- Презентация стероиды и вит. биотехн.

Содержание
- 2. Стероиды Циклопентанпергидрофенантрен К стеринам (стеролам) относятся стероиды, несущие в положении С-3 гидроксильную группу.
- 3. Гормоны – это БАВ, выделяемые железами внутренней секреции и являющиеся регуляторами обмена веществ. По химической природе:
- 4. Холестерин (класс зоостеринов), С27Н46О. Обнаруживается почти во всех органах и тканях животных и человека. Желчные камни
- 5. Стерины растений (фитостерины) Эргостерин по структуре отличается от холестерина дополнительной метильной группой в боковой цепи при
- 6. Стигмастерин С29H48О содержится в большом количестве в соевом масле и сахарном тростнике. По структуре стигмастерин отличается
- 7. Ситостерин С29Н50О. В хлопковом и соевом маслах, в зародышах пшеницы и натуральном каучуке, в сахарном тростнике
- 8. В основе истории синтеза стероидных гормонов лежат методы биотрансформации, результатом применения которых является превращение метаболитов в
- 9. Применение микроорганизмов в качестве носителей активных полиферментных систем, способных переводить экзогенные органические соединения в разнообразные полезные
- 10. Микробиологическая трансформация – использование ферментативной активности жизнеспособных клеток микроорганизмов, результатом чего является некоторое изменение молекулярной структуры
- 11. Еще в конце XIX в. было известно, что бактериальная флора кишечника млекопитающих превращает холестерин в копростерин,
- 12. В настоящее время принята классификация микробиологических трансформаций по типу возникновения и отщепления функциональных групп. окисление, восстановление,
- 14. Микробиологическое гидроксилирование Наличие гидроксильных групп в 3, 11, 16, 17 положениях молекулы стероида, как правило, обусловливает
- 15. Наличие в молекуле стероидов 11-гидроксильной группы обусловливает физиологическую активность гидрокортизона (кортизола) и преднизолона. Гидроксилированию подвергаются субстраты
- 16. Дегидрогенизация стероидов Преднизолон из кортизона и гидрокортизона, дианабол из метилтестостерона. Чаще всего микроорганизмы дегидрируют положения 1,2
- 17. Микробиологическое восстановление Этот процесс используется в меньшей степени, чем дегидрирование. Он осуществляется главным образом дрожжами и
- 18. Окисление гидроксильной группы в кетогруппу Одна из наиболее частых реакций, осуществляемых микроорганизмами (бактериями, актиномицетами, грибами). Наибольший
- 19. Гидролиз эфиров стероидов Открыт в 1938 г. Практическая ценность этой реакции определяется тем, что ацилированные стероиды
- 20. Дезацилирующая способность часто встречается среди микоформ, мукоровых и несовершенных грибов, актиномицетов. Особенность приведенной реакции состоит в
- 21. Отщепление боковых цепей стероидов Проблема расщепления боковой цепи стеринов с сохранением стероидного скелета может быть решена
- 22. Технологические особенности получения стероидных гормонов Преимущества иммобилизации Особенности подачи субстрата Получение целевого продукта Основные пути усовершенствования
- 23. Получение гидрокортизона (кортизола) из вещества S 1. Выращивание трансформирующей культуры (I стадия) производят путем трех последовательных
- 24. 2. Трансформация вещества S также начинается со стерилизации ферментера и воздушного фильтра водным раствором формалина. Размол
- 25. 3. Выделение гидрокортизона. Культуральная жидкость вместе с мицелием поступает на сепарацию. Отделенный мицелий промывается, промывные воды
- 26. Витамины Жизненно важные низкомолекулярные органические соединения, необходимые для человека в ничтожных количествах, но имеющие огромное значение.
- 27. История открытия В сентябре 1880 г. при защите своей докторской диссертации Лунин утверждал, что для сохранения
- 28. В 1906 году Фредерик Хопкинс предположил, что помимо белков, жиров, углеводов и т. д., пища содержит
- 29. Функции и свойства витаминов: 1. Биостимуляция ферментативных процессов 2. Составная часть коферментов (не всегда) 3. Антимутагенная
- 30. Поскольку бактерии для своего роста и размножения нуждаются в присутствии многих витаминов для синтеза коферментов, введение
- 31. Получение витамина В2 (рибофлавина) Для рибофлавина характерно функционирование в коэнзимных формах: -флавиномононуклеотид (ФМН) -флавинадениндинуклеотид (ФАД). К
- 32. Активным продуцентом рибофлавина являются культуры дрожжеподобных грибов Eremothecium ashbyii и Ashbya gossipii, синтезирующих до 3,8 и
- 33. После выращивания в инокуляторе в течение 21 – 26 часов культуру переводят в ферментёр с питательной
- 34. Именно на примере выделения рибофлавина в культуральную жидкость было открыто явление сверхсинтеза. Сверхсинтез рибофлавина можно получить,
- 35. Bacillus subtilis В 1983 году в институте генетики был сконструирован рекомбинантный штамм продуцента Bacillus subtilis, способный
- 36. Меласса (чёрная патока, фр. Mélasse) Кормовая патока, побочный продукт сахарного производства; сиропообразная жидкость тёмно-бурого цвета со
- 37. Розеофлавин 8-диметиламино(нор)-рибофлавин – антибиотик, образуемый Streptomyces davawensis. Показано, что розеофлавин ингибирует синтез 6,7-диметил-8- рибитиллюмазина и рибофлавинсинтетазу
- 41. Получение витамина В12 Этот витамин был открыт одновременно в США и в Англии. В 1972 г.
- 42. Продуцентом витамина В12 являются пропионовокислые бактерии из рода Propionibacterium. Продуктивность – более 50 мг на 1
- 43. По окончании ферментации витамин остаётся в клетках, поэтому полученную массу сепарируют, стабилизируют нитритом натрия, охлаждают, нейтрализуют,
- 46. Витамин С Одним из классических примеров промышленного использования микроорганизмов в получении витамина С является превращение D-сорбита
- 47. Наиболее сильной окислительной способностью обладают штаммы Acetobacter melanogenum и Gluconobacter oxydans. Для получения сорбозы Gluconobacter oxydans
- 48. Эргокальциферол
- 50. Скачать презентацию
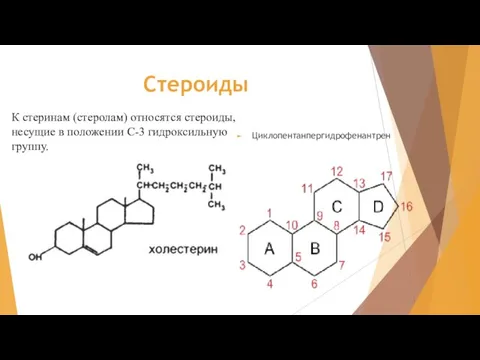
Слайд 2Стероиды
Циклопентанпергидрофенантрен
К стеринам (стеролам) относятся стероиды, несущие в положении С-3 гидроксильную группу.
Стероиды
Циклопентанпергидрофенантрен
К стеринам (стеролам) относятся стероиды, несущие в положении С-3 гидроксильную группу.
Слайд 3Гормоны – это БАВ, выделяемые железами внутренней секреции и являющиеся регуляторами обмена
Гормоны – это БАВ, выделяемые железами внутренней секреции и являющиеся регуляторами обмена

Слайд 4Холестерин (класс зоостеринов), С27Н46О. Обнаруживается почти во всех органах и тканях животных
Холестерин (класс зоостеринов), С27Н46О. Обнаруживается почти во всех органах и тканях животных

Слайд 5Стерины растений (фитостерины)
Эргостерин по структуре отличается от холестерина дополнительной метильной группой в
Стерины растений (фитостерины)
Эргостерин по структуре отличается от холестерина дополнительной метильной группой в

Слайд 6Стигмастерин С29H48О содержится в большом количестве в соевом масле и сахарном тростнике.
Стигмастерин С29H48О содержится в большом количестве в соевом масле и сахарном тростнике.

Слайд 7Ситостерин С29Н50О. В хлопковом и соевом маслах, в зародышах пшеницы и натуральном
Ситостерин С29Н50О. В хлопковом и соевом маслах, в зародышах пшеницы и натуральном

Слайд 8В основе истории синтеза стероидных гормонов лежат методы биотрансформации, результатом применения которых
В основе истории синтеза стероидных гормонов лежат методы биотрансформации, результатом применения которых

Слайд 9Применение микроорганизмов в качестве носителей активных полиферментных систем, способных переводить экзогенные органические
Применение микроорганизмов в качестве носителей активных полиферментных систем, способных переводить экзогенные органические

Слайд 10Микробиологическая трансформация – использование ферментативной активности жизнеспособных клеток микроорганизмов, результатом чего является
Микробиологическая трансформация – использование ферментативной активности жизнеспособных клеток микроорганизмов, результатом чего является

Слайд 11Еще в конце XIX в. было известно, что бактериальная флора кишечника млекопитающих превращает
Еще в конце XIX в. было известно, что бактериальная флора кишечника млекопитающих превращает

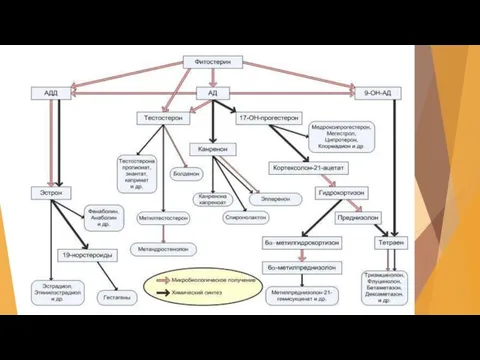
Слайд 12В настоящее время принята классификация микробиологических трансформаций по типу возникновения и отщепления
В настоящее время принята классификация микробиологических трансформаций по типу возникновения и отщепления

Слайд 14Микробиологическое гидроксилирование
Наличие гидроксильных групп в 3, 11, 16, 17 положениях молекулы стероида,
Микробиологическое гидроксилирование
Наличие гидроксильных групп в 3, 11, 16, 17 положениях молекулы стероида,

Слайд 15Наличие в молекуле стероидов 11-гидроксильной группы обусловливает физиологическую активность гидрокортизона (кортизола) и
Наличие в молекуле стероидов 11-гидроксильной группы обусловливает физиологическую активность гидрокортизона (кортизола) и

Слайд 16Дегидрогенизация стероидов
Преднизолон из кортизона и гидрокортизона, дианабол из метилтестостерона.
Чаще всего микроорганизмы
Дегидрогенизация стероидов
Преднизолон из кортизона и гидрокортизона, дианабол из метилтестостерона.
Чаще всего микроорганизмы

Слайд 17Микробиологическое восстановление
Этот процесс используется в меньшей степени, чем дегидрирование. Он осуществляется главным
Микробиологическое восстановление
Этот процесс используется в меньшей степени, чем дегидрирование. Он осуществляется главным

Слайд 18Окисление гидроксильной группы в кетогруппу
Одна из наиболее частых реакций, осуществляемых микроорганизмами (бактериями,
Окисление гидроксильной группы в кетогруппу
Одна из наиболее частых реакций, осуществляемых микроорганизмами (бактериями,

Слайд 19Гидролиз эфиров стероидов
Открыт в 1938 г. Практическая ценность этой реакции определяется тем, что
Гидролиз эфиров стероидов
Открыт в 1938 г. Практическая ценность этой реакции определяется тем, что

Слайд 20Дезацилирующая способность часто встречается среди микоформ, мукоровых и несовершенных грибов, актиномицетов. Особенность
Дезацилирующая способность часто встречается среди микоформ, мукоровых и несовершенных грибов, актиномицетов. Особенность

Слайд 21Отщепление боковых цепей стероидов
Проблема расщепления боковой цепи стеринов с сохранением стероидного скелета
Отщепление боковых цепей стероидов
Проблема расщепления боковой цепи стеринов с сохранением стероидного скелета

Слайд 22Технологические особенности получения стероидных гормонов
Преимущества иммобилизации
Особенности подачи субстрата
Получение целевого продукта
Основные пути усовершенствования
Технологические особенности получения стероидных гормонов
Преимущества иммобилизации
Особенности подачи субстрата
Получение целевого продукта
Основные пути усовершенствования

Слайд 23Получение гидрокортизона (кортизола) из вещества S
1. Выращивание трансформирующей культуры (I стадия) производят
Получение гидрокортизона (кортизола) из вещества S
1. Выращивание трансформирующей культуры (I стадия) производят

Слайд 242. Трансформация вещества S
также начинается со стерилизации ферментера и воздушного фильтра водным
2. Трансформация вещества S
также начинается со стерилизации ферментера и воздушного фильтра водным

Слайд 253. Выделение гидрокортизона.
Культуральная жидкость вместе с мицелием поступает на сепарацию. Отделенный
3. Выделение гидрокортизона.
Культуральная жидкость вместе с мицелием поступает на сепарацию. Отделенный

Слайд 26Витамины
Жизненно важные низкомолекулярные органические соединения, необходимые для человека в ничтожных количествах, но
Витамины
Жизненно важные низкомолекулярные органические соединения, необходимые для человека в ничтожных количествах, но

Слайд 27История открытия
В сентябре 1880 г. при защите своей докторской диссертации Лунин утверждал, что
История открытия
В сентябре 1880 г. при защите своей докторской диссертации Лунин утверждал, что

Слайд 28В 1906 году Фредерик Хопкинс предположил, что помимо белков, жиров, углеводов и
В 1906 году Фредерик Хопкинс предположил, что помимо белков, жиров, углеводов и

Слайд 29Функции и свойства витаминов:
1. Биостимуляция ферментативных процессов
2. Составная часть коферментов (не всегда)
3. Антимутагенная
4. Поступают в организм
Функции и свойства витаминов:
1. Биостимуляция ферментативных процессов
2. Составная часть коферментов (не всегда)
3. Антимутагенная
4. Поступают в организм

Слайд 30Поскольку бактерии для своего роста и размножения нуждаются в присутствии многих витаминов
Поскольку бактерии для своего роста и размножения нуждаются в присутствии многих витаминов

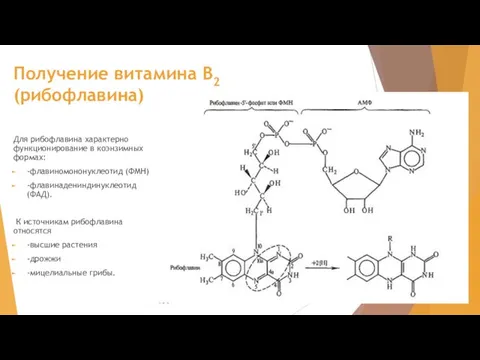
Слайд 31Получение витамина В2 (рибофлавина)
Для рибофлавина характерно функционирование в коэнзимных формах:
-флавиномононуклеотид (ФМН)
Получение витамина В2 (рибофлавина)
Для рибофлавина характерно функционирование в коэнзимных формах:
-флавиномононуклеотид (ФМН)
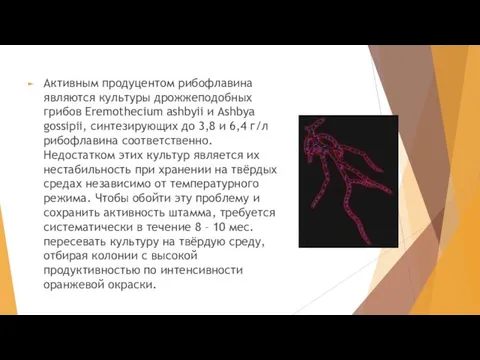
Слайд 32Активным продуцентом рибофлавина являются культуры дрожжеподобных грибов Eremothecium ashbyii и Ashbya gossipii,
Активным продуцентом рибофлавина являются культуры дрожжеподобных грибов Eremothecium ashbyii и Ashbya gossipii,
Слайд 33После выращивания в инокуляторе в течение 21 – 26 часов культуру переводят
После выращивания в инокуляторе в течение 21 – 26 часов культуру переводят

Слайд 34Именно на примере выделения рибофлавина в культуральную жидкость было открыто явление сверхсинтеза.
Именно на примере выделения рибофлавина в культуральную жидкость было открыто явление сверхсинтеза.

Слайд 35Bacillus subtilis
В 1983 году в институте генетики был сконструирован рекомбинантный штамм продуцента
Bacillus subtilis
В 1983 году в институте генетики был сконструирован рекомбинантный штамм продуцента

Слайд 36Меласса (чёрная патока, фр. Mélasse)
Кормовая патока, побочный продукт сахарного производства; сиропообразная жидкость
Меласса (чёрная патока, фр. Mélasse)
Кормовая патока, побочный продукт сахарного производства; сиропообразная жидкость

Слайд 37Розеофлавин
8-диметиламино(нор)-рибофлавин – антибиотик, образуемый Streptomyces davawensis. Показано, что розеофлавин ингибирует синтез 6,7-диметил-8-
Розеофлавин
8-диметиламино(нор)-рибофлавин – антибиотик, образуемый Streptomyces davawensis. Показано, что розеофлавин ингибирует синтез 6,7-диметил-8-


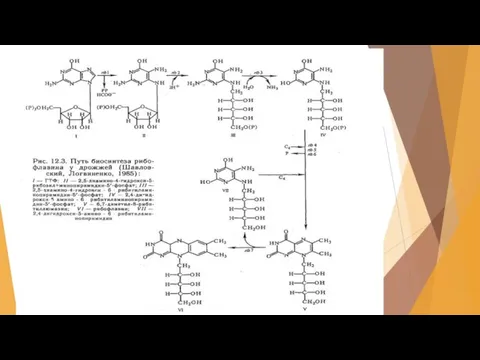

Слайд 41Получение витамина В12
Этот витамин был открыт одновременно в США и в Англии.
Получение витамина В12
Этот витамин был открыт одновременно в США и в Англии.

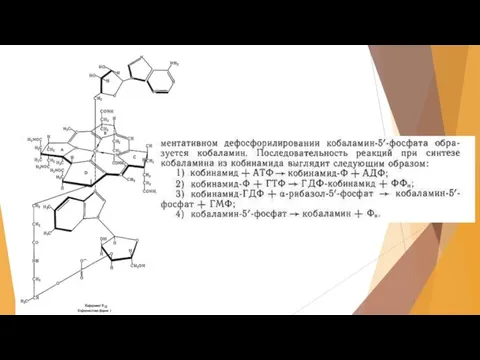
Слайд 42Продуцентом витамина В12 являются пропионовокислые бактерии из рода Propionibacterium. Продуктивность – более
Продуцентом витамина В12 являются пропионовокислые бактерии из рода Propionibacterium. Продуктивность – более

Слайд 43По окончании ферментации витамин остаётся в клетках, поэтому полученную массу сепарируют, стабилизируют
По окончании ферментации витамин остаётся в клетках, поэтому полученную массу сепарируют, стабилизируют


Слайд 46Витамин С
Одним из классических примеров промышленного использования микроорганизмов в получении витамина С
Витамин С
Одним из классических примеров промышленного использования микроорганизмов в получении витамина С

Слайд 47Наиболее сильной окислительной способностью обладают штаммы Acetobacter melanogenum и Gluconobacter oxydans.
Для
Наиболее сильной окислительной способностью обладают штаммы Acetobacter melanogenum и Gluconobacter oxydans.
Для

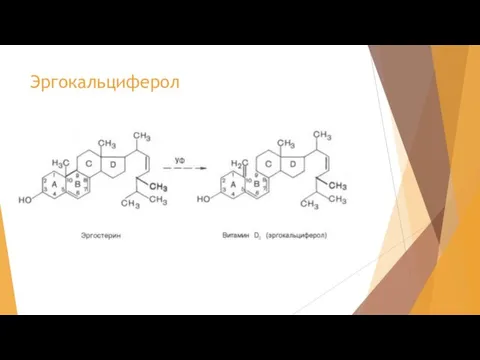
Слайд 48Эргокальциферол
Эргокальциферол
 Молодежный вид спорта SOCKS
Молодежный вид спорта SOCKS Золотые ворота в Древнюю Русь
Золотые ворота в Древнюю Русь Медициналық сақтандыру
Медициналық сақтандыру Электрические системы управления входными устройствами воздухозаборников реактивных двигателей
Электрические системы управления входными устройствами воздухозаборников реактивных двигателей Признаки права. Субъективное и объективное в праве
Признаки права. Субъективное и объективное в праве Крещенский вечерок
Крещенский вечерок ЗемледельцыАттики
ЗемледельцыАттики 15914562539417276895edbb1fde1816-Короткова В., школа 1 (2)
15914562539417276895edbb1fde1816-Короткова В., школа 1 (2) Функции даты и времени
Функции даты и времени Защита электродвигателейсредней и большой мощности
Защита электродвигателейсредней и большой мощности МБОУ Балахтинская средняя общеобразовательная школа № 1
МБОУ Балахтинская средняя общеобразовательная школа № 1  гипотеза стационарного состояния
гипотеза стационарного состояния 9 класс
9 класс Презентация на тему Норманны. Завоевание Англии
Презентация на тему Норманны. Завоевание Англии  Игра «Крестики - нолики»
Игра «Крестики - нолики» Жилище юрта
Жилище юрта Оптимизация фондовых портфелей с учетом нечетко-множественных описаний
Оптимизация фондовых портфелей с учетом нечетко-множественных описаний Булочка с творогом
Булочка с творогом Рекомендации для родителей по обучению детей ПДД
Рекомендации для родителей по обучению детей ПДД Анна Бузунко. Мастер техники лэмпворк
Анна Бузунко. Мастер техники лэмпворк Итоговое сочинение 2021-2022.метапредметность
Итоговое сочинение 2021-2022.метапредметность Основы безопасности: сетевые коммутаторы RTT
Основы безопасности: сетевые коммутаторы RTT Футбол. История футбола
Футбол. История футбола 共多少钱
共多少钱 Презентация на тему Доли Обыкновенные дроби
Презентация на тему Доли Обыкновенные дроби  Замок Перелада
Замок Перелада Использование лучших мировых практик управления промышленными корпорациями
Использование лучших мировых практик управления промышленными корпорациями Разработка и внедрение системы точного позиционирования рольганг - телег ЛПЦ-4 средствами АСУ ТП ЛЗС №2
Разработка и внедрение системы точного позиционирования рольганг - телег ЛПЦ-4 средствами АСУ ТП ЛЗС №2