- ОТ ГЕНОВ К ПРИЗНАКАМ

Содержание
- 2. 1. Введение После переоткрытия в 1900 законов Менделя году актуальными стали следующие вопросы: 1. Как генотип
- 3. 2. Наследственные нарушения метаболизма у человека (исследования Гэррода); 2.1. Врождённые ошибки метаболизма В 1902-09 гг. А.
- 4. В 1914 г. было показано, что у больных алкаптонурией отсут-ствует активность фермента - оксидазы гомогентизиновой кис-лоты,
- 5. 3. Neurosроra crassa – объект биохимической генетики В 30-х гг. XX в генетики хорошо понимали, что
- 6. Принципы подхода Бидла и Татума Бидл и Татум предположили, что обработка рентгеновскими лучами вызовет мутации в
- 7. Рис. 2. Жизненный цикл Neurosроra crassa Мицелий гриба растет на твердой питательной среде в виде сплетения
- 8. ми противоположных типов спаривания. Тип спаривания контролируется двумя аллелями, А и а в одной из хромосом.
- 9. В обычных условиях все ядра в гифах имеют один и тот же генотип (рис.2,В). Гифы с
- 10. Исследования ауксотрофных мутантов Neurosроra crassa Для повышения частоты мутаций споры облучали рент-геновскими лучами и проращивали их
- 11. Рис. 3. Метод выявления ауксотрофных мутаций Конидии облучают (для увеличения частоты мутаций), проращивают на полной среде,
- 12. Рис.4. Доказательство генетической природы ауксотрофности мутанов Neurospora. Мутантный штамм, выявленный как показано на рис. 3, скрещивают
- 13. Бидл и Татум получили много разных мутантов, у которых были нарушены разные этапы биосинтеза того или
- 14. 5. Ген – фермент - биохимическая реакция Различные ауксотрофные штаммы, требующие для своего роста добавки одного
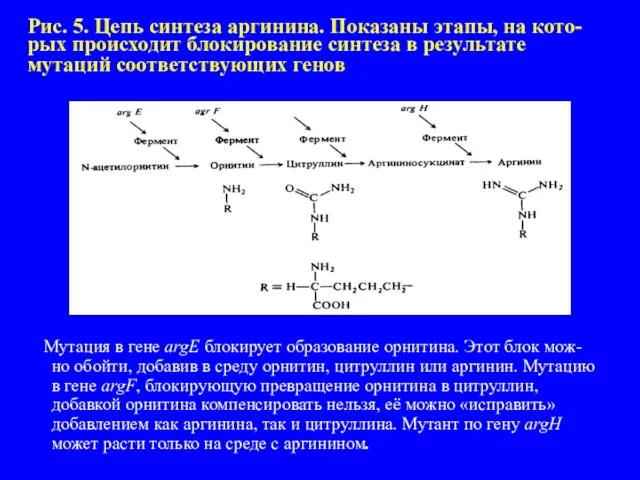
- 15. Рис. 5. Цепь синтеза аргинина. Показаны этапы, на кото-рых происходит блокирование синтеза в результате мутаций соответствующих
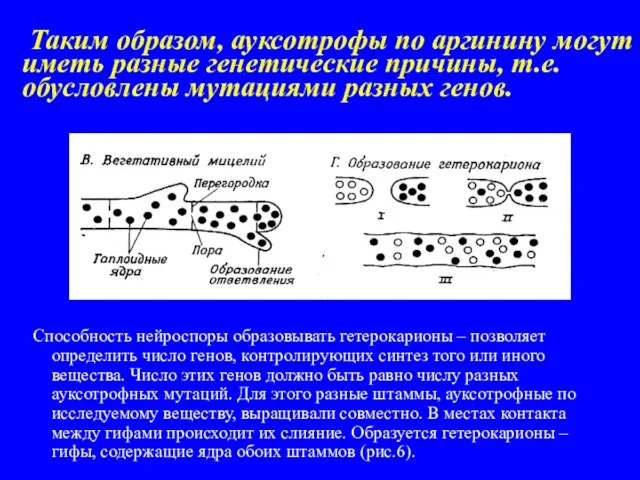
- 16. Таким образом, ауксотрофы по аргинину могут иметь разные генетические причины, т.е. обусловлены мутациями разных генов. Способность
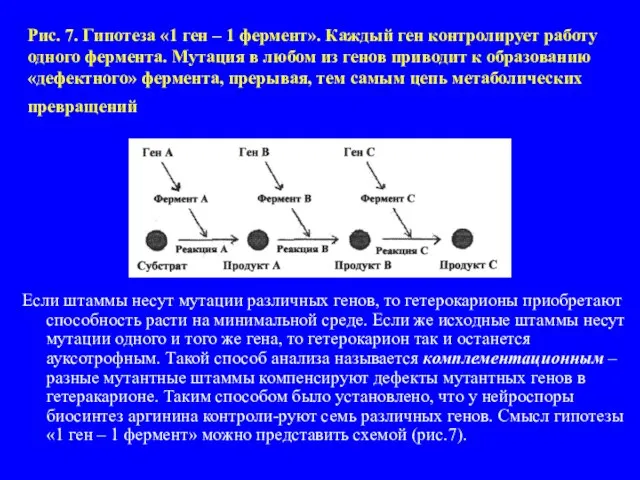
- 17. Рис. 7. Гипотеза «1 ген – 1 фермент». Каждый ген контролирует работу одного фермента. Мутация в
- 18. Заключение В настоящее время мы знаем, что теория «один ген — один фермент» была упрощением: не
- 20. Скачать презентацию

Слайд 3
2. Наследственные нарушения метаболизма у человека (исследования Гэррода);
2.1. Врождённые ошибки метаболизма
2. Наследственные нарушения метаболизма у человека (исследования Гэррода);
2.1. Врождённые ошибки метаболизма

Слайд 4 В 1914 г. было показано, что у больных алкаптонурией отсут-ствует активность
В 1914 г. было показано, что у больных алкаптонурией отсут-ствует активность

Слайд 5
3. Neurosроra crassa – объект биохимической генетики
В 30-х гг. XX в
3. Neurosроra crassa – объект биохимической генетики
В 30-х гг. XX в

Слайд 6Принципы подхода Бидла и Татума
Бидл и Татум предположили, что обработка рентгеновскими
Принципы подхода Бидла и Татума
Бидл и Татум предположили, что обработка рентгеновскими

Слайд 7Рис. 2. Жизненный цикл Neurosроra crassa
Мицелий гриба растет на твердой
Рис. 2. Жизненный цикл Neurosроra crassa
Мицелий гриба растет на твердой

Слайд 8ми противоположных типов спаривания. Тип спаривания контролируется двумя аллелями, А и а
ми противоположных типов спаривания. Тип спаривания контролируется двумя аллелями, А и а

Слайд 9 В обычных условиях все ядра в гифах имеют один и тот
В обычных условиях все ядра в гифах имеют один и тот

Слайд 10Исследования ауксотрофных мутантов Neurosроra crassa
Для повышения частоты мутаций споры облучали
Исследования ауксотрофных мутантов Neurosроra crassa
Для повышения частоты мутаций споры облучали

Слайд 11Рис. 3. Метод выявления ауксотрофных мутаций
Конидии облучают (для увеличения частоты мутаций), проращивают
Рис. 3. Метод выявления ауксотрофных мутаций
Конидии облучают (для увеличения частоты мутаций), проращивают

Слайд 12Рис.4. Доказательство генетической природы ауксотрофности мутанов Neurospora.
Мутантный штамм, выявленный как показано
Рис.4. Доказательство генетической природы ауксотрофности мутанов Neurospora.
Мутантный штамм, выявленный как показано

Слайд 13 Бидл и Татум получили много разных мутантов, у которых были нарушены
Бидл и Татум получили много разных мутантов, у которых были нарушены

Слайд 14
5. Ген – фермент - биохимическая реакция
Различные ауксотрофные штаммы, требующие для
5. Ген – фермент - биохимическая реакция
Различные ауксотрофные штаммы, требующие для

Слайд 15Рис. 5. Цепь синтеза аргинина. Показаны этапы, на кото-рых происходит блокирование синтеза
Рис. 5. Цепь синтеза аргинина. Показаны этапы, на кото-рых происходит блокирование синтеза
Слайд 16 Таким образом, ауксотрофы по аргинину могут иметь разные генетические причины, т.е.
Таким образом, ауксотрофы по аргинину могут иметь разные генетические причины, т.е.
Слайд 17Рис. 7. Гипотеза «1 ген – 1 фермент». Каждый ген контролирует работу
Рис. 7. Гипотеза «1 ген – 1 фермент». Каждый ген контролирует работу
Слайд 18 Заключение
В настоящее время мы знаем, что теория «один ген —
Заключение В настоящее время мы знаем, что теория «один ген —

 Сера
Сера 20171210_pervye_evropeytsy_na_krayu_azii
20171210_pervye_evropeytsy_na_krayu_azii ГРИПП и другие острые респираторные заболевания- ОРЗ
ГРИПП и другие острые респираторные заболевания- ОРЗ Интеграция Бренда в фестиваль Petshop
Интеграция Бренда в фестиваль Petshop Комплексная оптимизация городского транспорта с использованием искусственных нейронных сетей
Комплексная оптимизация городского транспорта с использованием искусственных нейронных сетей День Конституции России
День Конституции России Рекурсивные алгоритмы ЕГЭ-2017 по информатике Задание 11
Рекурсивные алгоритмы ЕГЭ-2017 по информатике Задание 11  "Одурманенные травкой"
"Одурманенные травкой" CIT CTF
CIT CTF 20171012_kak_nauchit_shkolnikov_samostoyatelno_rabotat_s_tekstom_uchebnika
20171012_kak_nauchit_shkolnikov_samostoyatelno_rabotat_s_tekstom_uchebnika Организация медицинского освидетельствования граждан при постановке их на воинский учет
Организация медицинского освидетельствования граждан при постановке их на воинский учет Пауль Клее. Ангел истории
Пауль Клее. Ангел истории Презентация на тему Полисахариды
Презентация на тему Полисахариды Творческий - исследовательский проект по технологии Итальянская Кухня. Пицца
Творческий - исследовательский проект по технологии Итальянская Кухня. Пицца Обзор жизни и творчества А.И. Солженицына
Обзор жизни и творчества А.И. Солженицына Язык поисковых запросов
Язык поисковых запросов Раскраска от студии. Чем пахнут ремесла?
Раскраска от студии. Чем пахнут ремесла? Зачем у флейты дырочки?
Зачем у флейты дырочки? Энергоэффективная экономика начинается с точных измерений
Энергоэффективная экономика начинается с точных измерений Обособленные определения и обстоятельства
Обособленные определения и обстоятельства Д-р. Петра ОпитцРазвитие малой и вознобновляемой энергетики: презентация немецких технологий и возможности кооперации Москва, 23.
Д-р. Петра ОпитцРазвитие малой и вознобновляемой энергетики: презентация немецких технологий и возможности кооперации Москва, 23. ЛУЧШЕ БОРОТЬСЯ С БОГОМ ЛИЦОМ К ЛИЦУ, ЧЕМ ДРУЖИТЬ НА РАССТОЯНИИ Бытие 32:22-31.
ЛУЧШЕ БОРОТЬСЯ С БОГОМ ЛИЦОМ К ЛИЦУ, ЧЕМ ДРУЖИТЬ НА РАССТОЯНИИ Бытие 32:22-31. Разный пол - разные мысли.
Разный пол - разные мысли. п.Советский 2007
п.Советский 2007 Студия маникюра и педикюра Малышка
Студия маникюра и педикюра Малышка Важность преаналитического этапа для лабораторных исследованийЗаведующая клинико-диагностической лаборатории, к.б.н. Сон В.Н.
Важность преаналитического этапа для лабораторных исследованийЗаведующая клинико-диагностической лаборатории, к.б.н. Сон В.Н. Презентация на тему Рельеф России
Презентация на тему Рельеф России  Презентация на тему Викторина по чтению В гостях у сказки
Презентация на тему Викторина по чтению В гостях у сказки