- Азотистый обмен. Общие пути

Содержание
- 2. Азотистый обмен. Общие пути. В организме человека содержится примерно 15 кг белков. Количество свободных аминокислот составляет
- 3. Биологическая роль аминокислот
- 4. Переваривание белков в ЖКТ. Переваривание белков включает гидролиз пищевых белков до свободных аминокислот. Расщепление пептидных связей
- 5. Переваривание белков в ЖКТ
- 6. Переваривание белков в желудке. Переваривание белков в желудке происходит под действием пепсина. Профермент вырабатывается главными клетками
- 7. Переваривание белков в кишечнике Происходит при pH ~ 8,0, изменение pH происходит под действием HCO3-, поступающего
- 8. Переваривание белков в кишечнике
- 9. Трипсин синтезируется в поджелудочной железе в виде профермента трипсиногена. Превращение трипсиногена в трипсин происходит 2 путями:
- 10. Ферменты, переваривающие белки в кишечнике обладают специфичностью к определенным аминокислотам: Трипсин гидролизует пептидные связи между арг
- 11. ЖКТ ТКАНИ КРОВЬ ПЕЧЕНЬ БЕЛКИ АМ. КИСЛОТЫ АМИН-ТЫ АМИНЫ ГЛЮКОЗА Заменимые аминок-ты БЕЛКИ КРОВИ АМИН-ТЫ БЕЛКИ
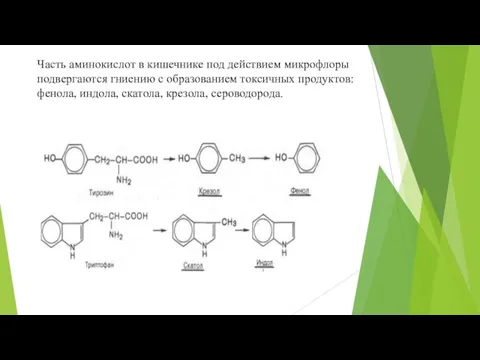
- 12. Часть аминокислот в кишечнике под действием микрофлоры подвергаются гниению с образованием токсичных продуктов: фенола, индола, скатола,
- 13. Общие пути обмена аминокислот Пути распада аминокислот до конечных продуктов можно разделить на три основные группы:
- 14. Основные этапы катаболизма аминокислот
- 15. Превращение α-аминогрупп аминокислот Процесс удаления –аминогрупп называется дезаминированием. Дезаминированию подвергаются все аминокислоты, кроме лизина. Виды дезаминирования
- 16. Виды дезаминирования 1. Окислительное дезаминирование (прямое, характерное для глу) происходит в митохондриях многих тканей, а наиболее
- 17. 2. Прямое неокислительное дезаминирование характерно для серина, треонина, цистеина и гистидина. Серин и треонин дезаминируются с
- 18. Гистидин дезаминируется внутримолекулярным способом Цистеин дезаминируется с выделением H2S c использованием воды.
- 19. Большинство аминокислот в клетках непрямому дезаминированию, которое включает 2 стадии: Трансаминирование с α-кетоглутаратом, образование в цитозоле
- 20. Схема непрямого дезаминирования аминокислот
- 21. Аминотрансферазы, катализирующие реакцию трансаминирования обладают субстратной специфичностью. Наиболее важными являются АСТ и АЛТ. АЛТ катализирует реакцию
- 22. В результате трансаминирования образуется глутаминовая кислота. Затем глутаминовая кислота подвергается окислительному дезаминированию:
- 23. Клиническое значение определения активности аминотрансфераз В сыворотке крови здоровых людей активность аминотрансфераз ниже, чем в органах.
- 24. Декарбоксилирование аминокислот. Процесс отщепления карбоксильной группы в виде СО2 называется декарбоксилированием и приводит к образованию биогенных
- 25. При декарбоксилировании Три, Гис, Фен, Глу образуются биогенные амины: Серотонин обладает сосудосуживающим действием, участвует в регуляции
- 26. Недостаточность дофамина в черной субстанции мозга приводит к болезни Паркинсона. При этом снижена активность ДОФАдекарбоксилазы, тирозингидроксилазы.
- 27. Гистамин обладает сосудорасширяющим действием. Много образуется в области воспаления, участвует в развитии аллергических реакций. ГАМК является
- 28. Орнитин, декарбоксилируясь, дает диамин путресцин (1,4-диаминобутан): При декарбоксилировании лизина образуется кадаверин (1,5-диаминопентан):
- 29. Превращение углеродного скелета аминокислот Катаболизм всех аминокислот сводится к образованию 6 веществ, вступающих в общий путь
- 31. Скачать презентацию
Слайд 2Азотистый обмен. Общие пути.
В организме человека содержится примерно 15 кг белков. Количество
Азотистый обмен. Общие пути.
В организме человека содержится примерно 15 кг белков. Количество

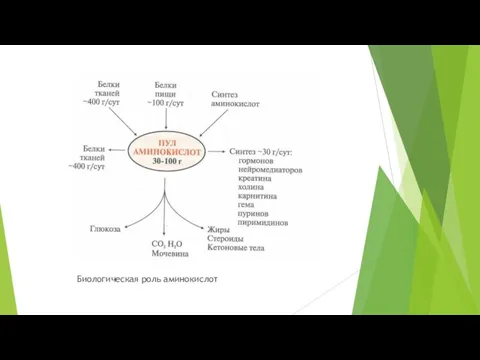
Слайд 3Биологическая роль аминокислот
Биологическая роль аминокислот
Слайд 4Переваривание белков в ЖКТ.
Переваривание белков включает гидролиз пищевых белков до свободных аминокислот.
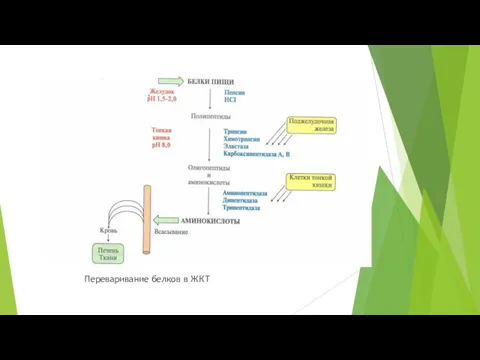
Переваривание белков в ЖКТ.
Переваривание белков включает гидролиз пищевых белков до свободных аминокислот.

Слайд 5Переваривание белков в ЖКТ
Переваривание белков в ЖКТ
Слайд 6Переваривание белков в желудке.
Переваривание белков в желудке происходит под действием пепсина. Профермент
Переваривание белков в желудке.
Переваривание белков в желудке происходит под действием пепсина. Профермент

Слайд 7
Переваривание белков в кишечнике
Происходит при pH ~ 8,0, изменение pH происходит под
Переваривание белков в кишечнике
Происходит при pH ~ 8,0, изменение pH происходит под

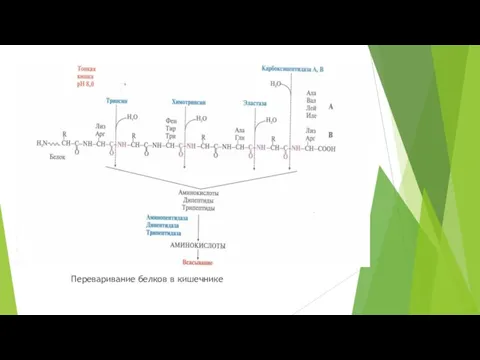
Слайд 8Переваривание белков в кишечнике
Переваривание белков в кишечнике
Слайд 9Трипсин синтезируется в поджелудочной железе в виде профермента трипсиногена. Превращение трипсиногена в
Трипсин синтезируется в поджелудочной железе в виде профермента трипсиногена. Превращение трипсиногена в

Слайд 10Ферменты, переваривающие белки в кишечнике обладают специфичностью к определенным аминокислотам:
Трипсин гидролизует пептидные
Ферменты, переваривающие белки в кишечнике обладают специфичностью к определенным аминокислотам:
Трипсин гидролизует пептидные

Слайд 11ЖКТ
ТКАНИ
КРОВЬ
ПЕЧЕНЬ
БЕЛКИ
АМ. КИСЛОТЫ
АМИН-ТЫ
АМИНЫ
ГЛЮКОЗА
Заменимые аминок-ты
БЕЛКИ КРОВИ
АМИН-ТЫ
БЕЛКИ КРОВИ
БЕЛКИ ПЕЧЕНИ
АМИН-ТЫ
БЕЛКИ
Биологически активные вещества
Продукты гниения: скатол, индол, фенол.
Обезвреживание
ЖКТ
ТКАНИ
КРОВЬ
ПЕЧЕНЬ
БЕЛКИ
АМ. КИСЛОТЫ
АМИН-ТЫ
АМИНЫ
ГЛЮКОЗА
Заменимые аминок-ты
БЕЛКИ КРОВИ
АМИН-ТЫ
БЕЛКИ КРОВИ
БЕЛКИ ПЕЧЕНИ
АМИН-ТЫ
БЕЛКИ
Биологически активные вещества
Продукты гниения: скатол, индол, фенол.
Обезвреживание
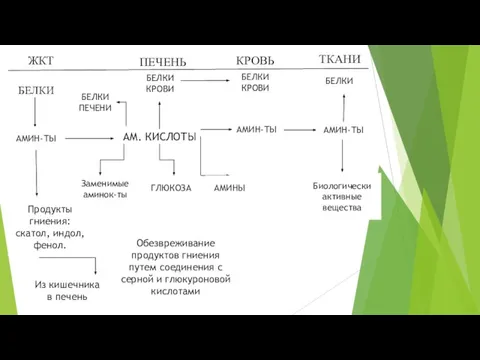
Слайд 12Часть аминокислот в кишечнике под действием микрофлоры подвергаются гниению с образованием токсичных
Часть аминокислот в кишечнике под действием микрофлоры подвергаются гниению с образованием токсичных
Слайд 13Общие пути обмена аминокислот
Пути распада аминокислот до конечных продуктов можно разделить на
Общие пути обмена аминокислот
Пути распада аминокислот до конечных продуктов можно разделить на

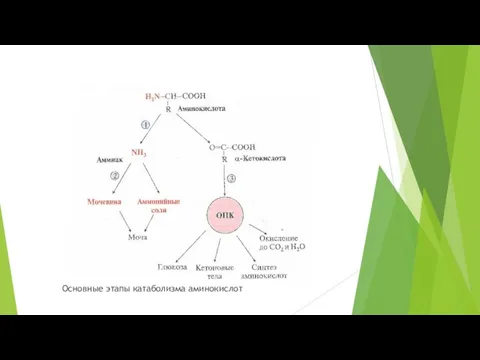
Слайд 14Основные этапы катаболизма аминокислот
Основные этапы катаболизма аминокислот
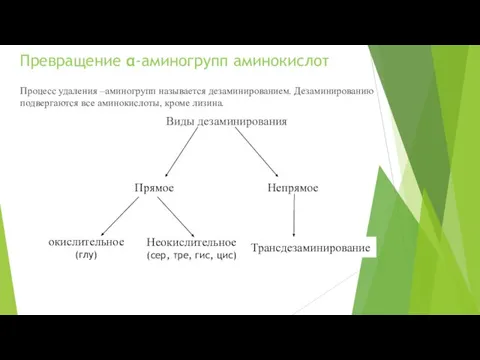
Слайд 15Превращение α-аминогрупп аминокислот
Процесс удаления –аминогрупп называется дезаминированием. Дезаминированию подвергаются все аминокислоты, кроме
Превращение α-аминогрупп аминокислот
Процесс удаления –аминогрупп называется дезаминированием. Дезаминированию подвергаются все аминокислоты, кроме
Слайд 16Виды дезаминирования
1. Окислительное дезаминирование (прямое, характерное для глу) происходит в митохондриях многих
Виды дезаминирования
1. Окислительное дезаминирование (прямое, характерное для глу) происходит в митохондриях многих

Слайд 172. Прямое неокислительное дезаминирование характерно для серина, треонина, цистеина и гистидина. Серин

Слайд 18Гистидин дезаминируется внутримолекулярным способом
Цистеин дезаминируется с выделением H2S c использованием воды.
Гистидин дезаминируется внутримолекулярным способом
Цистеин дезаминируется с выделением H2S c использованием воды.

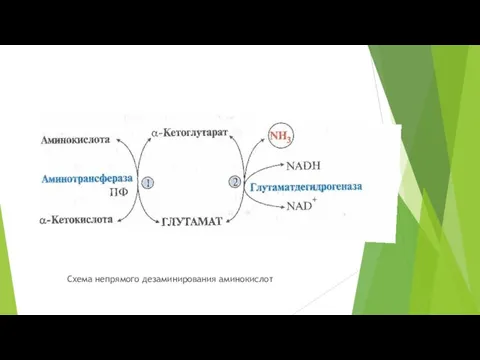
Слайд 19Большинство аминокислот в клетках непрямому дезаминированию, которое включает 2 стадии:
Трансаминирование с
Большинство аминокислот в клетках непрямому дезаминированию, которое включает 2 стадии:
Трансаминирование с

Слайд 20Схема непрямого дезаминирования аминокислот
Схема непрямого дезаминирования аминокислот
Слайд 21Аминотрансферазы, катализирующие реакцию трансаминирования обладают субстратной специфичностью. Наиболее важными являются АСТ и
Аминотрансферазы, катализирующие реакцию трансаминирования обладают субстратной специфичностью. Наиболее важными являются АСТ и

Слайд 22В результате трансаминирования образуется глутаминовая кислота. Затем глутаминовая кислота подвергается окислительному дезаминированию:
В результате трансаминирования образуется глутаминовая кислота. Затем глутаминовая кислота подвергается окислительному дезаминированию:

Слайд 23Клиническое значение определения активности аминотрансфераз
В сыворотке крови здоровых людей активность аминотрансфераз ниже,
Клиническое значение определения активности аминотрансфераз
В сыворотке крови здоровых людей активность аминотрансфераз ниже,

Слайд 24Декарбоксилирование аминокислот.
Процесс отщепления карбоксильной группы в виде СО2 называется декарбоксилированием и приводит
Декарбоксилирование аминокислот.
Процесс отщепления карбоксильной группы в виде СО2 называется декарбоксилированием и приводит

Слайд 25При декарбоксилировании Три, Гис, Фен, Глу образуются биогенные амины:
Серотонин обладает сосудосуживающим действием,
При декарбоксилировании Три, Гис, Фен, Глу образуются биогенные амины:
Серотонин обладает сосудосуживающим действием,

Слайд 26Недостаточность дофамина в черной субстанции мозга приводит к болезни Паркинсона. При этом
Недостаточность дофамина в черной субстанции мозга приводит к болезни Паркинсона. При этом

Слайд 27Гистамин обладает сосудорасширяющим действием. Много образуется в области воспаления, участвует в развитии
Гистамин обладает сосудорасширяющим действием. Много образуется в области воспаления, участвует в развитии

Слайд 28Орнитин, декарбоксилируясь, дает диамин путресцин (1,4-диаминобутан):
При декарбоксилировании лизина образуется кадаверин (1,5-диаминопентан):
Орнитин, декарбоксилируясь, дает диамин путресцин (1,4-диаминобутан):
При декарбоксилировании лизина образуется кадаверин (1,5-диаминопентан):

Слайд 29Превращение углеродного скелета аминокислот
Катаболизм всех аминокислот сводится к образованию 6 веществ, вступающих
Превращение углеродного скелета аминокислот
Катаболизм всех аминокислот сводится к образованию 6 веществ, вступающих

 Ткани растений. Физиология растения. Агротехнические приемы
Ткани растений. Физиология растения. Агротехнические приемы Презентация на тему Викторина по биологии "Как называется"
Презентация на тему Викторина по биологии "Как называется"  Все о собаках
Все о собаках Синдромы поражения краниальных нервов
Синдромы поражения краниальных нервов Полезная еда. Проростки гречки
Полезная еда. Проростки гречки Топография поверхностей (рельеф) полушарий мозга
Топография поверхностей (рельеф) полушарий мозга Медицинская гельминтология
Медицинская гельминтология Соцветия. Схемы соцветий
Соцветия. Схемы соцветий Геном про – та еукаріот
Геном про – та еукаріот Обмен веществ и энергии
Обмен веществ и энергии Биологическое действие радиации. Закон радиоактивного распада. Термоядерные реакции
Биологическое действие радиации. Закон радиоактивного распада. Термоядерные реакции Наглядный материал к предложенным занятиям - Грибы
Наглядный материал к предложенным занятиям - Грибы Влияние комнатных растений
Влияние комнатных растений Виды бактерий
Виды бактерий Занятие 6. Основные и выделительные ткани
Занятие 6. Основные и выделительные ткани Состав, строение и рост костей. 8 класс
Состав, строение и рост костей. 8 класс Platinum. 70% парного мяса для вашего питомца
Platinum. 70% парного мяса для вашего питомца Многообразие млекопитающих. 7 класс
Многообразие млекопитающих. 7 класс Саванна
Саванна Животный мир Песчанокопского района
Животный мир Песчанокопского района Анатомия и физиология кожи
Анатомия и физиология кожи Значение опорно-двигательного аппарата, его состав . Строение костей. 8 класс
Значение опорно-двигательного аппарата, его состав . Строение костей. 8 класс Контрольное тестирование по теме Анализаторы. Значение анализаторов в жизни человека
Контрольное тестирование по теме Анализаторы. Значение анализаторов в жизни человека Комнатные растения
Комнатные растения Мутации. Виды мутаций
Мутации. Виды мутаций Утомление при статической и динамической нагрузке
Утомление при статической и динамической нагрузке Бактерии, их строение
Бактерии, их строение Олигосахариды и их классификация
Олигосахариды и их классификация