- Дробление

Содержание
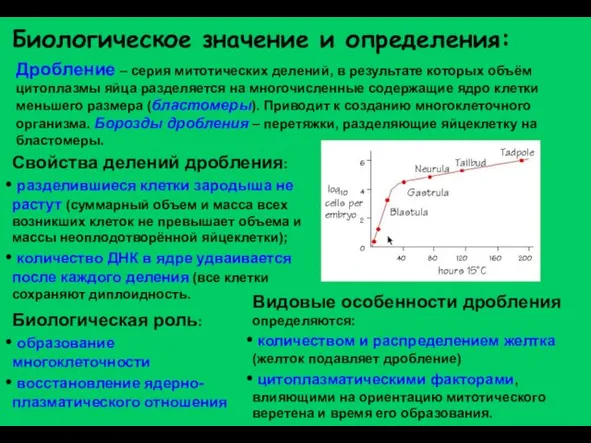
- 2. Дробление – серия митотических делений, в результате которых объём цитоплазмы яйца разделяется на многочисленные содержащие ядро
- 3. Отличие дробления от деления (1): В период дробления зиготы объем цитоплазмы не возрастает. С каждым следующим
- 4. Отличие дробления от деления (2): Два координированных процесса: кариокинез (митотическое деление ядра) – распределение хромосом клетки
- 5. Механизм дробления: Укорочение клеточных циклов при дроблении – в фазе G1 дробящихся эмбрионов отсутствует экспрессия генов,
- 6. Механизм дробления: Цитостатический фактор (CSF, продукт с-mos гена) блокирует развития мейоза, предотвращая деградацию циклина и сохраняя
- 7. Пространственная организация дробления: Правила Гертвига–Сакса (2): клеточное ядро стремится расположиться в центре чистой, свободной от желтка
- 8. По количеству желтка: алецитальные: безжелтковые (плацентарные млекопитающие, некоторые беспозвоночные – первичнотрахейные). олиголецитальные: маложелтковые (черви, моллюски, иглокожие).
- 9. По расположению (относительно полярной оси яйца) желтка: гомо- (изо-) лецитальные: желток в яйцеклетке распределен равномерно и
- 10. Классификация дробления: По характеру образования и расположению бластомеров: полное (голобластическое) – характерно для мезо- и изолецитальных
- 11. Голобластическое дробление: радиальное спиральное билатерально-симметричное чередующееся неправильное (анархическое) Классификация основана на взаимном пространственном расположении бластомеров Радиальный
- 12. Голобластическое дробление: Спиральный тип дробления характеризуется утерей элементов симметрии уже на стадии четырех, а иногда и
- 13. Билатеральный тип дробления (круглые черви, оболочники) характеризуется наличием одной плоскости симметрии. Голобластическое дробление: Анархический тип дробления
- 14. Меробластическое дробление: поверхностное и дискоидальное При поверхностном дроблении (насекомые) после слияния прону-клеусов ядро зиготы делится на
- 15. Зачастую, на ранних стадиях дробления внутренние концы бластомеров расходятся и между ними возникает небольшая, постепенно увеличивающаяся
- 16. Целобластула (от греч. koilos – пустой) характерна для иглокожих и ланцетника. Бластомеры на анимальном и вегетативном
- 17. Бластоциста (от греч. kystis – пузырь) характерна для млекопитающих. Образуется после прохождения стадии морулы. Представлена в
- 18. Дискобластула характерна для костистых рыб, рептилий и птиц. В результате дискоидального дробления образуется однослойная бластодерма (бластодиск),
- 19. Типы бластул (4): Перибластула характерна для насекомых и представляет собой заключительную стадию поверхностного дробления. Энергиды (ядра
- 21. Скачать презентацию
Слайд 2Дробление – серия митотических делений, в результате которых объём цитоплазмы яйца разделяется
Дробление – серия митотических делений, в результате которых объём цитоплазмы яйца разделяется
Слайд 3Отличие дробления от деления (1):
В период дробления зиготы объем цитоплазмы не возрастает.
С
Отличие дробления от деления (1):
В период дробления зиготы объем цитоплазмы не возрастает.
С

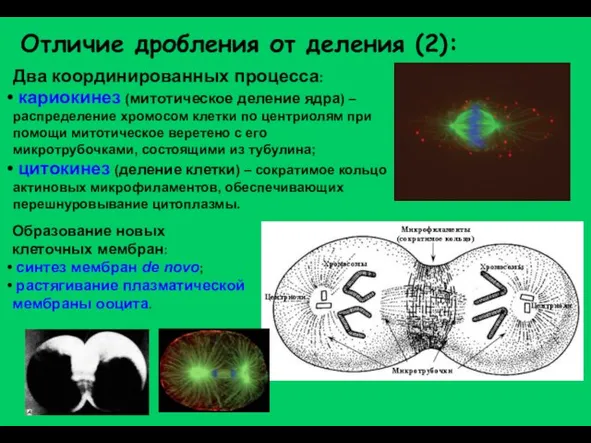
Слайд 4Отличие дробления от деления (2):
Два координированных процесса:
кариокинез (митотическое деление ядра) –
Отличие дробления от деления (2):
Два координированных процесса:
кариокинез (митотическое деление ядра) –
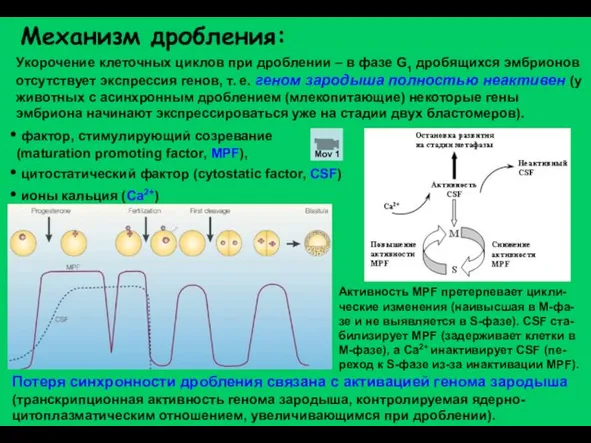
Слайд 5Механизм дробления:
Укорочение клеточных циклов при дроблении – в фазе G1 дробящихся эмбрионов
Механизм дробления:
Укорочение клеточных циклов при дроблении – в фазе G1 дробящихся эмбрионов
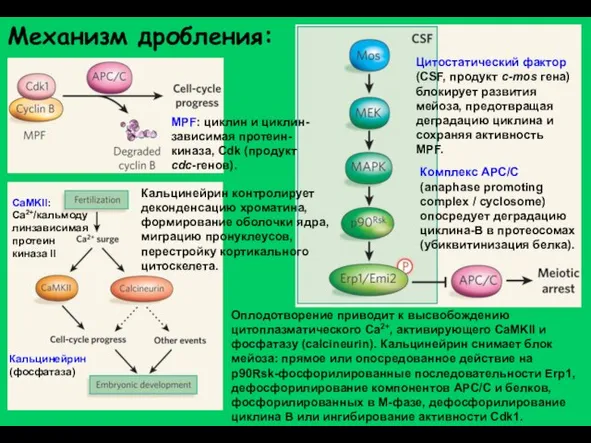
Слайд 6Механизм дробления:
Цитостатический фактор (CSF, продукт с-mos гена) блокирует развития мейоза, предотвращая деградацию
Механизм дробления:
Цитостатический фактор (CSF, продукт с-mos гена) блокирует развития мейоза, предотвращая деградацию
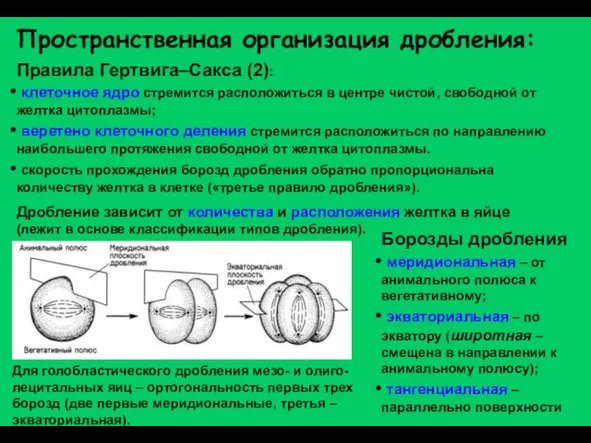
Слайд 7Пространственная организация дробления:
Правила Гертвига–Сакса (2):
клеточное ядро стремится расположиться в центре чистой,
Пространственная организация дробления:
Правила Гертвига–Сакса (2):
клеточное ядро стремится расположиться в центре чистой,
Слайд 8По количеству желтка:
алецитальные: безжелтковые (плацентарные млекопитающие, некоторые беспозвоночные – первичнотрахейные).
олиголецитальные:
По количеству желтка:
алецитальные: безжелтковые (плацентарные млекопитающие, некоторые беспозвоночные – первичнотрахейные).
олиголецитальные:

Слайд 9По расположению (относительно полярной оси яйца) желтка:
гомо- (изо-) лецитальные: желток в
По расположению (относительно полярной оси яйца) желтка:
гомо- (изо-) лецитальные: желток в

Слайд 10Классификация дробления:
По характеру образования и расположению бластомеров:
полное (голобластическое) – характерно для
Классификация дробления:
По характеру образования и расположению бластомеров:
полное (голобластическое) – характерно для

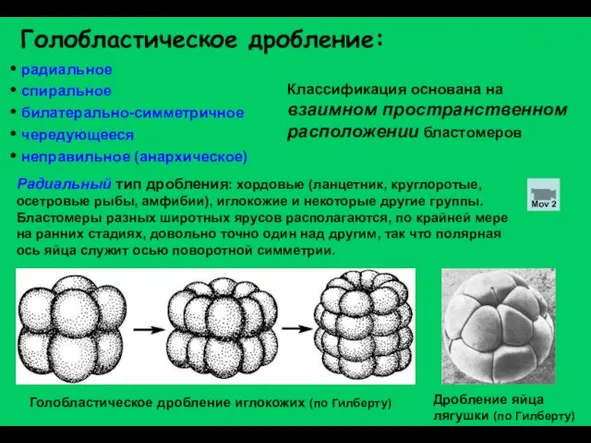
Слайд 11Голобластическое дробление:
радиальное
спиральное
билатерально-симметричное
чередующееся
неправильное (анархическое)
Классификация основана на взаимном пространственном
Голобластическое дробление:
радиальное
спиральное
билатерально-симметричное
чередующееся
неправильное (анархическое)
Классификация основана на взаимном пространственном
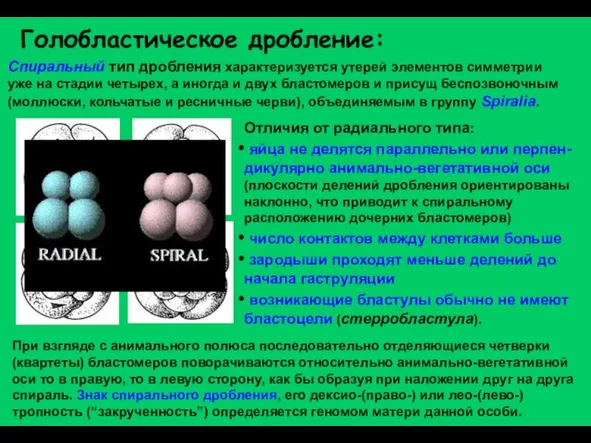
Слайд 12Голобластическое дробление:
Спиральный тип дробления характеризуется утерей элементов симметрии уже на стадии четырех,
Голобластическое дробление:
Спиральный тип дробления характеризуется утерей элементов симметрии уже на стадии четырех,
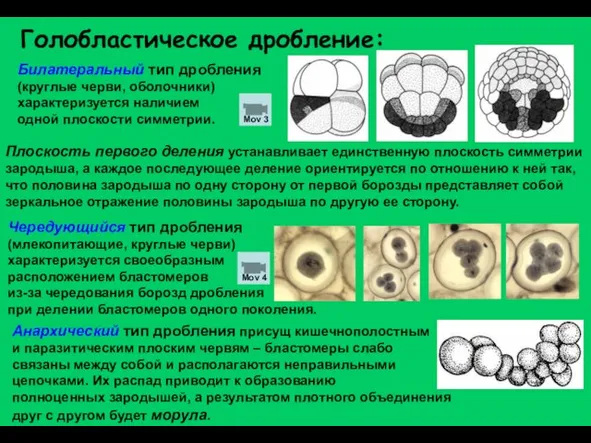
Слайд 13Билатеральный тип дробления (круглые черви, оболочники) характеризуется наличием
одной плоскости симметрии.
Голобластическое дробление:
Анархический
Билатеральный тип дробления (круглые черви, оболочники) характеризуется наличием
одной плоскости симметрии.
Голобластическое дробление:
Анархический
Слайд 14Меробластическое дробление:
поверхностное и дискоидальное
При поверхностном дроблении (насекомые) после слияния прону-клеусов ядро
Меробластическое дробление:
поверхностное и дискоидальное
При поверхностном дроблении (насекомые) после слияния прону-клеусов ядро

Слайд 15Зачастую, на ранних стадиях дробления внутренние концы бластомеров расходятся и между ними
Зачастую, на ранних стадиях дробления внутренние концы бластомеров расходятся и между ними

Слайд 16Целобластула (от греч. koilos – пустой) характерна для иглокожих и ланцетника. Бластомеры
Целобластула (от греч. koilos – пустой) характерна для иглокожих и ланцетника. Бластомеры
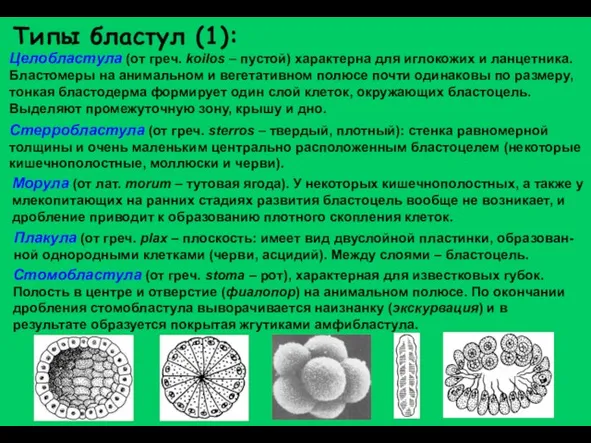
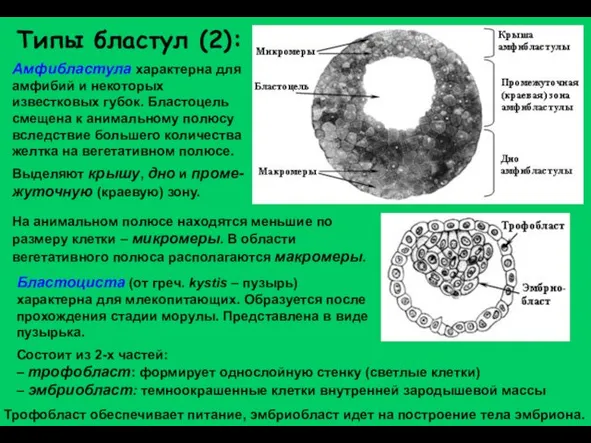
Слайд 17Бластоциста (от греч. kystis – пузырь) характерна для млекопитающих. Образуется после прохождения
Бластоциста (от греч. kystis – пузырь) характерна для млекопитающих. Образуется после прохождения
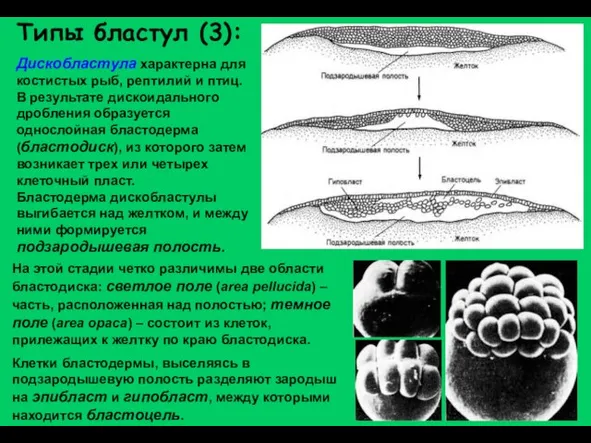
Слайд 18Дискобластула характерна для костистых рыб, рептилий и птиц. В результате дискоидального дробления
Дискобластула характерна для костистых рыб, рептилий и птиц. В результате дискоидального дробления
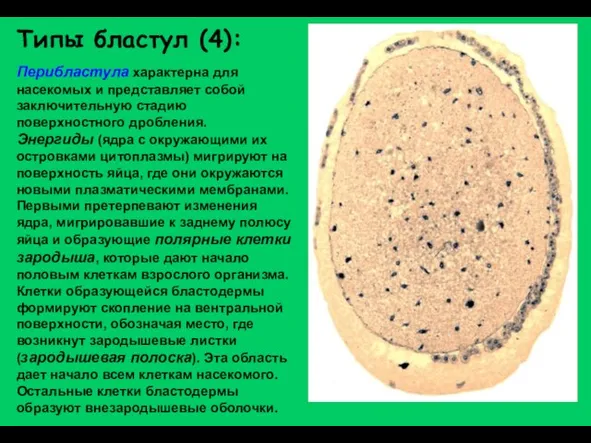
Слайд 19Типы бластул (4):
Перибластула характерна для насекомых и представляет собой заключительную стадию поверхностного
Типы бластул (4):
Перибластула характерна для насекомых и представляет собой заключительную стадию поверхностного
 Достопримечательности г. Омска
Достопримечательности г. Омска The Past Simple Tense Прошедшее простое время
The Past Simple Tense Прошедшее простое время Стратегия внедрения e-learning в ВУЗе Тихомирова Елена Владимировна Генеральный директор компании Центр eLearning Директор НИИ Управления
Стратегия внедрения e-learning в ВУЗе Тихомирова Елена Владимировна Генеральный директор компании Центр eLearning Директор НИИ Управления Презентация на тему Решение логарифмических уравнений
Презентация на тему Решение логарифмических уравнений Аральская средняя школа №14 им.Н.К.Крупской
Аральская средняя школа №14 им.Н.К.Крупской Презентация на тему Изложение текста "Кормушка" 4 класс
Презентация на тему Изложение текста "Кормушка" 4 класс Религиозно-рекреационный комплекс Шедруб линг в посёлке Косья
Религиозно-рекреационный комплекс Шедруб линг в посёлке Косья Инструктаж по поведению на льду
Инструктаж по поведению на льду Распознавание спряжения глаголов по неопределённой форме
Распознавание спряжения глаголов по неопределённой форме Налог на доходы физических лиц
Налог на доходы физических лиц Витражная аппликация
Витражная аппликация Δευτερο μάθημα. Посвящается году Греции в России
Δευτερο μάθημα. Посвящается году Греции в России Проектирование электрического освещения
Проектирование электрического освещения Натюрморт с рыбой и хлебом
Натюрморт с рыбой и хлебом Низов Сергей Валерьевич начальник управления образования Администрации Ивановского муниципального района
Низов Сергей Валерьевич начальник управления образования Администрации Ивановского муниципального района Proofreading
Proofreading Глагол
Глагол Презентация на тему Дед Мороз и Санта Клаус
Презентация на тему Дед Мороз и Санта Клаус Лазарев Дмитрий
Лазарев Дмитрий Краткий очерк истории развития химии
Краткий очерк истории развития химии Есть сказка, которая называется "Принцесса или тигр?" В этой сказке один узник должен угадать, в какой из двух комнат находится прин
Есть сказка, которая называется "Принцесса или тигр?" В этой сказке один узник должен угадать, в какой из двух комнат находится прин Бухгалтерская (финансовая) отчетность. Электронный курс лекций
Бухгалтерская (финансовая) отчетность. Электронный курс лекций Публичная презентация профессиональному и местному сообществу результатов педагогической деятельности и инновационной работы
Публичная презентация профессиональному и местному сообществу результатов педагогической деятельности и инновационной работы 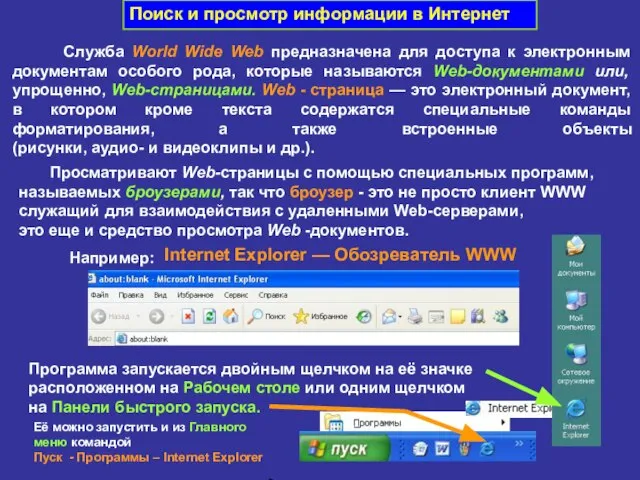 Поиск и просмотр информации в Интернет
Поиск и просмотр информации в Интернет 04_ASBIS_Almaty_MS ppt
04_ASBIS_Almaty_MS ppt Миссия нашего бренда. Befagor
Миссия нашего бренда. Befagor Что ценного дает коучинг?
Что ценного дает коучинг? Ахматова
Ахматова