- ФИЗИОЛОГИЯ НЕРВОВ И МЫШЦ

Содержание
- 2. ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ Нервное волокно (нервный проводник) представляет собой отросток нейрона, заключенный в глиальную
- 3. Морфологическая организация нервных проводников Различают два вида нервных волокон: Немиелинизированные (безмякотные). Миелинизированные (мякотные)
- 4. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А–Г) Соотношение леммоцита и безмиелиновых волокон
- 5. Классификация нервных волокон по Эрлангеру-Гассеру
- 6. МЕХАНИЗМ ПРОВЕДЕНИЯ В НЕРВНЫХ ВОЛОКНАХ Проводимость – специализированное свойство нервного волокна. Возбуждение распространяется посредством электротонической связи
- 7. Законы проведения возбуждения в нервных волокнах 1. Закон двустороннего проведения. 2. Закон анатомической и физиологической целостности.
- 8. 1. Закон двустороннего проведения Возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места
- 9. 2. Закон анатомической и физиологической целостности Возбуждение может распространяться по нервному волокну только в случае его
- 10. 3. Закон изолированного проведения Возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние
- 11. 4. Закон бездекрементного проведения Амплитуда потенциала действия не изменяется с увеличением расстояния от места его возникновения.
- 12. ФИЗИОЛОГИЯ МЫШЦ
- 13. Физиологические свойства мышц Кроме общий свойств, характерных для всех возбудимых систем - возбудимость, проводимость, лабильность (200
- 14. Скелетные мышцы Мышечное волокно – многоядерное образование, имеет: плазматическую мембрану (сарколемму) от которой внутрь отходят многочисленные
- 15. Соотношение актиновых и миозиновых нитей в саркомере
- 16. Строение актиновых нитей
- 17. Строение молекулы миозина АТФ – аденозинтрифосфат; НММ – тяжелые цепи меромиозина; LMM – легкие цепи меромиозина;
- 18. Механизм развития сокращения мышцы При сокращении актиновые и миозиновые нити не укорачиваются, а скользят относительно друг
- 19. Электромеханическое сопряжение Контрактура - стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может
- 20. Механизм «гребка» при развитии мышечного сокращения
- 21. ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ В зависимости от условий, в которых мышца осуществляет сокращение, принято различать следующие типы
- 22. ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ При раздражении одиночным стимулом мышца отвечает одиночным сокращением, в котором выделяют три фазы:
- 23. Тетаническое сокращение Зубчатый – если очередной импульс приходится на фазу расслабления - происходит неполная суммация. Гладкий
- 24. ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ Мышцы иннервируются мотонейронами. Каждый аксон мотонейрона иннервирует группу мышечных волокон. Мотонейрон вместе с иннервируемыми
- 25. Мышечные волокна, которые входят в состав двигательных единиц, неоднородны Волокна, входящие в состав одной мышцы, могут
- 26. Физические свойства скелетной мышцы 1. Растяжимость – способность изменять длину под действием растягивающей силы. 2. Эластичность
- 27. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений,
- 28. ГЛАДКИЕ МЫШЦЫ Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток, связанных между собой нексусами. Нексусы обеспечивают
- 29. ОСОБЕННОСТИ ГЛАДКИХ МЫШЦ 1. Распространение возбуждения происходит за счет нексусов. Эти области с низким электрическим сопротивлением
- 31. Скачать презентацию
Слайд 2ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ
Нервное волокно (нервный проводник) представляет собой отросток нейрона,
ФИЗИОЛОГИЯ НЕРВОВ ИЛИ НЕРВНЫХ ПРОВОДНИКОВ
Нервное волокно (нервный проводник) представляет собой отросток нейрона,

Слайд 3Морфологическая организация
нервных проводников
Различают два вида нервных волокон:
Немиелинизированные (безмякотные).
Миелинизированные (мякотные)
Морфологическая организация
нервных проводников
Различают два вида нервных волокон:
Немиелинизированные (безмякотные).
Миелинизированные (мякотные)

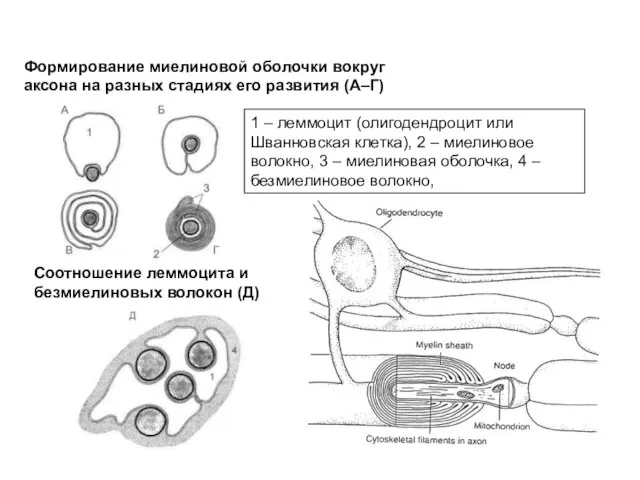
Слайд 4Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А–Г)
Соотношение леммоцита
Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А–Г)
Соотношение леммоцита
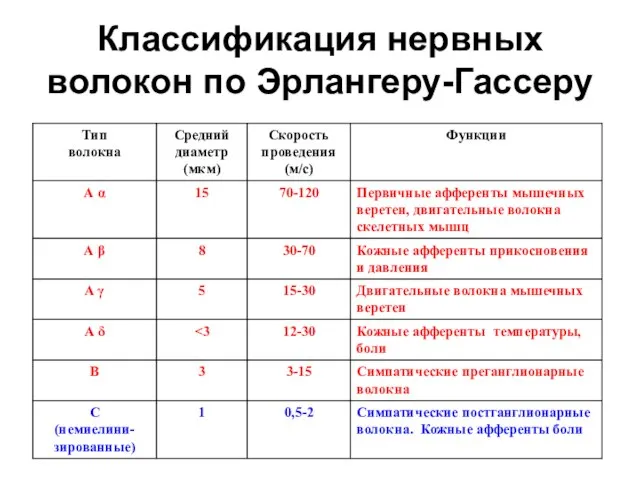
Слайд 5Классификация нервных волокон по Эрлангеру-Гассеру
Классификация нервных волокон по Эрлангеру-Гассеру
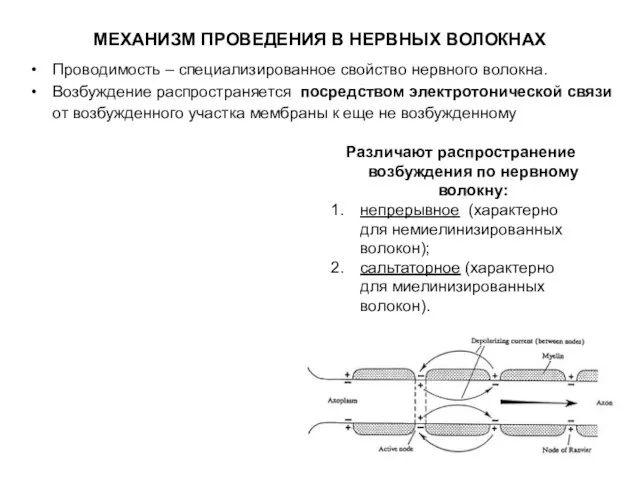
Слайд 6МЕХАНИЗМ ПРОВЕДЕНИЯ В НЕРВНЫХ ВОЛОКНАХ
Проводимость – специализированное свойство нервного волокна.
Возбуждение распространяется посредством
МЕХАНИЗМ ПРОВЕДЕНИЯ В НЕРВНЫХ ВОЛОКНАХ
Проводимость – специализированное свойство нервного волокна.
Возбуждение распространяется посредством
Слайд 7Законы проведения возбуждения в нервных волокнах
1. Закон двустороннего проведения.
2. Закон анатомической
Законы проведения возбуждения в нервных волокнах
1. Закон двустороннего проведения.
2. Закон анатомической

Слайд 81. Закон двустороннего проведения
Возбуждение, возникающее в одном участке нерва, распространяется в обе
1. Закон двустороннего проведения
Возбуждение, возникающее в одном участке нерва, распространяется в обе

Слайд 92. Закон анатомической и физиологической целостности
Возбуждение может распространяться по нервному волокну только
2. Закон анатомической и физиологической целостности
Возбуждение может распространяться по нервному волокну только

Слайд 103. Закон изолированного проведения
Возбуждение, распространяющееся по волокну, входящему в состав нерва, не
3. Закон изолированного проведения
Возбуждение, распространяющееся по волокну, входящему в состав нерва, не

Слайд 114. Закон бездекрементного проведения
Амплитуда потенциала действия не изменяется с увеличением расстояния от
4. Закон бездекрементного проведения
Амплитуда потенциала действия не изменяется с увеличением расстояния от

Слайд 12ФИЗИОЛОГИЯ МЫШЦ
ФИЗИОЛОГИЯ МЫШЦ

Слайд 13Физиологические свойства мышц
Кроме общий свойств, характерных для всех возбудимых систем - возбудимость,
Физиологические свойства мышц
Кроме общий свойств, характерных для всех возбудимых систем - возбудимость,
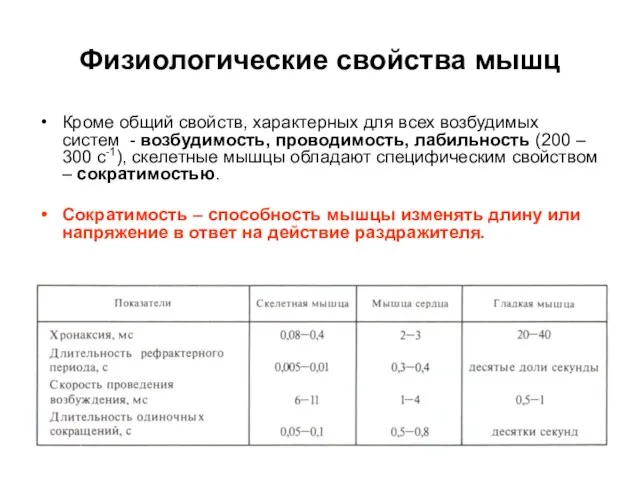
Слайд 14Скелетные мышцы
Мышечное волокно – многоядерное образование, имеет:
плазматическую мембрану (сарколемму) от которой внутрь
Скелетные мышцы
Мышечное волокно – многоядерное образование, имеет:
плазматическую мембрану (сарколемму) от которой внутрь
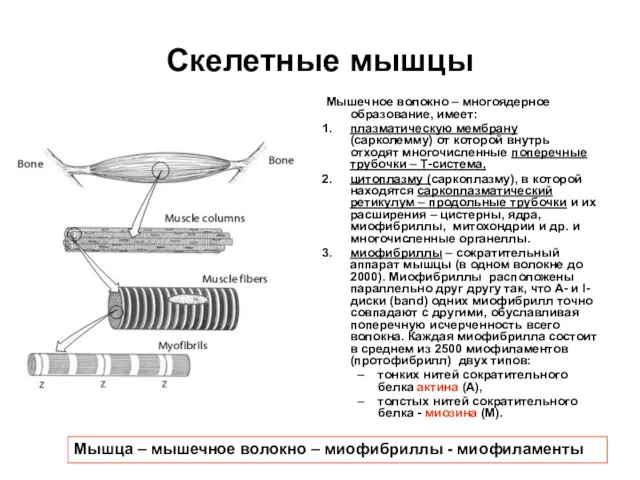
Слайд 15Соотношение актиновых и миозиновых нитей в саркомере
Соотношение актиновых и миозиновых нитей в саркомере
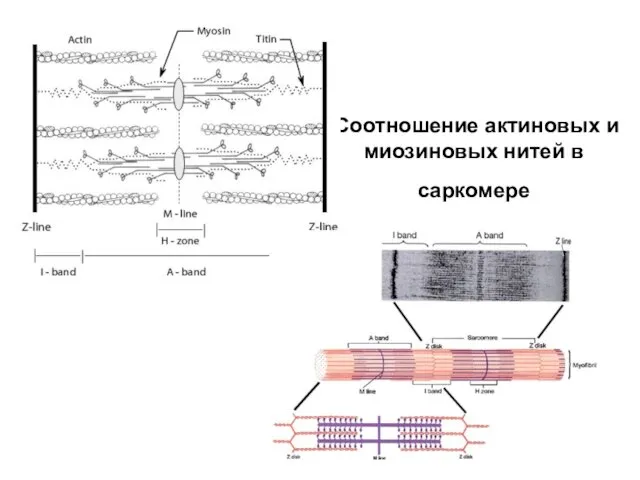
Слайд 16Строение актиновых нитей
Строение актиновых нитей
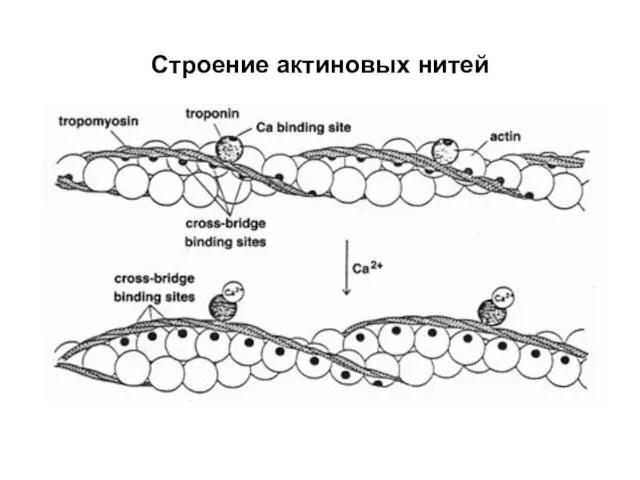
Слайд 17Строение молекулы миозина
АТФ – аденозинтрифосфат;
НММ – тяжелые цепи меромиозина;
LMM –
Строение молекулы миозина
АТФ – аденозинтрифосфат;
НММ – тяжелые цепи меромиозина;
LMM –
Слайд 18Механизм развития сокращения мышцы
При сокращении актиновые и миозиновые нити не укорачиваются,
Механизм развития сокращения мышцы
При сокращении актиновые и миозиновые нити не укорачиваются,

Слайд 19Электромеханическое сопряжение
Контрактура - стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя.
Электромеханическое сопряжение
Контрактура - стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя.

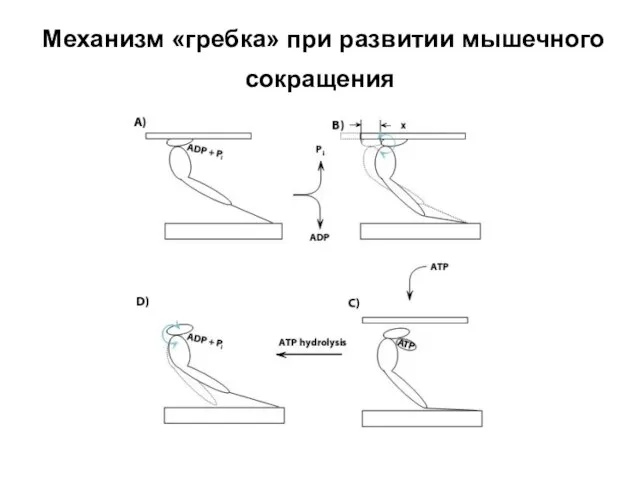
Слайд 20Механизм «гребка» при развитии мышечного сокращения
Механизм «гребка» при развитии мышечного сокращения
Слайд 21ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
В зависимости от условий, в которых мышца осуществляет сокращение, принято
ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
В зависимости от условий, в которых мышца осуществляет сокращение, принято

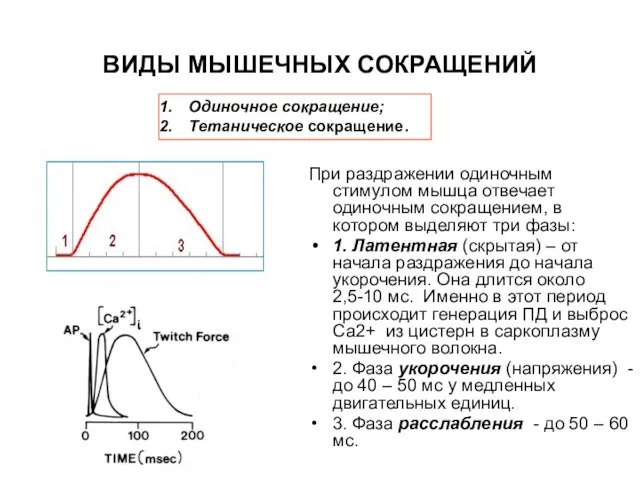
Слайд 22ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
При раздражении одиночным стимулом мышца отвечает одиночным сокращением, в
ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
При раздражении одиночным стимулом мышца отвечает одиночным сокращением, в
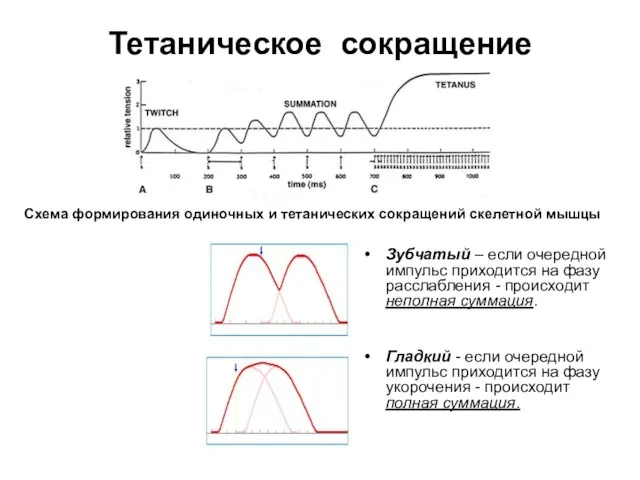
Слайд 23Тетаническое сокращение
Зубчатый – если очередной импульс приходится на фазу расслабления -
Тетаническое сокращение
Зубчатый – если очередной импульс приходится на фазу расслабления -
Слайд 24ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
Мышцы иннервируются мотонейронами. Каждый аксон мотонейрона иннервирует группу мышечных волокон.
Мотонейрон
ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ
Мышцы иннервируются мотонейронами. Каждый аксон мотонейрона иннервирует группу мышечных волокон.
Мотонейрон

Слайд 25Мышечные волокна, которые входят в состав двигательных единиц, неоднородны
Волокна, входящие в состав
Мышечные волокна, которые входят в состав двигательных единиц, неоднородны
Волокна, входящие в состав

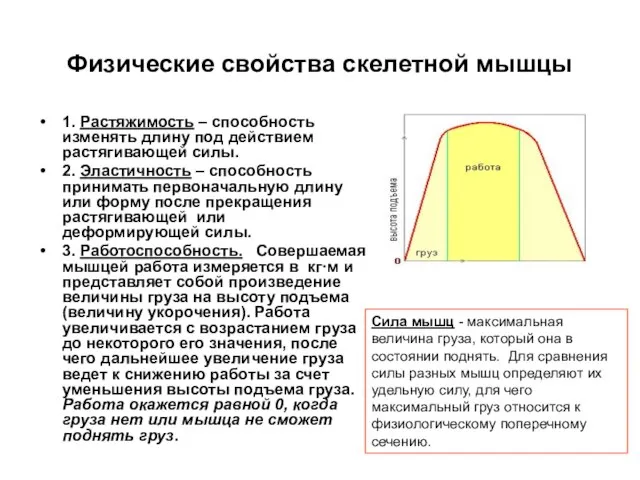
Слайд 26Физические свойства скелетной мышцы
1. Растяжимость – способность изменять длину под действием
Физические свойства скелетной мышцы
1. Растяжимость – способность изменять длину под действием
Слайд 27Утомление
– физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется
Утомление
– физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется

Слайд 28ГЛАДКИЕ МЫШЦЫ
Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток, связанных между собой
ГЛАДКИЕ МЫШЦЫ
Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток, связанных между собой

Слайд 29ОСОБЕННОСТИ ГЛАДКИХ МЫШЦ
1. Распространение возбуждения происходит за счет нексусов. Эти области с
ОСОБЕННОСТИ ГЛАДКИХ МЫШЦ
1. Распространение возбуждения происходит за счет нексусов. Эти области с

 Числительное. Употребление Грамматических форм
Числительное. Употребление Грамматических форм История античной философии
История античной философии «Зимний запас» по Н.Сладкову
«Зимний запас» по Н.Сладкову Урок презентация на тему Азбука – первая учебная книга
Урок презентация на тему Азбука – первая учебная книга  Конституция РФ
Конституция РФ Цифровая схемотехника. Светодиодные семисегментные индикаторы
Цифровая схемотехника. Светодиодные семисегментные индикаторы Гибкость и ее развитие. Две формы гибкости
Гибкость и ее развитие. Две формы гибкости Деловое общение
Деловое общение Непревзойденные русские сказочники
Непревзойденные русские сказочники Пригласительный билет
Пригласительный билет Дифференциация звуков и букв Ф и В
Дифференциация звуков и букв Ф и В Самовосстанавливающийся бетон
Самовосстанавливающийся бетон МаловаЕлена ВалентиновнаМДОУ №9 «Сказка»
МаловаЕлена ВалентиновнаМДОУ №9 «Сказка» Отраслевые решения «1С» для сельского хозяйства и пищевой промышленности в ВУЗЫ
Отраслевые решения «1С» для сельского хозяйства и пищевой промышленности в ВУЗЫ Классификация ЧС техногенного характера
Классификация ЧС техногенного характера Запуск интернет- стартапов, вывод на рынок Максим Спиридонов www.spiridonov.ruwww.spiridonov.ru «Продюсерский центр «Ройбер» www.roiber.ruwww.roiber.ru.
Запуск интернет- стартапов, вывод на рынок Максим Спиридонов www.spiridonov.ruwww.spiridonov.ru «Продюсерский центр «Ройбер» www.roiber.ruwww.roiber.ru. Как подготовить конспект урока
Как подготовить конспект урока Устное решение некоторых квадратных уравнений
Устное решение некоторых квадратных уравнений SeasteadingГомстед нейтральных вод
SeasteadingГомстед нейтральных вод Королева С.В
Королева С.В Презентация на тему Правописание НЕ с причастиями
Презентация на тему Правописание НЕ с причастиями Добавки для ненасыщенных полиэфирных ивинил-эфирных систем
Добавки для ненасыщенных полиэфирных ивинил-эфирных систем Великие открытия Христофор Колумб
Великие открытия Христофор Колумб Званый ужин
Званый ужин Закон сохранения механической энергии
Закон сохранения механической энергии сочинение по картинке "Зимние забавы детей"
сочинение по картинке "Зимние забавы детей" История развития электронных двигателей
История развития электронных двигателей Язык в координатах массмедиа: лексика пандемии. Как вирус заражает язык
Язык в координатах массмедиа: лексика пандемии. Как вирус заражает язык