- Аминокислоты.Транспорт аминокислот. Тема 12

Содержание
- 2. Пути образования и поддержани пула аминокислот в организме. Транспорт аминокислот в клетки. 5. Транспортные белки (транспорт
- 3. Биологические функции аминoкислoт. 1. Стpуктуpные элeменты пeптидов и белков. В состав белков входят 20 протеиногенных аминокислот,
- 4. Пути образования и поддержани пула аминокислот в организме.
- 6. Транспорт аминокислот через клеточные мембраны Различная скорость проникновения аминокислот через мембраны клеток, установленная при помощи метода
- 7. Транспорт аминокислот через клеточные мембраны
- 9. ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ Промежуточный метаболизм аминокислот белковых молекул, как и других питательных веществ в
- 10. ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ Рис. 12.2. Катаболизм аминокислот.
- 11. Общие пути обмена аминокислот Общие пути превращения аминокислот включают реакции деаминирования, трансаминирования, декарбоксилирования, биосинтеза и рацемизации.
- 12. Общие пути обмена аминокислот Деаминирование аминокислот Доказано существование 4 типов деаминирования аминокислот (отщепление аминогруппы). Во всех
- 13. 1. Восстановительное деаминирование 2. Гидролитическое деаминирование
- 14. 3. Внутримолекулярное деаминирование (элиминирование) 4. Окислительное деаминирование
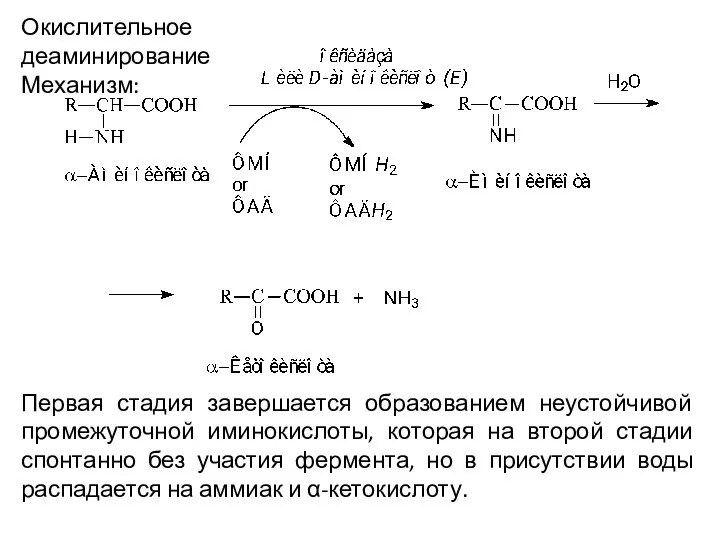
- 15. Окислительное деаминирование Механизм: Первая стадия завершается образованием неустойчивой промежуточной иминокислоты, которая на второй стадии спонтанно без
- 16. Помимо аммиака, продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Для животных тканей, растений и большинства
- 17. Восстановленные флавиновые мононуклеотиды могут быть окислены кислородом (O2). Окислительное деаминирование глутамата
- 18. Схематически реакции окислительного деаминирования аминокислот с участием коферментов могут быть представлены в следующем виде:
- 19. Восстановленные флавин-нуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным кислородом. При этом образуется перекись водорода,
- 20. Впервые в лаборатории Д. Грина из ткани печени и почек крыс была выделена оксидаза, катализирующая деаминирование
- 22. Неокислительное деаминирование Cерина, Треонина и Цистеина.
- 23. Декарбоксилирование аминокислот α-Декарбоксилирование аминокислот ω-Декарбоксилирование аминокислот
- 24. Декарбоксилирование аминокислот сопряженное с трансаминированием. Декарбоксилирование аминокислот сопряженное с конденсацией двух молекул.
- 27. Скачать презентацию
Слайд 2Пути образования и поддержани пула аминокислот в организме. Транспорт аминокислот в клетки.
5.
Пути образования и поддержани пула аминокислот в организме. Транспорт аминокислот в клетки.
5.

Слайд 3Биологические функции аминoкислoт.
1. Стpуктуpные элeменты пeптидов и белков. В состав белков
Биологические функции аминoкислoт.
1. Стpуктуpные элeменты пeптидов и белков. В состав белков

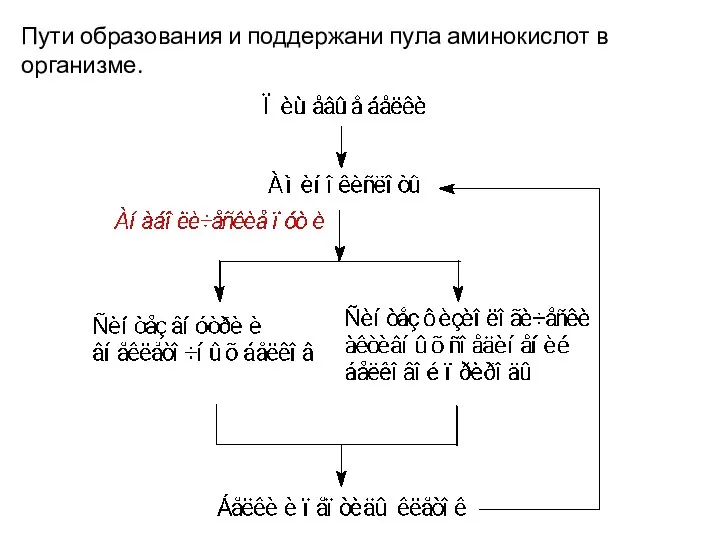
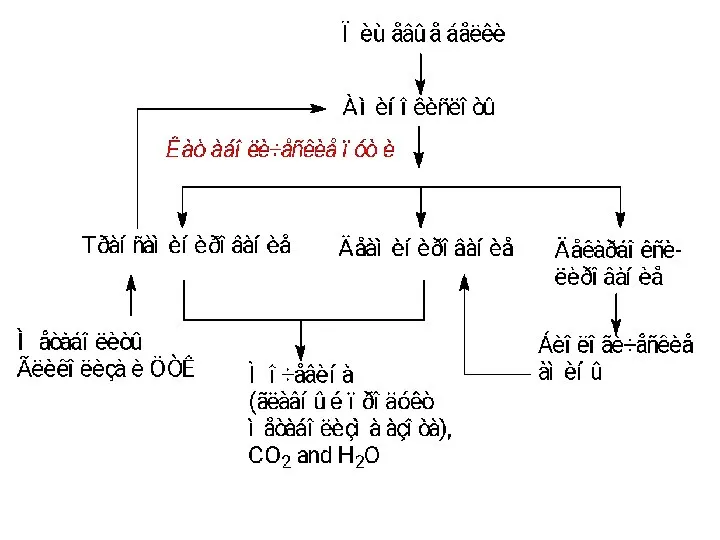
Слайд 4Пути образования и поддержани пула аминокислот в организме.
Пути образования и поддержани пула аминокислот в организме.
Слайд 6Транспорт аминокислот через клеточные мембраны
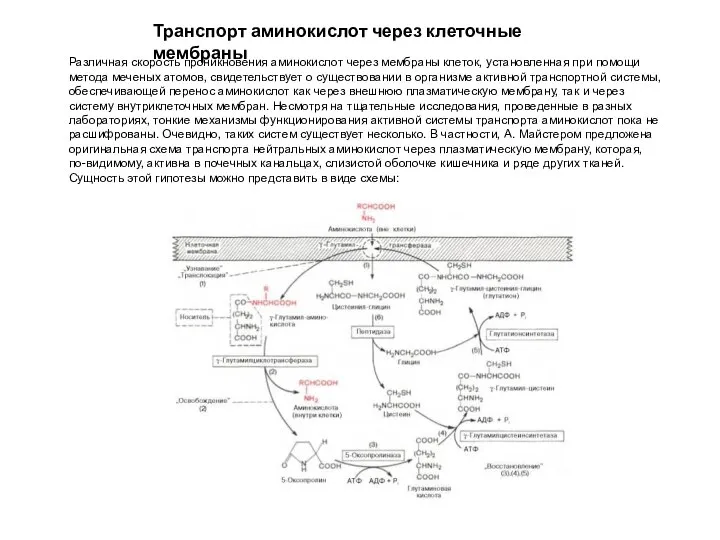
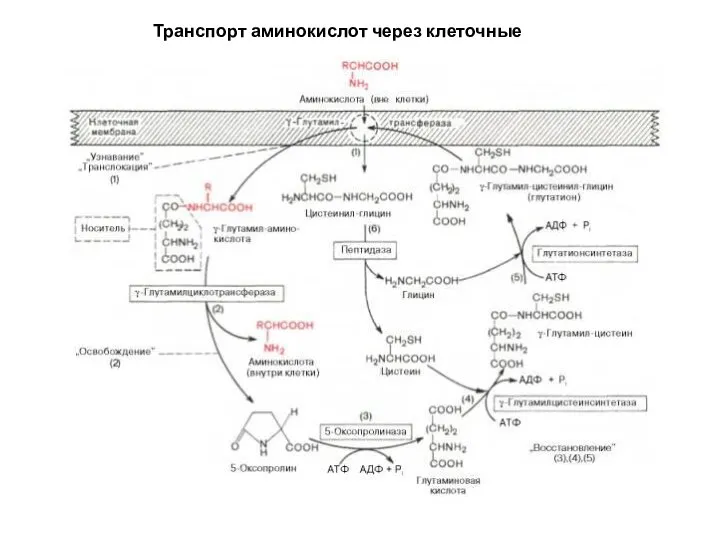
Различная скорость проникновения аминокислот через мембраны клеток, установленная
Транспорт аминокислот через клеточные мембраны
Различная скорость проникновения аминокислот через мембраны клеток, установленная
Слайд 7Транспорт аминокислот через клеточные мембраны
Транспорт аминокислот через клеточные мембраны
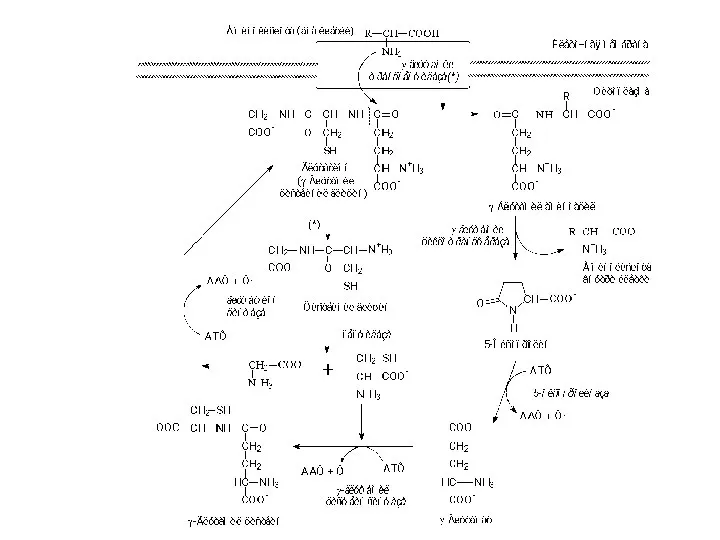
Слайд 9ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ
Промежуточный метаболизм аминокислот белковых молекул, как и других
ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ
Промежуточный метаболизм аминокислот белковых молекул, как и других

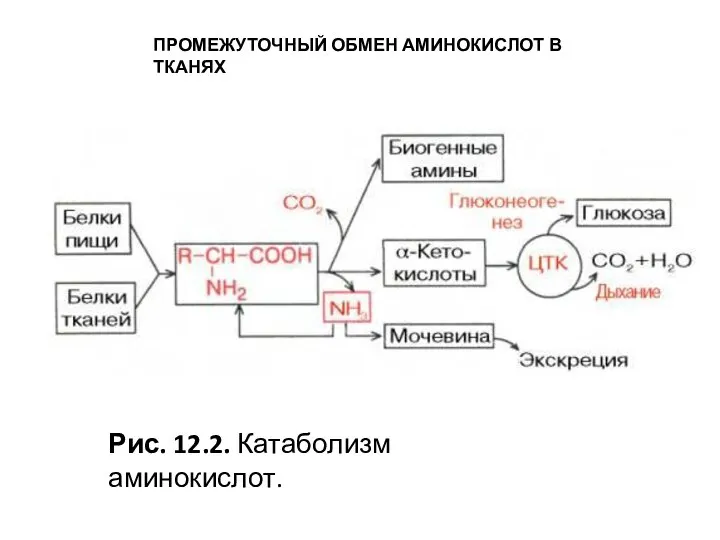
Слайд 10ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ
Рис. 12.2. Катаболизм аминокислот.
ПРОМЕЖУТОЧНЫЙ ОБМЕН АМИНОКИСЛОТ В ТКАНЯХ
Рис. 12.2. Катаболизм аминокислот.
Слайд 11Общие пути обмена аминокислот
Общие пути превращения аминокислот включают реакции
деаминирования,
трансаминирования,
декарбоксилирования,
Общие пути обмена аминокислот
Общие пути превращения аминокислот включают реакции
деаминирования,
трансаминирования,
декарбоксилирования,

Слайд 12Общие пути обмена аминокислот
Деаминирование аминокислот
Доказано существование 4 типов деаминирования аминокислот (отщепление аминогруппы).
Общие пути обмена аминокислот
Деаминирование аминокислот
Доказано существование 4 типов деаминирования аминокислот (отщепление аминогруппы).

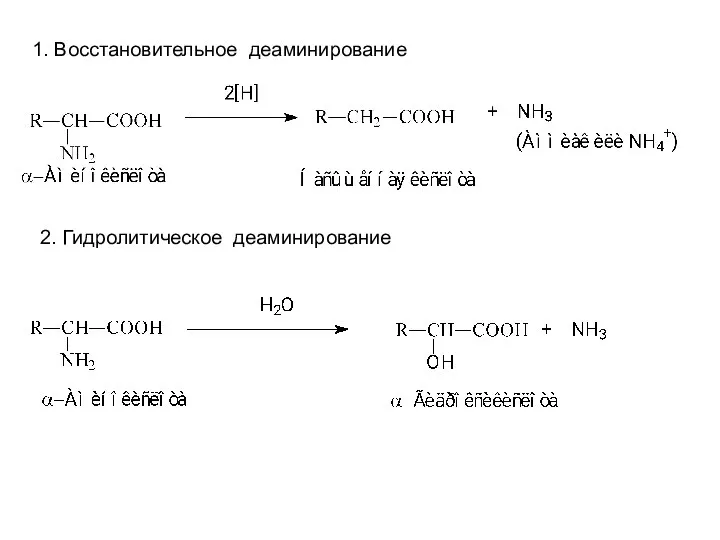
Слайд 131. Восстановительное деаминирование
2. Гидролитическое деаминирование
1. Восстановительное деаминирование
2. Гидролитическое деаминирование
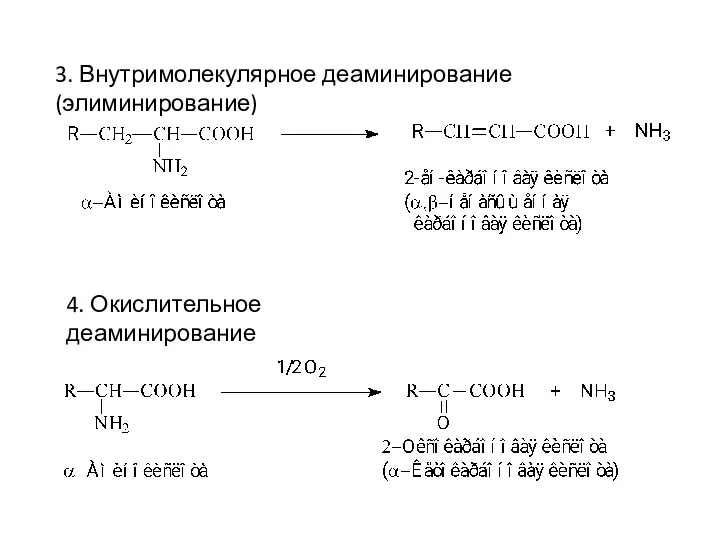
Слайд 143. Внутримолекулярное деаминирование (элиминирование)
4. Окислительное деаминирование
3. Внутримолекулярное деаминирование (элиминирование)
4. Окислительное деаминирование
Слайд 15Окислительное деаминирование
Механизм:
Первая стадия завершается образованием неустойчивой промежуточной иминокислоты, которая на второй стадии
Окислительное деаминирование
Механизм:
Первая стадия завершается образованием неустойчивой промежуточной иминокислоты, которая на второй стадии
Слайд 16Помимо аммиака, продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Для животных
Помимо аммиака, продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Для животных

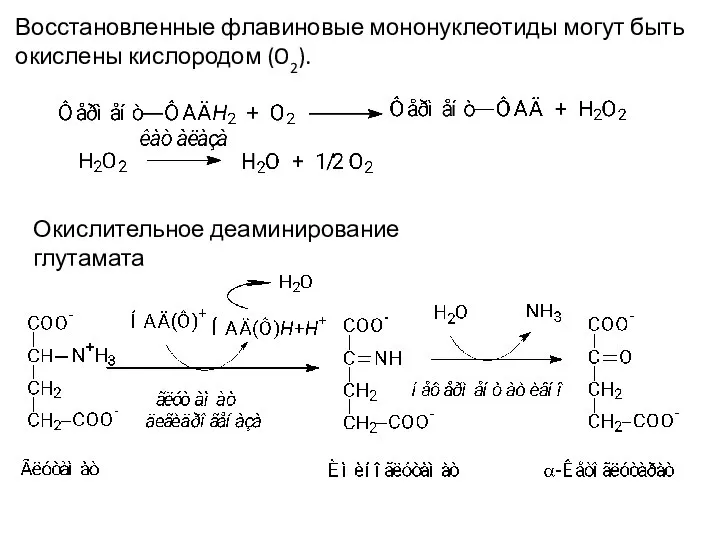
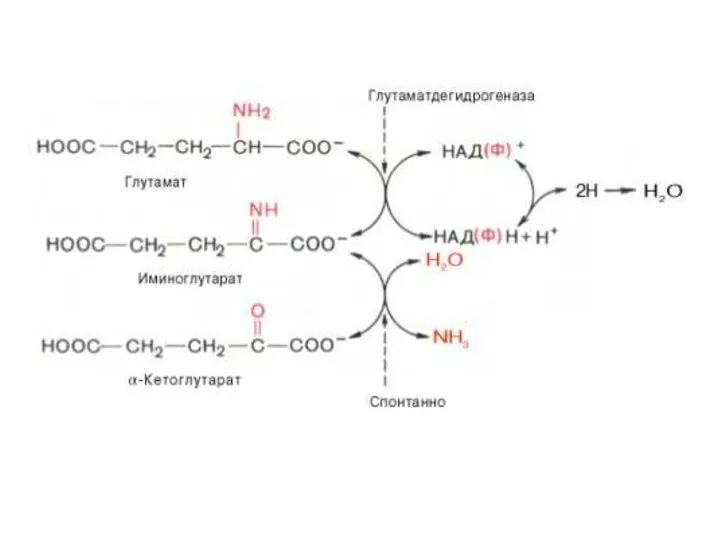
Слайд 17Восстановленные флавиновые мононуклеотиды могут быть окислены кислородом (O2).
Окислительное деаминирование глутамата
Восстановленные флавиновые мононуклеотиды могут быть окислены кислородом (O2).
Окислительное деаминирование глутамата
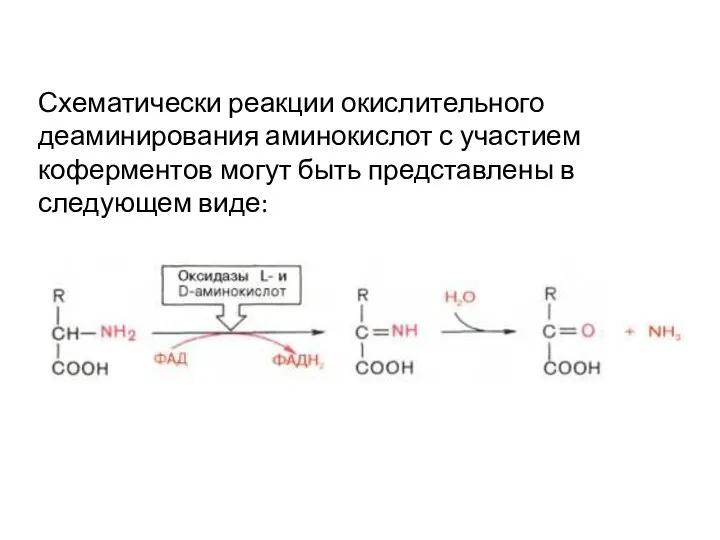
Слайд 18Схематически реакции окислительного деаминирования аминокислот с участием коферментов могут быть представлены в
Схематически реакции окислительного деаминирования аминокислот с участием коферментов могут быть представлены в
Слайд 19Восстановленные флавин-нуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным кислородом. При
Восстановленные флавин-нуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным кислородом. При

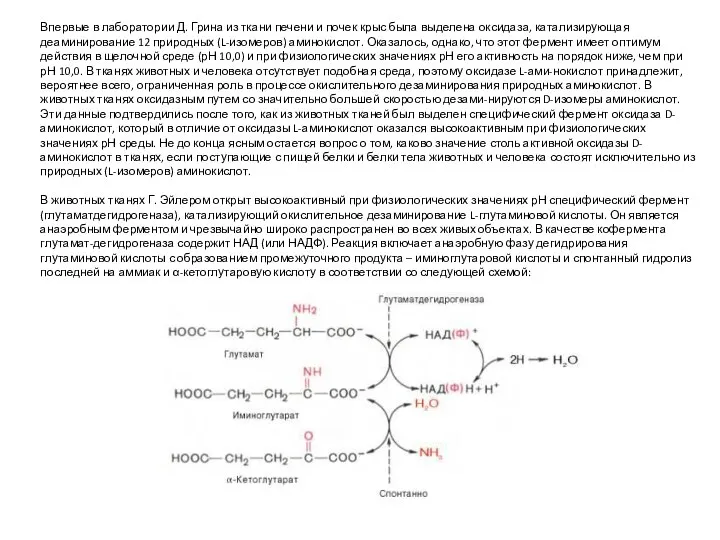
Слайд 20Впервые в лаборатории Д. Грина из ткани печени и почек крыс была
Впервые в лаборатории Д. Грина из ткани печени и почек крыс была
Слайд 22Неокислительное деаминирование Cерина, Треонина и Цистеина.
Неокислительное деаминирование Cерина, Треонина и Цистеина.

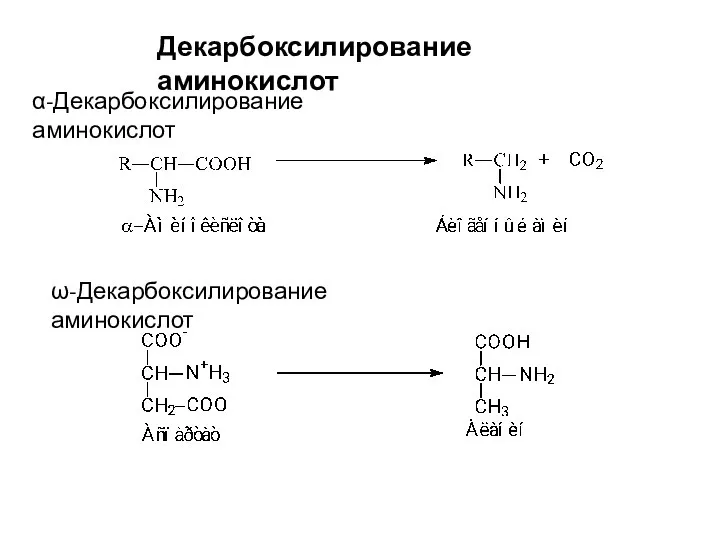
Слайд 23Декарбоксилирование аминокислот
α-Декарбоксилирование аминокислот
ω-Декарбоксилирование аминокислот
Декарбоксилирование аминокислот
α-Декарбоксилирование аминокислот
ω-Декарбоксилирование аминокислот
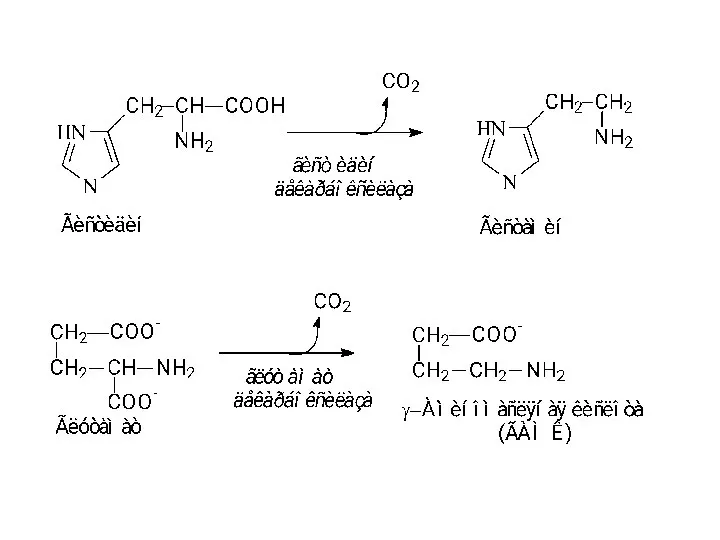
Слайд 24Декарбоксилирование аминокислот сопряженное с трансаминированием.
Декарбоксилирование аминокислот сопряженное с конденсацией двух молекул.
Декарбоксилирование аминокислот сопряженное с трансаминированием.
Декарбоксилирование аминокислот сопряженное с конденсацией двух молекул.

 Росянка. Очаровательный убийца
Росянка. Очаровательный убийца Биологические парки Татарстана
Биологические парки Татарстана Әр түрлі жастағы егеуқұйрықтардың тамақтану рационына асқабақ пен топинамбур талшығын қосу арқылы
Әр түрлі жастағы егеуқұйрықтардың тамақтану рационына асқабақ пен топинамбур талшығын қосу арқылы Регенера́ция (восстановление)
Регенера́ция (восстановление) Низшие. Плесневые грибы
Низшие. Плесневые грибы Жизнь на Земле
Жизнь на Земле Пищеварительная система
Пищеварительная система Презентация на тему РЕЙТИНГ САМЫХ ОПАСНЫХ ЖИВОТНЫХ В МИРЕ
Презентация на тему РЕЙТИНГ САМЫХ ОПАСНЫХ ЖИВОТНЫХ В МИРЕ  Надкласс рыбы
Надкласс рыбы Основные процессы в корнях
Основные процессы в корнях Першо-звіри. Історія появи тварин
Першо-звіри. Історія появи тварин Характеристика и многообразие рыб
Характеристика и многообразие рыб Тип хордовые. Ланцетник
Тип хордовые. Ланцетник Знатоки природы
Знатоки природы Пищеварение
Пищеварение Основы цитологии
Основы цитологии Формы естественного отбора
Формы естественного отбора Vylučovanie - močová sústava človeka
Vylučovanie - močová sústava človeka Годовой жизненный цикл и сезонные явления в жизни птиц
Годовой жизненный цикл и сезонные явления в жизни птиц Горох и его разновидности
Горох и его разновидности Строение животной клетки
Строение животной клетки СКЕЛЕТНЫЕ МЫШЦЫ
СКЕЛЕТНЫЕ МЫШЦЫ Стеблевые суккуленты
Стеблевые суккуленты Презентация на тему История развития жизни
Презентация на тему История развития жизни  Гетерохронность развития
Гетерохронность развития Боль в спине. Истории пациентов (собак)
Боль в спине. Истории пациентов (собак) Сравнительно анатомические доказательства инволюции
Сравнительно анатомические доказательства инволюции Тип Моллюски или Мягкотелые
Тип Моллюски или Мягкотелые