- ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЖИЗНИ НА ЗЕМЛЕ

Содержание
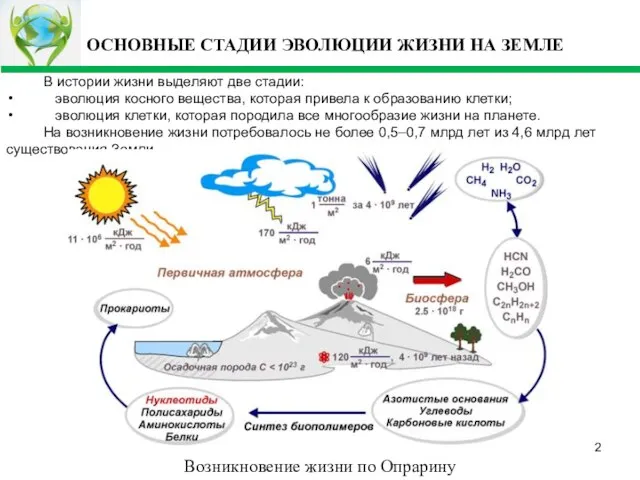
- 2. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ НА ЗЕМЛЕ В истории жизни выделяют две стадии: эволюция косного вещества, которая
- 3. ЭВОЛЮЦИЯ КОСНОГО ВЕЩЕСТВА Основные этапы эволюции Состав первичной атмосферы: 60% - CO2, 40% - аммиак, хлористый
- 4. Автокаталический синтез органических соединений известен - это синтез сахаров из формальдегида. Он протекает в присутствии ионов
- 5. ЕСТЕСТВЕННЫЙ ОТБОР И АВТОКАТАЛИЧЕСКИЕ РЕАКЦИИ Пусть R >Qкрит, где Qкрит - критический уровень пищи, при котором
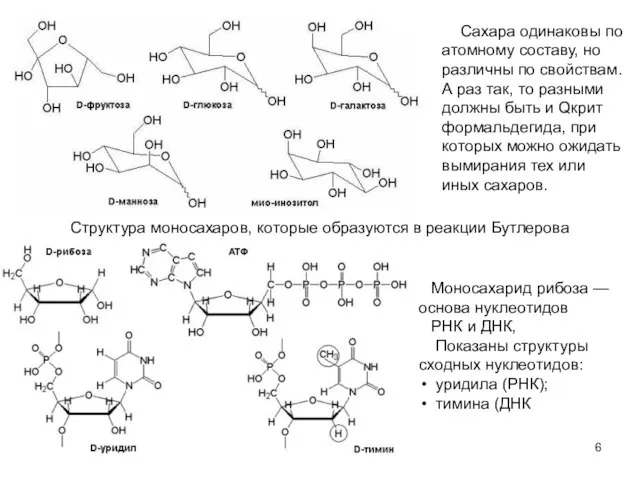
- 6. Структура моносахаров, которые образуются в реакции Бутлерова Моносахарид рибоза — основа нуклеотидов РНК и ДНК, Показаны
- 7. ВЫВОДЫ Естественный отбор начинался простых соединений, которые являются основой РНК и ДНК. Первичный бульон мог быть
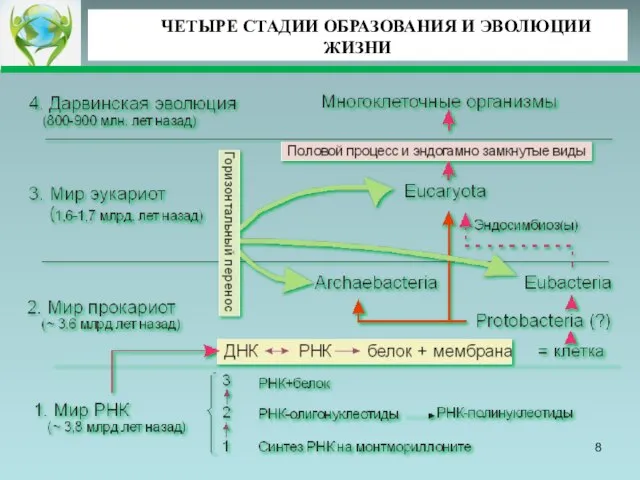
- 8. ЧЕТЫРЕ СТАДИИ ОБРАЗОВАНИЯ И ЭВОЛЮЦИИ ЖИЗНИ
- 9. СЦЕНАРИЙ ОБРАЗОВАНИЯ И РАЗВИТИЯ ЖИЗНИ Стадия 1. Мир РНК Экспериментально показано, что РНК могла синтезироваться на
- 10. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ Стадия 2. Мир прокариот Существуют доказательства, что вначале появились одноклеточные без ядра
- 11. Биогеохимические циклы прокариот Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические связи должны быть
- 12. Биогеохимические циклы прокариот Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические связи должны быть
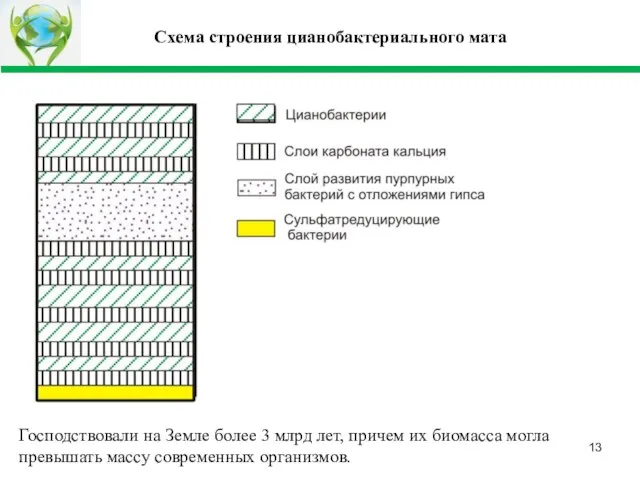
- 13. Схема строения цианобактериального мата Господствовали на Земле более 3 млрд лет, причем их биомасса могла превышать
- 14. Изменение разнообразия строматолитов
- 15. Пояснение к слайду Цианобактериальные системы имели несколько максимумов: Первый максимум, около 2 назад млрд. лет, соответствовал
- 16. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ Стадия 3. Эукариоты III стадия ~1,7 млрд лет назад. Появление эукариот связывают
- 17. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ Точка Пастера Критические для эволюции биосферы содержания кислорода в атмосфере получили название
- 18. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ Временные этапы мнение палеонтологов... Если наиболее древние нитчатые микрофоссилии, напоминающие ныне живущих
- 19. ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ Этапы формирования Биосферы Зарождение гетеротрофных форм жизни. Возникновение автотрофных форм, реализующих хемосинтез.
- 20. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ усложнение форм жизни; аддитивный характер; неравномерность развития. Усложнение форм жизни Очевидно, что
- 21. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ Макросистема биоты и способы метаболизма
- 22. Итак, эволюция простейших была направлена на развитие всевозможных формы метаболизма, что позволяло использовать различные источники питания
- 23. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ Пути эволюции на разных этапах становления биосферы
- 24. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ
- 25. ОСНОВНЫЕ ПУТИ ЭВОЛЮЦИИ НА РАЗНЫХ ЭТАПАХ На ранних этапах эволюции усложнение затрагивало в основном биохимию. Появлялись
- 26. Рост устойчивости и приспособляемости живых систем
- 27. РОСТ УСТОЙЧИВОСТИ И ПРИСПОСОБЛЯЕМОСТИ ЖИВЫХ СИСТЕМ Эволюционные линии, дожившие до наших дней, испытали множество кризисов, катастроф
- 28. РОСТ БИОРАЗНООБРАЗИЯ Основные закономерности изменения глобального таксономического разнообразия на примере морской биоты в фанерозое следующие (см.
- 29. ГЛОБАЛЬНЫЕ КРИЗИСЫ Временные падения разнообразия биоты в биосферы получили название массовых вымираний. Кризисы могли иметь не
- 30. ГЛОБАЛЬНЫЕ КРИЗИСЫ В ИСТОРИИ ЗЕМЛИ 1. Раннекембрийская радиация. Появление и быстрое развитие организмов, имеющих скелет. Это
- 31. НЕРАВНОМЕРНОСТЬ РАЗВИТИЯ. РОЛЬ КРИЗИСОВ В ЭВОЛЮЦИИ Вернадский В.И. определил важнейшие биогеохимические свойства жизни как «растекание» и
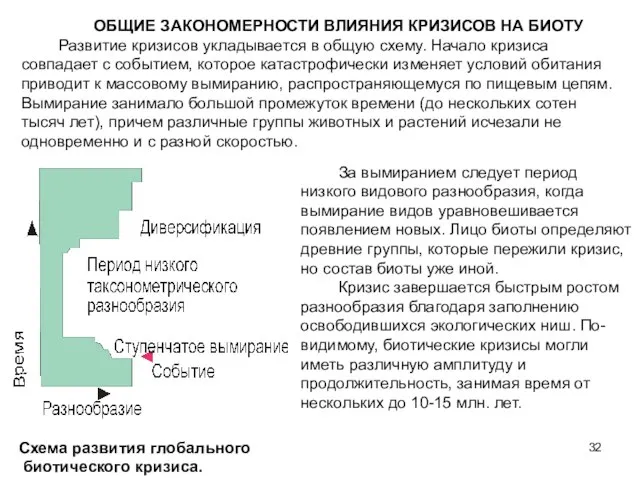
- 32. ОБЩИЕ ЗАКОНОМЕРНОСТИ ВЛИЯНИЯ КРИЗИСОВ НА БИОТУ Развитие кризисов укладывается в общую схему. Начало кризиса совпадает с
- 33. ОСОБЕННОСТИ МАССОВЫХ ВЫМИРАНИЙ Известно, что некоторые вымирания (например, исчезновение 65 млн. лет назад динозавров, крылатых ящеров
- 35. Различия направлений эволюции neanderthalensis (Н) и sapiens (С) Линии (Н) и (С) разошлись 500-600 тыс. лет
- 37. Скачать презентацию
Слайд 3ЭВОЛЮЦИЯ КОСНОГО ВЕЩЕСТВА
Основные этапы эволюции
Состав первичной атмосферы: 60% - CO2, 40%
ЭВОЛЮЦИЯ КОСНОГО ВЕЩЕСТВА
Основные этапы эволюции
Состав первичной атмосферы: 60% - CO2, 40%

Слайд 4 Автокаталический синтез органических соединений известен - это синтез сахаров из формальдегида. Он
Автокаталический синтез органических соединений известен - это синтез сахаров из формальдегида. Он

Слайд 5 ЕСТЕСТВЕННЫЙ ОТБОР И
АВТОКАТАЛИЧЕСКИЕ РЕАКЦИИ
Пусть R >Qкрит, где Qкрит -
ЕСТЕСТВЕННЫЙ ОТБОР И
АВТОКАТАЛИЧЕСКИЕ РЕАКЦИИ
Пусть R >Qкрит, где Qкрит -

Слайд 6Структура моносахаров, которые образуются в реакции Бутлерова
Моносахарид рибоза — основа нуклеотидов
РНК и
Структура моносахаров, которые образуются в реакции Бутлерова
Моносахарид рибоза — основа нуклеотидов
РНК и
Слайд 7ВЫВОДЫ
Естественный отбор начинался простых соединений, которые являются основой РНК и ДНК.
Первичный
ВЫВОДЫ
Естественный отбор начинался простых соединений, которые являются основой РНК и ДНК.
Первичный

Слайд 8ЧЕТЫРЕ СТАДИИ ОБРАЗОВАНИЯ И ЭВОЛЮЦИИ ЖИЗНИ
ЧЕТЫРЕ СТАДИИ ОБРАЗОВАНИЯ И ЭВОЛЮЦИИ ЖИЗНИ
Слайд 9СЦЕНАРИЙ ОБРАЗОВАНИЯ И РАЗВИТИЯ ЖИЗНИ
Стадия 1. Мир РНК
Экспериментально показано, что РНК могла
СЦЕНАРИЙ ОБРАЗОВАНИЯ И РАЗВИТИЯ ЖИЗНИ
Стадия 1. Мир РНК
Экспериментально показано, что РНК могла

Слайд 10ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Стадия 2. Мир прокариот
Существуют доказательства, что вначале появились
ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Стадия 2. Мир прокариот
Существуют доказательства, что вначале появились

Слайд 11Биогеохимические циклы прокариот
Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические
Биогеохимические циклы прокариот
Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические

Слайд 12Биогеохимические циклы прокариот
Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические
Биогеохимические циклы прокариот
Биогеохимические циклы – важное условие устойчивости экосистемы. В цикле трофические

Слайд 13Схема строения цианобактериального мата
Господствовали на Земле более 3 млрд лет, причем их
Схема строения цианобактериального мата
Господствовали на Земле более 3 млрд лет, причем их
Слайд 14Изменение разнообразия строматолитов
Изменение разнообразия строматолитов
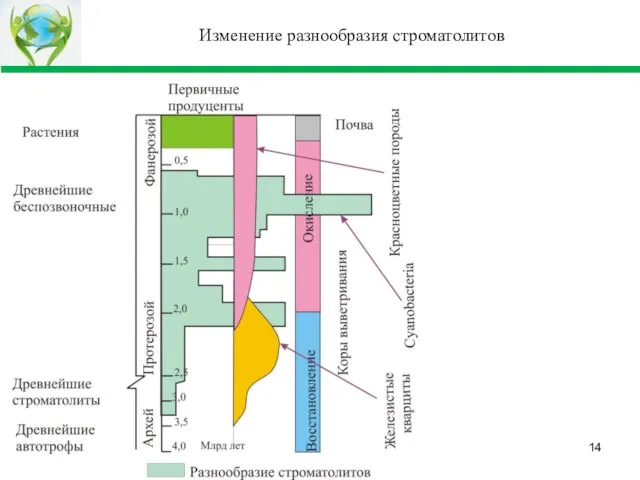
Слайд 15Пояснение к слайду
Цианобактериальные системы имели несколько максимумов:
Первый максимум, около 2 назад млрд.
Пояснение к слайду
Цианобактериальные системы имели несколько максимумов:
Первый максимум, около 2 назад млрд.

Слайд 16ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Стадия 3. Эукариоты
III стадия ~1,7 млрд лет назад.
ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Стадия 3. Эукариоты
III стадия ~1,7 млрд лет назад.

Слайд 17ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Точка Пастера
Критические для эволюции биосферы содержания кислорода в атмосфере
ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Точка Пастера
Критические для эволюции биосферы содержания кислорода в атмосфере

Слайд 18ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Временные этапы
мнение палеонтологов...
Если наиболее древние нитчатые микрофоссилии, напоминающие
ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Временные этапы
мнение палеонтологов...
Если наиболее древние нитчатые микрофоссилии, напоминающие

Слайд 19ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Этапы формирования Биосферы
Зарождение гетеротрофных форм жизни.
Возникновение
ОСНОВНЫЕ СТАДИИ ЭВОЛЮЦИИ ЖИЗНИ
Этапы формирования Биосферы
Зарождение гетеротрофных форм жизни.
Возникновение

Слайд 20ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ
усложнение форм жизни;
аддитивный характер;
неравномерность развития.
Усложнение форм
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ
усложнение форм жизни;
аддитивный характер;
неравномерность развития.
Усложнение форм

Слайд 21ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ Макросистема биоты и способы метаболизма
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ Макросистема биоты и способы метаболизма
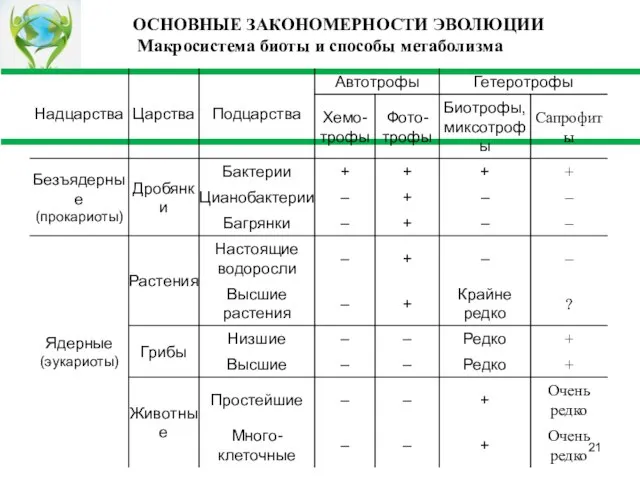
Слайд 22 Итак, эволюция простейших была направлена на развитие всевозможных формы метаболизма, что позволяло
Итак, эволюция простейших была направлена на развитие всевозможных формы метаболизма, что позволяло

Слайд 23ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Пути эволюции на разных этапах становления биосферы
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Пути эволюции на разных этапах становления биосферы

Слайд 24ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ
ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ ЖИЗНИ

Слайд 25ОСНОВНЫЕ ПУТИ ЭВОЛЮЦИИ НА РАЗНЫХ ЭТАПАХ
На ранних этапах эволюции усложнение затрагивало
ОСНОВНЫЕ ПУТИ ЭВОЛЮЦИИ НА РАЗНЫХ ЭТАПАХ
На ранних этапах эволюции усложнение затрагивало

Слайд 26Рост устойчивости и приспособляемости живых систем
Рост устойчивости и приспособляемости живых систем

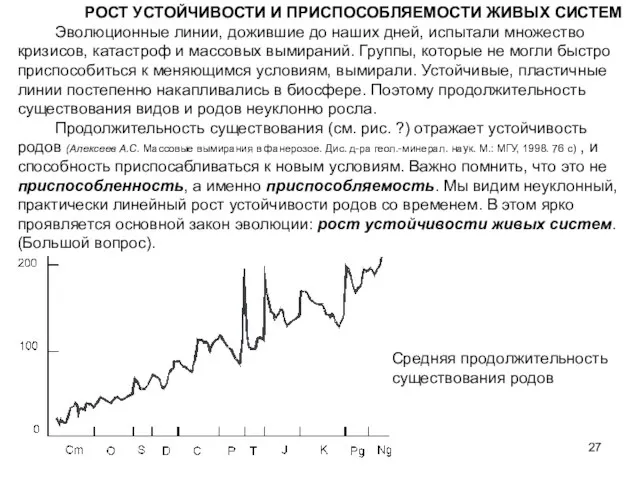
Слайд 27РОСТ УСТОЙЧИВОСТИ И ПРИСПОСОБЛЯЕМОСТИ ЖИВЫХ СИСТЕМ
Эволюционные линии, дожившие до наших дней,
РОСТ УСТОЙЧИВОСТИ И ПРИСПОСОБЛЯЕМОСТИ ЖИВЫХ СИСТЕМ
Эволюционные линии, дожившие до наших дней,
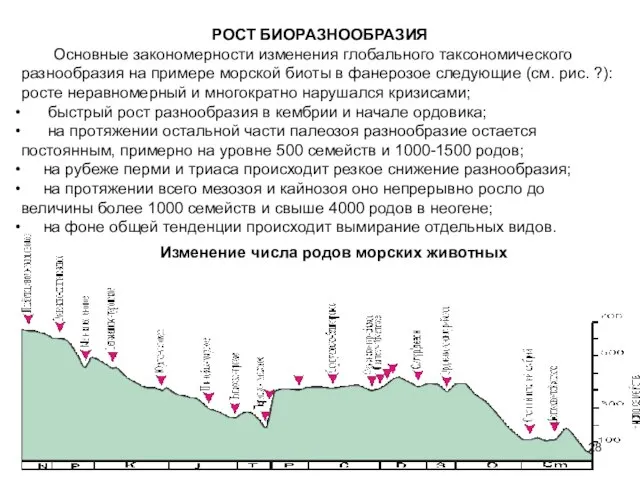
Слайд 28РОСТ БИОРАЗНООБРАЗИЯ
Основные закономерности изменения глобального таксономического разнообразия на примере морской биоты в
РОСТ БИОРАЗНООБРАЗИЯ
Основные закономерности изменения глобального таксономического разнообразия на примере морской биоты в
Слайд 29ГЛОБАЛЬНЫЕ КРИЗИСЫ
Временные падения разнообразия биоты в биосферы получили название массовых вымираний.
ГЛОБАЛЬНЫЕ КРИЗИСЫ
Временные падения разнообразия биоты в биосферы получили название массовых вымираний.
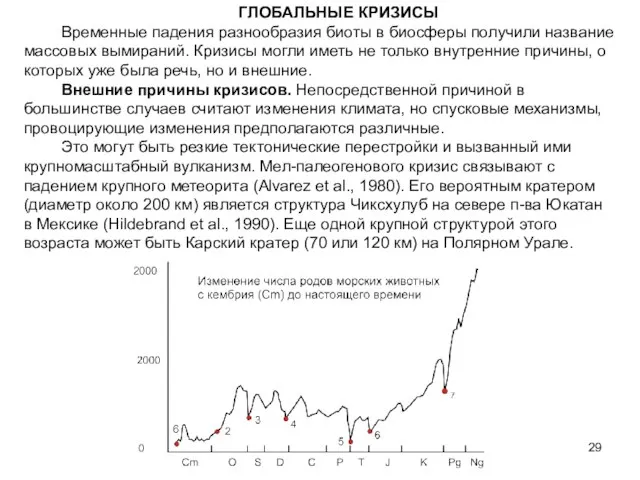
Слайд 30ГЛОБАЛЬНЫЕ КРИЗИСЫ В ИСТОРИИ ЗЕМЛИ
1. Раннекембрийская радиация. Появление и быстрое развитие организмов,
ГЛОБАЛЬНЫЕ КРИЗИСЫ В ИСТОРИИ ЗЕМЛИ
1. Раннекембрийская радиация. Появление и быстрое развитие организмов,

Слайд 31НЕРАВНОМЕРНОСТЬ РАЗВИТИЯ. РОЛЬ КРИЗИСОВ В ЭВОЛЮЦИИ
Вернадский В.И. определил важнейшие биогеохимические свойства жизни
НЕРАВНОМЕРНОСТЬ РАЗВИТИЯ. РОЛЬ КРИЗИСОВ В ЭВОЛЮЦИИ
Вернадский В.И. определил важнейшие биогеохимические свойства жизни

Слайд 32ОБЩИЕ ЗАКОНОМЕРНОСТИ ВЛИЯНИЯ КРИЗИСОВ НА БИОТУ
Развитие кризисов укладывается в общую схему. Начало
ОБЩИЕ ЗАКОНОМЕРНОСТИ ВЛИЯНИЯ КРИЗИСОВ НА БИОТУ
Развитие кризисов укладывается в общую схему. Начало
Слайд 33ОСОБЕННОСТИ МАССОВЫХ ВЫМИРАНИЙ
Известно, что некоторые вымирания (например, исчезновение 65 млн. лет
ОСОБЕННОСТИ МАССОВЫХ ВЫМИРАНИЙ
Известно, что некоторые вымирания (например, исчезновение 65 млн. лет

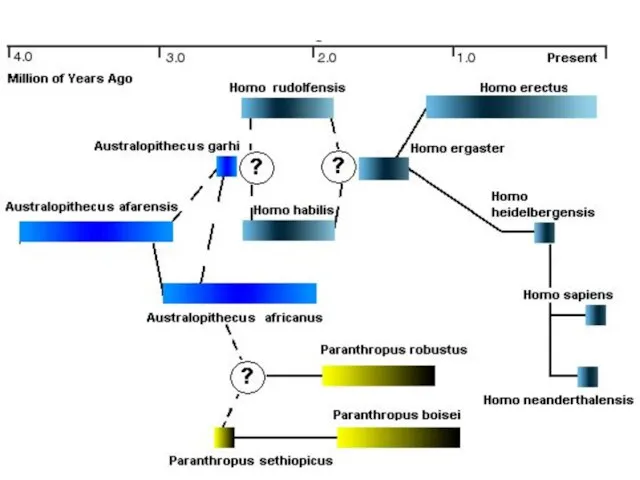
Слайд 35Различия направлений эволюции neanderthalensis (Н) и sapiens (С)
Линии (Н) и (С) разошлись
Различия направлений эволюции neanderthalensis (Н) и sapiens (С)
Линии (Н) и (С) разошлись

 Правила поведения на водоёмах в осенне-зимний период
Правила поведения на водоёмах в осенне-зимний период Уставный капитал 480 млн. рублей ОАО «Камский Индустриальный парк «Мастер» основано 29 июля 2004 года в городе Набережные Челны на базе
Уставный капитал 480 млн. рублей ОАО «Камский Индустриальный парк «Мастер» основано 29 июля 2004 года в городе Набережные Челны на базе Охрана окружающей среды и рациональное использование природных ресурсов
Охрана окружающей среды и рациональное использование природных ресурсов Презентация на тему Духовно-нравственное воспитание в современной школе
Презентация на тему Духовно-нравственное воспитание в современной школе Das ist eine alte deutsche Stadt
Das ist eine alte deutsche Stadt Моя помощь ближнему
Моя помощь ближнему 4 ноября - День народного единства. Историческая справка
4 ноября - День народного единства. Историческая справка Опекаемые и публичные благав экономической теории
Опекаемые и публичные благав экономической теории Семион Полоцкий
Семион Полоцкий Для презентаций
Для презентаций Долина смерти
Долина смерти животный мир
животный мир ПРОБЛЕМНОЕ ОБУЧЕНИЕ(PbL)
ПРОБЛЕМНОЕ ОБУЧЕНИЕ(PbL) В новый год со спортом. Фотопрезентация Железновой Анастасии
В новый год со спортом. Фотопрезентация Железновой Анастасии Организаторы:
Организаторы: Административные механизмы и стимулы в законодательстве об энергосбережении и о повышении энергетической эффективности в Россий
Административные механизмы и стимулы в законодательстве об энергосбережении и о повышении энергетической эффективности в Россий Архитектура вычислительной системы. Классификация компьютеров
Архитектура вычислительной системы. Классификация компьютеров Психология и творчество
Психология и творчество Деловой клуб
Деловой клуб Инвентаризация, её роль в бухгалтерском учёте
Инвентаризация, её роль в бухгалтерском учёте Палагайская школа вчера, сегодня, завтра
Палагайская школа вчера, сегодня, завтра Электронный журнал творческих работ учащихся «Путь к слову»
Электронный журнал творческих работ учащихся «Путь к слову» Личность. Индивид. Индивидуальность. Сознание. Самосознание
Личность. Индивид. Индивидуальность. Сознание. Самосознание В СПб и ЛО
В СПб и ЛО Промышленность Приозерска
Промышленность Приозерска Основания, кислоты, соли
Основания, кислоты, соли Сравнительный анализ конституции и транспортной системы России и Германии
Сравнительный анализ конституции и транспортной системы России и Германии Метод проектов – способ формирования исследовательской деятельности учащихся
Метод проектов – способ формирования исследовательской деятельности учащихся