразие, формы Лекарственные препараты 10 класс
Лекарственные препараты 10 класс Долина гейзеров на полуострове Камчатка
Долина гейзеров на полуострове Камчатка Схемотехника аналоговых электронных устройств. Практика 8. Операционные усилители и устройства на их основе
Схемотехника аналоговых электронных устройств. Практика 8. Операционные усилители и устройства на их основе Правомерное поведение, правонарушение и юридическая ответственность
Правомерное поведение, правонарушение и юридическая ответственность История фортепиано
История фортепиано Политические партии в России на рубеже XIX – XX веков
Политические партии в России на рубеже XIX – XX веков Теория Вечной Вселенной
Теория Вечной Вселенной Использование комплексов электронного голосования (КЭГ) на выборах депутатов Новгородской областной Думы 8 октября 2006 г. и на выбо
Использование комплексов электронного голосования (КЭГ) на выборах депутатов Новгородской областной Думы 8 октября 2006 г. и на выбо Общество с ограниченной ответственностью Нефть-Сервис Прокат. Соединитель волновой. Свидетельство о консервации
Общество с ограниченной ответственностью Нефть-Сервис Прокат. Соединитель волновой. Свидетельство о консервации Реализация Указа Президента РФ о долгосрочной государственной экономической политике. Город Курчатов
Реализация Указа Президента РФ о долгосрочной государственной экономической политике. Город Курчатов Презентация на тему Биотехнология
Презентация на тему Биотехнология Где жили динозавры? 1 класс
Где жили динозавры? 1 класс Мясо домашней птицы и дичи
Мясо домашней птицы и дичи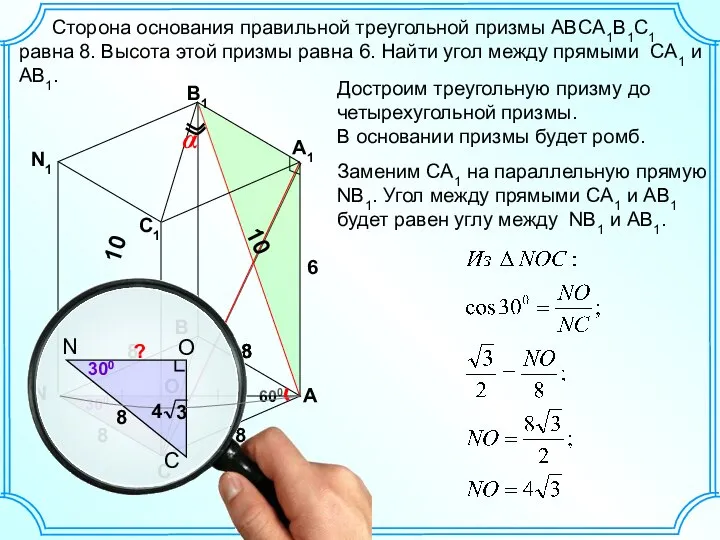 задача по пространственной геометрии
задача по пространственной геометрии Бенчмаркинг как путь к повышению эффективности компании Олег ДАВИДОВИЧ, Директор по маркетингу
Бенчмаркинг как путь к повышению эффективности компании Олег ДАВИДОВИЧ, Директор по маркетингу Брестская крепость"Я умираю, но не сдаюсь!"
Брестская крепость"Я умираю, но не сдаюсь!" Порядок подготовки и проведения государственной (итоговой) аттестации выпускников IХ классов в новой форме в 2010-2011 учебном году(П
Порядок подготовки и проведения государственной (итоговой) аттестации выпускников IХ классов в новой форме в 2010-2011 учебном году(П Ответственные родители
Ответственные родители AutoCAD ElectricalРешение задач энергетики
AutoCAD ElectricalРешение задач энергетики Интересная физика
Интересная физика Уильям Шекспир
Уильям Шекспир